БІОХІМІЯ - Підручник - Остапченко Л. І. - 2012
Розділ 4. МОЛЕКУЛЯРНА ОРГАНІЗАЦІЯ І БІОЛОГІЧНІ ФУНКЦІЇ МЕМБРАН
4.3. Функції мембран
4.3.1. Мембранний транспорт
Живі системи на всіх рівнях організації, у тому числі клітина і клітинні органоїди, є відкритими термодинамічними системами. Тому мембранний транспорт речовин є необхідною умовою життя. Порушення транспорту речовин через біомембрани приводить до патології. А саме лікування здебільшого також зв'язане із взаємодією і перенесенням лікувальних речовин через мембрани як безпосередньо, так і опосередковано ліпосомами. Транспорт речовин через клітинні мембрани можна поділити на два типи: активний і пасивний (рис. 4.8).
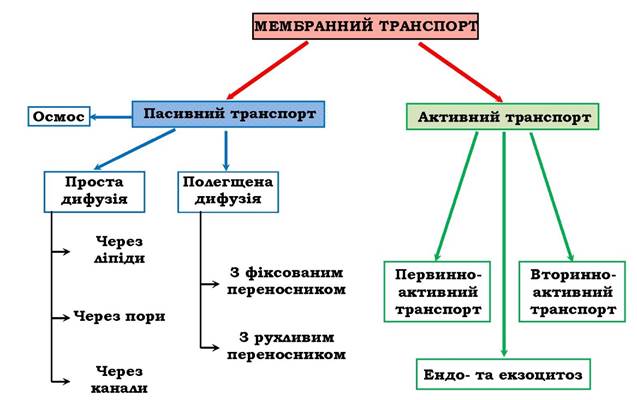
Рис. 4.8. Схема основних видів трансмембранного переміщення речовин
Зрозуміло, що чим ліпше речовини розчиняються в ліпідах, тим легше вони проходять через мембрану - це називається дифузією. Полярні речовини через мембрану проходять гірше, оскільки вони нерозчинні в ліпідах. Разом із тим, вода проникає добре (її Р = 10-2 см/с). Це пояснюється не тільки тим, що вода переноситься через іонні канали та ліпідні пори (див. далі), а й тим, що вона переноситься тиками - вільними порожнинами між хвостами фосфоліпідів (kink - петля), які утворюються при їхньому тепловому русі. Вони переносять упоперек мембрани дрібні гідрофільні молекули, у першу чергу молекули води, що в них попали.
Полегшена дифузія відбувається за участю молекул- переносників. Прикладом є антибіотик валіноміции - переносник іонів K+. Це циклічний депсипептид з м. м. 1111: L-лактат-L-валін-D-оксіізовалеріанова кислота-Д-валін. Особливістю хімічної будови валіноміцину є те, що його полярні групи локалізовані всередині структури, а зовні локалізуються неполярні гідрофобні залишки молекул валіну. Така структура дозволяє утворювати з іонами калію комплекс, розчинний (як і вільний валіноміцин) у ліпідній фазі мембрани. Тому такі комплекси "захоплюються" мембраною, у ній вони дифундують за градієнтом концентрації
й переносять калій на інший бік мембрани. Вільний від калію валіноміцин (також за градієнтом його концентрації) повертається назад і комплексує з новим іоном K+ (рис. 4.9). Таким чином відбувається човникове перенесення іонів калію через мембрану.
Інший тип полегшеної дифузії - канальне перенесення іонів. Прикладом цього є наведений на рис. 4.9, В канал з граміцидину. Молекули цього антибіотика легко вбудовуються в мембрану (завдяки його гідрофобній поверхні) і, димеризуючись N-кінцями, утворюють канал, який пропускає катіони на кшталт естафети, не змінюючи свого положення в мембрані. Такі два типи речовин називаються відповідно іонофорами та каналоформерами.

Рис. 4.9. Схема індукованого іонного транспорту:
А - іонофором валіноміцином і В - каналоформером граміцидином.
Б - структура комплексу K+-валіноміцин: іон K+ фіксується в центрі за рахунок іон-дипольної взаємодії за участю карбонільних груп пептиду
Отже, відмінності полегшеної дифузії від простої такі:
✵ перенесення речовин за участю переносників проходить швидше.
✵ полегшеній дифузії притаманна властивість насичення (коли всі молекули переносників зайняті речовиною, то збільшення її концентрації не змінює інтенсивність потоку речовини(рис. 4.10).
✵ є речовини, які блокують полегшену дифузію - інгібітори, утворюючи міцний комплекс з молекулами переносника.
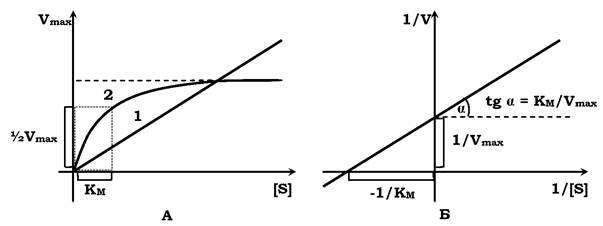
Рис. 4.10. Кінетика простої (1) і полегшеної (2) дифузій:
А: 1 - швидкість пропорційна концентрації речовини, яка переміщується через мембрану; 2 - швидкість досягає максимального значення (Vmax) при насиченні білків-переносників. Константа зв'язування речовини з переносником (Км) - це концентрація речовини, за якої ν = 1/2 Vmax. [S] - концентрація речовини, яка транспортується через мембрану.
Б - крива 2 в координатах Лайнуївера - Берка, які дають можливість визначити основні кінетичні параметри процесу транспорту: константу Міхаеліса Км і максимальну швидкість процесу Vmax
Якщо молекула речовини не несе заряду, то напрямок її трансмембранного руху визначається тільки різницею концентрації (градієнта концентрації) цієї речовини по обидва боки мембрани - хімічним потенціалом. Якщо молекула несе електричний заряд, то на її трансмембранний рух впливає не тільки хімічний потенціал, а й різниця електричних потенціалів на мембрані (мембранний потенціал (див. далі) - електричний потенціал. Разом хімічний і електричний потенціали становлять електрохімічний потенціал - μ.
Одні білки-переносники переносять одну речовину через мембрану - таке перенесення називається уніпортом Інші білки є котранспортними системами, коли перенесення однієї речовини залежить від перенесення іншої. Якщо ці дві речовини переносяться в одному напрямку - це симпорт, якщо у протилежних - це антипорт. Прикладом уніпорту є перенесення глюкози в м'язову клітину (де її концентрація завжди нижча, ніж у позаклітинному просторі). Прикладом симпорту є поглинання епітеліальними клітинами кишечнику глюкози разом з іонами Na+, позаклітинна концентрація яких у позаклітинному просторі висока. Прикладом антипорту є перенесення іонів Cl- і HCO3- білком смуги 3 еритроцитів та перенесення Na+ і Ca2+ натрій-кальцієвим антипортом у клітинах м'язів і нервових клітинах.
Молекулярні механізми роботи білків-переносників полягають в оборотних конформаційних змінах їхніх молекул (рис. 4.11). Ці зміни конформацій забезпечують зв'язування речовин з одного боку мембрани й вивільнення його з іншого боку. Як видно з рис. 4.11, білок-переносник існує в трьох конформаційних станах: А - відкритий із зовнішнього боку ("понг"-стан); Б - проміжний, навантажений глюкозою; В - відкритий із цитоплазматичного боку ("пінг"-стан). Така система "пінг-понг" забезпечує перенесення глюкози з позаклітинного середовища, оскільки її там більше. Пунктиром позначено місця зв'язування іона Na+. За рахунок градієнта концентрації натрію швидкість перенесення глюкози збільшується - симпорт.

Рис. 4.11. Схема забезпечення конформаційними змінами білка-переносника (уніпорт) глюкози через мембрану.
Симпорт - транспорт глюкози разом з іонами Na+
Канали і пори (табл. 4.2) також змінюють конформації своїх білкових складових, але такі зміни регулюють лише їхнє відкривання і закривання і не стосуються самого перенесення. За способом регуляції канали поділяються на потенціалозалежні та хімічно чутливі. Канали першого типу відповідають на зміни трансмембранного потенціалу, другого - відкриваються на дію специфічних хімічних агентів (нейромедіатори). Терміни "пора" і "канал" часто використовують як синоніми. Але це не так. Пора - це неселективна мембранна структура, яка "розпізнає" речовини в основному за розмірами молекул. У процесі транспорту вони не змінюють своєї конформації, а тому пропускають лише невеликі молекули. Під каналами розуміють іонні канали, які селективно транспортують основні "фізіологічні" катіони. Тому й називаються K+ -, Na+ -, Ca2+ -, H+ -каналами, тобто пора - це постійно відкрита структура, канал - структура, яка змінює свою структуру, "дихає", може перебувати щонайменше в трьох станах: закритому, проміжному й відкритому.
Таблиця 4.2
Класифікація мембранних транспортних систем
Системи та швидкість транспорту за секунду |
Характеристика |
Приклади |
І. Пори, ~ 107 |
Неселективні структури, "розпізнають" речовини в основному за розмірами |
Пори у бактерій і в міто хондріях, пори утворені антибіотиками (граміцидин, аламетицин та ін.) |
ІІ. Канали, ~ 106-107 |
1. Потенціалозалежні |
K+ -, Ca2+ - і Na+ -канали клітин збудливих тканин |
2. Хімічно чутливі |
Ацетилхоліновий рецептор |
|
3. Інші канали, які електричним потенціалом і хімічними речовинами не регулюються |
Рецептори до механічного тиску |
|
ІІІ. Пасивні переносники, ~ 100 |
1. Уніпорт |
Переносник глюкози в еритроцитах |
2. Симпорт |
Переносник глюкози й Na+ в епітеліоцитах кишечнику |
|
3. Антипорт |
Переносник Na+ усередину, а Ca2+- назовні міоцитів |
|
Активні переносники, 50-103 |
а) Спряжені з поглинанням світла - бактеріородопсин. |
Мембрани пурпурних бактерій |
б) Спряжені з окисно- відновними реакціями - ци- тохромоксидаза |
Внутрішня мембрана мітохондрій |
|
в) АТФази: Na+, K+-АТФаза, Mg2+ , Ca2+ -АТФаза |
Плазматична мембрана клітин |
Осмос як тип пасивного транспорту є, по суті, дифузією води з місць її більшої концентрації в місця з меншою концентрацією (або - рух води в бік вищих концентрацій розчинів). Осмос надзвичайно важливе явище в багатьох біологічних процесах, у тому числі гемолізі еритроцитів у гіпотонічних розчинах, тургорі у рослин тощо.
За рахунок активного транспорту в організмах створюються і різниці (градієнти) концентрацій речовин, електричних потенціалів, тиску. Тобто, з точки зору термодинаміки, активне перенесення речовин та іонів утримує організм у нерівноважному стані (рівновага - це смерть організму). У біологічних
мембранах є кілька типів іонних насосів, які працюють за рахунок енергії гідролізу АТФ. Є три типи електрогенних насосів (рис. 4.12), основою яких є три транспортні АТФази. При роботі K+ ,Na+ -АТФази за рахунок енергії макроергічного зв'язку однієї молекули АТФ у клітину переноситься два іони К+ , а із клітини - три іони Na+. Це створює градієнти К+ і Na+ на плазматичній мембрані, які направлені у клітину (для калію) і з клітини (для натрію). Оскільки на одну молекулу АТФ викачується на один іон більше, ніж закачується (1 АТФ : 2K+ : 3Na+), на мембрані утворюються і різниця (градієнт) потенціалів, яка називається мембранним потенціалом спокою (МПС) зі знаком "мінус" з цитоплазматичного боку (див. 4.3.2). Са-АТФаза забезпечує активний транспорт двох іонів Ca2+ у цистерни ендоплазматичного рети- кулума, а протонна (Н+-АТФаза) - двох протонів на одну молекулу АТФ (рис. 4.12). Молекулярний механізм такого транспорту у випадку натрієвого насосу включає сім етапів перенесення іонів, спряжених з гідролізом АТФ. Зі схеми, зображеної на рис. 4.12, Б, видно, що ключовими етапами роботи ферменту є: 1) утворення комплексу ферменту з АТФ на цитоплазматичному боці, що активується іонами Mg2+; 2) зв'язування комплексом трьох іонів Na+ ; 3) фосфорилювання ферменту з утворенням аденозиндифосфату; 4) "транслокація" (фліп-флоп) ферменту з іонами натрію всередині мембрани; 5) реакція іонного обміну натрію на калій, що відбувається вже на зовнішньому боці ПМ; 6) зворотна транслокація ферментного комплексу з перенесенням двох іонів калію всередину клітини; 7) набуття ферментом вихідного стану з вивільненням іонів калію і неорганічного фосфату.
Розглянутий активний транспорт називається первинним. Існують системи і вторинного активного транспорту речовин, насоси для яких відсутні (вуглеводи, амінокислоти). Транспорт речовин у цьому випадку опосередкований мембранним потенціалом і/або градієнтом концентрації іонів за наявності в мембрані специфічних білкових переносників. Є кілька типів вторинного активного транспорту. Один із них - це транспорт одновалентних іонів за участю переносника, який у "навантаженому" і вільному станах однаково добре перетинає мембрани. Джерелом енергії тут є МПС. Однонаправлене перенесення у комплексі зі специфічним переносником називається (як і для полегшеної дифузії) уніпортом. Результатом перенесення буде накопичення іонів за рахунок зниження мембранного потенціалу. Прикладом уніпорту є накопичення іонів K+ у присутності валіноміцину в мітохондріях (див. рис. 4.9).

Рис. 4.12. Схема активного транспорту іонів транспортними АТФазами (А) і етапи такого транспорту для Na+, K+-АТФази (Б): Е - фермент АТФаза на внутрішній (Е1) і зовнішній (Е2) поверхнях ПМ;
Р - неорганічний фосфат у реакції АТФ → АДФ+Р; * - активний комплекс.
В - схема локалізації Na+, K+-АТФази в мембрані у вигляді тетрамера: а- і β- субодиниці ферменту
Протилежний потік іонів за участю молекул переносника називається антипортом. Перенесення відбувається в два етапи: спочатку один іон перетинає мембрану зліва направо, а потім другий іон - у зворотному напрямку (рис. 4.13). Важливо, що мембранний потенціал при цьому не змінюється, а рухомою силою цього перенесення є градієнт концентрації одного з іонів, яка створена первинним активним транспортом. Класичним прикладом антипорту є перенесення через плазматичну мембрану іонів калію і водню за участю антибіотика нігерицину.
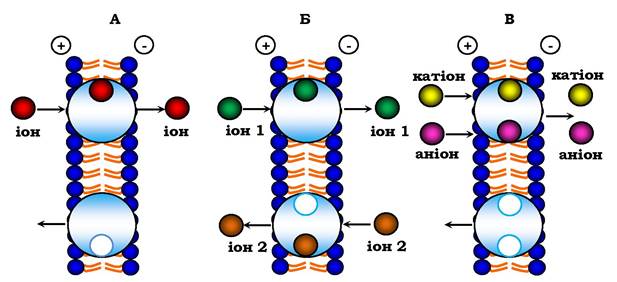
Рис. 4.13. Схеми трьох основних типів вторинного активного транспорту іонів:
А - уніпорт; Б - антипорт; В — симпорт
При симпорті у мембрані знаходяться дві електронейтральні частки: переносник з катіоном і аніоном та порожній переносник (рис. 4.13, В). Оскільки мембранний потенціал за цією схемою не змінюється, то причиною перенесення є різниця концентрації одного з іонів. Вважається, що за схемою симпорту відбувається накопичення клітинами амінокислот. Калієво-натрієвий насос створює початковий градієнт концентрації іонів натрію (рис. 4.12, А), які потім за схемою симпорту сприяють накопиченню в клітині амінокислот. Зі схеми симпорту видно, що процес має супроводжуватись значними змінами осмотичної рівноваги, оскільки в одному циклі через мембрану переноситься дві частки в одному напрямку.
Треба зауважити з приводу класифікації типів транспорту речовин через мембрану. Така класифікація як за структурними особливостями, так і за енергетичними затратами є умовною. Так, терміни переносники, транслокази, пермеази, які в різних підручниках часто використовуються стосовно транспортних білків, є синонімами. Зазвичай термін пермеаза застосовують при описанні бактеріальних транспортних білків. Термін "переносник" частіше використовують щодо іонофорів і переносників у плазматичних мембранах еукаріотів, а "транслоказа" - при описанні загальних питань трансмембранного переміщення речовини. Що ж до поділу процесів мембранного транспорту речовин, то тут ще більше умовностей. Узагалі, транспорту речовин через мембрану без використання енергії немає. Перенесення іонів за допомогою іонофорів і каналоформерів (про що йшлося вище) можливе лише тоді, коли є градієнт концентрації цих іонів. А градієнти в нормальних умовах утворюються тільки з використанням енергії. Тому в нашому прикладі (рис. 4.9) перенесення іонів К+ валіноміцином (уніпорт) можливе лише після того, коли Na+, K+ -АТФаза за рахунок енергії гідролізу АТФ створила градієнт концентрації калію. Перенесення амінокислот за рахунок зниження градієнта іонів Na+ (симпорт) теж не може відбутися, поки клітина не створить цей градієнт за рахунок енергії гідролізу АТФ мембранною АТФазою. Антипорт - це теж процес, який збільшує градієнт одного іона (речовини) за рахунок зниження градієнта іншого типу, що і є рушійною силою антипорту (рис. 4.13). Навіть дифузійні процеси в мембрані за великим рахунком є процесом енергозатратним: напрямок дифузійного перенесення речовини визначається градієнтом її концентрації, на створення якого потрібно витратити енергію.
У клітинах тварин котранспортуючим іоном зазвичай є іон Na+. Його електрохімічний градієнт забезпечує енергією активний транспорт іншого іона чи нейтральної молекули. Кожна транспортна система мембрани (білкові компоненти) специфічна для перенесення всередину клітини невеликої групи споріднених цукрів або амінокислот (що чітко видно при розгляді процесів усмоктування їх епітеліоцитами кишечнику). У таких системах ці молекули та іони Na+ зв'язуються з різними ділянками на білку-переноснику (рис. 4.10): іони натрію намагаються ввійти в клітину за своїм градієнтом концентрації й "тягнуть" у тому ж напрямку глюкозу (амінокислоту). Чим вищий градієнт Na+ , тим більша швидкість транспорту.
У рослин і тварин як котранспортуючий іон використовується Н+. Прикладом цього є пермеаза - переносник лактози. Це трансмембранний білок з одного поліпептидного ланцюга, який перетинає мембрану дев'ять разів. З кожною молекулою лактози в клітину приноситься один протон, градієнт якого підтримується Н+ -АТФазою.
У всіх клітинах підтримується постійне значення рН (7,1-7,2). Це, в основному, забезпечується Na+, H+ -антипортом: надлишки протонів у клітині, які постійно утворюються в окисно- відновних реакціях, видаляються за рахунок надходження в клітину іонів Na+. Цей переносник-обмінник активується низькими значеннями рН, що зумовлено зв'язуванням H+ на цитоплазматичному боці мембрани. Інший обмінник, навпаки, активується високими значеннями рН (~ 7,7). Це Cl- , HCO3 -антипорт: виведення HCO3 в обмін на СI- знижує рН цитозолю.
Розглядаючи мембранний транспорт, не можна обійти питання, як клітина "забороняє" вихід із неї речовини тим самим шляхом, яким вона увійшла. Такі механізми є. Скажімо, глюкоза, яка транспортувалася у клітину, може модифікуватися або транспортуватись із неї в інше "потрібне" місце. Останній спосіб - це трансцитоз, транспорт глюкози з епітеліоцитів кишечнику в кров. При трансцитозі в клітину глюкоза потрапляє завдяки Kа+-залежному симпорту, локалізованому в апікальній частині плазматичної мембрани, і виходить із клітини за градієнтом концентрації шляхом уніпорту, локалізованого у базолатеральній мембрані. Щільні міжклітинні контакти перешкоджають дифузії переносників систем симпорту й уніпорту по всій плазматичній мембрані. Тому, білки-переносники утримуються у своїх мембранних доменах (асиметрія мембран) і забезпечують трансцитоз. Але частина глюкози в таких епітеліоцитах не транспортується в кров трансцитозом, а використовується на власні енергетичні потреби. Для цього глюкоза фосфорилюється глюкозо-6-фосфатазою (донором фосфату є АТФ), з утворенням глюкозо-6- фосфату. Оскільки це вже заряджена молекула, то вийти вона не може й бере участь у процесах гліколізу. У бактерій аналогічна система як донор високоенергетичного фосфату використовує фосфоенолпіруват. Оскільки фосфатні групи переносяться на молекулу глюкози після надходження їх у клітину, цей тип транспорту інколи називають векторним, або направленим перенесенням груп.
Отже, у процесі життєдіяльності клітини її плазматичну мембрану перетинають різні речовини, потоки яких ефективно регулюються. Це забезпечується мембранними транспортними системами, які включають іонні насоси, молекули-переносники і високоселективні канали.