БІОХІМІЯ - Підручник - Остапченко Л. І. - 2012
Розділ 4. МОЛЕКУЛЯРНА ОРГАНІЗАЦІЯ І БІОЛОГІЧНІ ФУНКЦІЇ МЕМБРАН
4.2. Молекулярна організація мембран
4.2.1. Ліпіди
Ліпіди є амфіфільними молекулами, оскільки складаються як із гідрофільних, так і з гідрофобних частин. Тому у водних розчинах ліпідні молекули можуть взаємодіяти як з молекулами води, так і між собою. На межі поділу фаз "вода - повітря» такі молекули утворюють моношари. За відсутності обмежень ліпіди намагаються зайняти максимальну водну поверхню й така структура аналогічна "двовимірному газу" (рис. 4.2): молекули ліпідів не взаємодіють. У разі зменшення площі водної поверхні молекули починають взаємодіяти між собою, утворюють суцільну мономолекулярну плівку, яка відповідає стану "двовимірній рідині". При подальшому стискуванні моношару молекули ліпідів упорядковують свою орієнтацію в моношарі так, що утворюється "частокіл" - конденсований моношар (рис. 4.2). Коли тиск перевищить граничну величину (молекули не можуть більше наближатись одна до одної) - тиск колапсу - моношар руйнується.

Рис. 4.2. Схема залежності поверхневого тиску від площі, яка припадає на одну молекулу ліпідів у моношарі:
моношари: 1 - газоподібний; 2 - "рідкий"; 3 - конденсований ("твердий"); 4 — колапс
Коли ліпіди знаходиться у воді (не на поверхні) і їх мало, то вони розсіяні по всьому об'єму. Коли їхня концентрація більша, ліпідні молекули частіше зіштовхуються й займають енергетично більш вигідне положення - утворюються різні агрегати, в яких гідрофобні хвости заховані всередину, а гідрофільні голівки контактують з водою. Однією з таких структур є міцели, а також циліндри, еліпсоїди тощо. Ця властивість амфіфільних молекул називається поліморфізмом.
Окрім таких моношарових структур, у воді утворюється бімолекулярна ліпідна мембрана (БЛМ) у якій голівки ліпідних молекул контактують з водою, а гідрофобні хвости - один з одним. Цей процес також енергетично вигідний, у чому й полягає самозбирання ліпідних бішарових мембран. Першу таку мембрану одержала наукова група П. Мюллера (1962). Для отримання БЛМ краплю розчину наносять під водою на отвір (1-2 мм) у тонкій тефлоновій перегородці, по обидва боки якої міститься розчин електроліту (найчастіше 100 ммоль/л NaCl). При дії поверхневих сил крапелька поступово розтікається і стоншується. Коли її товщина наближається до довжини хвилі видимого світла, мембрана має колір райдуги (d ліпідної голівки дорівнює 100-600 нм). Через кілька хвилин плівка стає чорною. Це означає, що плівка має товщину, меншу за довжину хвилі видимого світла й така плівка не відбиває світла (тому й чорна). У процесі стоншення плівки органічний розчинник витісняється до країв, а мембрана набуває товщини двох ліпідних молекул (рис. 4.3). Завдяки можливості проведення різних електричних вимірів (провідність, ємність, потенціал пробою, мембранний потенціал тощо) БЛМ відіграє важливу роль у вивченні іонного транспорту біологічних мембран, реконструкції окремих білкових компонентів природних мембран на БЛМ, створення біосенсорних систем тощо.
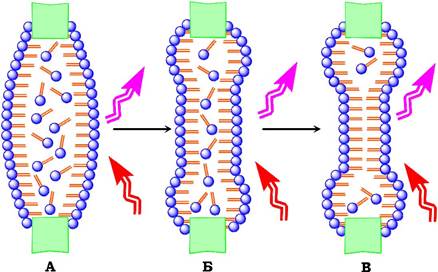
Рис. 4.3. Схема процесу формування БЛМ:
А - товста плівка після нанесення краплі розчину ліпіду на отвір тефлонової перегородки. Б - стоншення плівки.
В - БЛМ з товщиною 4-6 нм). Стрілками показано падаюче й відбите світло
Коли бішарові структури формуються у воді, бішар у разі його достатньої довжини замикається на себе, утворюючи везикули, або ліпосоми, хоча в історичному плані ліпосомами вперше були названі частки, які утворюються при механічному диспергуванні фосфоліпідів у воді. Такі структури є багатошаровими, тому тепер їх називають мультиламелярними везикулами: вони складаються з багатьох ліпідних бішарів, розділених водними проміжками (рис. 4.4), і мають розміри до 50 мкм. Найменші ліпосоми (~ 20 нм) утворені одним ліпідним бішаром (ММВ).
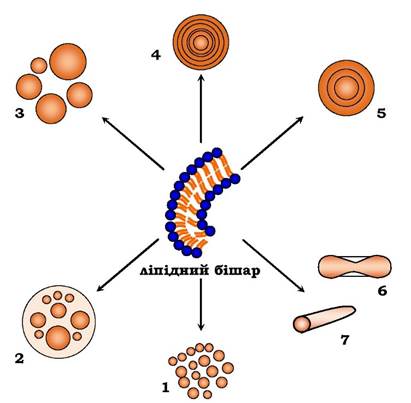
Рис. 4.4. Види ліпосом: 1 - малі моноламелярні везикули (ММВ);
2 - оліговезикулярні ліпосоми (ОВЛ); 3 - великі моноламелярні везикули (ВМВ); 4 - мультиламелярні везикули (МЛВ);
5 - оліголамелярні везикули (ОЛВ); 6 - дискоми (дископодібні ліпосоми); 7 - тубулярні (трубчасті) везикули
Ліпосоми як моделі стоять ближче до біологічних мембран, ніж БЛМ. Як і біологічні мембрани, вони є замкнутими системами, що робить їх придатними для вивчення трансмембранного транспорту речовин, вони стабільніші, ніж БЛМ, у них легко змінювати ліпідний склад, включати в їхній склад мембранні білки (тоді вони називаються протеоліпосомами) і моделювати не тільки транспортні, а й ферментативні та рецепторні функції клітинних мембран. Їх використовують для доставки лікувальних речовин, пептидів, білків і нуклеїнових кислот (рис. 4.5). А це є перспективним у хіміотерапії раку, лікуванні діабету, артриту, лейшманіозу, корекції ферментативної недостатності й дефектів клітинних мембран, у генній інженерії та біотехнології.
Рис. 4.5. Основні способи включення різних речовин у ліпосоми:
водорозчинні речовини (білки, іони) включаються в ліпосоми у процесі їхнього утворення; гідрофобні молекули можна включати в ліпідний бішар (1) або адсорбувати (чи хімічно зв'язувати) речовини на поверхню ліпосоми (2)
Основна маса ліпідів у мембрані представлена фосфоліпідами - фосфатидилетаноламінами (ФЕ), фосфатидилхолінами (ФХ), фос- фатидилсеринами (ФС), а також кардюліпіном, фосфатидилінозитолами, гліко- та сфінголіпідами і стеринами. ФЕ і ФХ становлять у мембранах 65-85 % усіх ліпідів. У плазматичних мембранах високий вміст стеринів - холестеролу у тварин і ергостеролу в рослин. Стерини займають "пустоти" між молекулами фосфоліпідів у ділянці подвійних зв'язків жирнокислотних ланцюгів і цим зміцнюють мембранну структуру. Орієнтуючись у бішарі таким чином, що гідроксильні групи холестеролів контактують з полярними голівками фосфоліпідів, а стероїдні кільця іммобілізують ділянки вуглеводневих ланцюгів, холестерол знижує плинність мембрани в цілому. Окрім цього, холестерол перешкоджає "злипанню" та "кристалізації" вуглеводневих ланцюгів, чим гальмує можливі фазові переходи (рідкий стан → кристалічний стан). Наслідком цих процесів є зміни проникності мембрани, пружності й механічної міцності бішару. Це сприяє змінам форм мембрани за дії прикладеної сили, і пояснюється тим, що холестерол може швидко перерозподілитися між моношарами завдяки низькому енергетичному бар'єру для механізму транспорту на кшталт фліп-флопу.
Залежно від умов життя організмів (сезонність, солоність води, адаптація до умов середовища) клітинні мембрани за складом ліпідів мають значні відмінності. Вони різняться складом жирно- кислотних хвостів мембранних ліпідів. Так, зниження температури довкілля викликає збільшення кількості подвійних зв'язків у жирнокислотних ланцюгах мембранних ліпідів холоднокровних тварин. Це забезпечує більш рихлу упаковку бішару й протидіє зростанню в'язкості (зменшенню плинності) мембрани при низькій температурі. Такі ж зміни складу і властивостей мембран може викликати зростання концентрації солей, яке відбувається при міграції риб із річок у моря. Такі варіації в жирнокислотному складі мембранних ліпідів під впливом зовнішніх умов є адаптацією, яка забезпечує пристосування організму до змін довкілля. Разом із тим, склад основних ліпідів мембран досить стабільний, що є результатом еволюції.
Крім основних мембранних ліпідів (табл. 4.1), у мембранах присутні й інші ліпіди в невеликих кількостях. Тому вони називаються мінорними. До них відносять вільні жирні кислоти, лізофосфоліпіди, моноацил- і діацилгліцероли, поліізопреноїдні ліпіди (убіхінон і механохінон - компоненти ланцюга перенесення електронів, ундакапренол і доліхол - ліпідні переносники проміжних продуктів при біосинтезі клітинної стінки у прокаріотів і при біосинтезі глікопротеїнів у ендоплазматичному ретикулумі еукаріотів відповідно) тощо.
Таблиця 4.1
Склад ліпідів окремих типів мембран
(узагальнені дані, % від загального вмісту ліпідів)
Ліпіди |
Плазматична мембрана |
Мембрани мітохондрії (обидві) |
Ендоплазматичний ретикулум |
Мієлін |
||
Еритроцитів |
Гепатоцитів |
E. coli |
||||
Фосфатидилетаноламін |
18 |
7 |
70 |
35 |
17 |
15 |
Фосфатидилхолін |
17 |
24 |
0 |
39 |
40 |
10 |
Фосфатидилсерин |
8 |
4 |
~ 0 |
2 |
5 |
9 |
Сфінгомієлін |
19 |
19 |
0 |
0 |
5 |
8 |
Холестерол |
23 |
17 |
0 |
3 |
6 |
|
Інші ліпіди |
13 |
22 |
30 |
21 |
27 |
8 |
Завдяки руху мембранних ліпідів перпендикулярно площині мембрани (фліп-флоп) і обміну ліпідів між мембранами при контактах мембран, везикулярному транспорту (ендо- та екзоцитоз тощо) і за рахунок ліпід-переносних білків у кожній мембрані існує асиметрія ліпідів. Так, для малих моноламелярних везикул із суміші фосфоліпідів зовнішній шар збагачений сфінгомієліном, а внутрішній - фосфатидилсерином. Це тому, що ліпіди з більш об'ємними полярними голівками намагаються локалізуватись там, де більша площа на молекулу (а це зовнішній моношар). В еритроцитах фосфатидилхолін і сфінгомієлін містяться переважно в зовнішньому моношарі, а ФЕ і ФС - головним чином у внутрішньому. Причиною цього є пряме зв'язування білків цитоскелета з амінофосфоліпідами. До того ж трансмембранна міграція амінофосфо- ліпідів (фліп-флоп) каталізується ферментами типу транслоказ, що підтримує їхній стаціонарний трансмембранний розподіл.
У плазматичних мембранах тваринних клітин найбільш асиметрично розподілені гліколіпіди. Вони локалізовані лише в зовнішньому моношарі, а їхні сахаридні групи "звисають" у глікокалікс. Це свідчить про участь таких молекул у міжклітинних взаємодіях і взаємодії клітини з її оточенням. Найскладніші гліколіпіди - гангліозиди, локалізуючись у зовнішньому моношарі ліпідного мактриксу мембран, виступають у ролі рецепторів для певних медіаторів міжклітинної взаємодії, а також як структури, що іммобілізують токсини, у т. ч. і холерний токсин.
Оскільки ліпідний матрикс змінюється залежно від сезонних явищ і умов довкілля, з віком організму, з розвитком різних патологій, то це також відбивається і на характері функціонування мембранних білків.