БИОЛОГИЯ Том 1 - руководство по общей биологии - 2004
7. АВТОТРОФНОЕ ПИТАНИЕ
7.4. Строение листа
У цветковых растений основным фотосинтезирующим органом является лист. В листе, как и во всех живых органах, структура и функция тесно взаимосвязаны. Из уравнения фотосинтеза

можно заключить, что, во-первых, листу требуются источники диоксида углерода и воды, во- вторых, лист должен содержать хлорофилл и быть способным поглощать солнечный свет, в- третьих, кислород улетучивается как побочный продукт, и, наконец, углевод как полезный продукт должен транспортироваться в другие части растения или запасаться. Структура листа прекрасно адаптирована для удовлетворения этих требований. На рис. 7.2 показано внешнее строение листа. Рис. 7.3 дает представление о микроскопическом строении листа двудольного растения. На рис. 7.4 показаны детали строения клетки палисадной паренхимы мезофилла листа. На рис. 7.5 схематично изображен поперечный срез листа двудольного растения. (Рекомендации по зарисовке объектов в световом микроскопе приведены в разд. 5.13.) Строение эпидермиса различных типов листьев представлено на рис. 6.3, а детали строения и функции устьиц обсуждаются в гл. 13.
Строение и функции различных тканей листа двудольного растения представлены в табл. 7.1.

Рис. 7.2. Внешнее строение листа двудольного растения.

Рис. 7.3. Поперечный срез листа бирючины (Ligustrum) в области средней жилки. Бирючина — типичное двудольное растение.
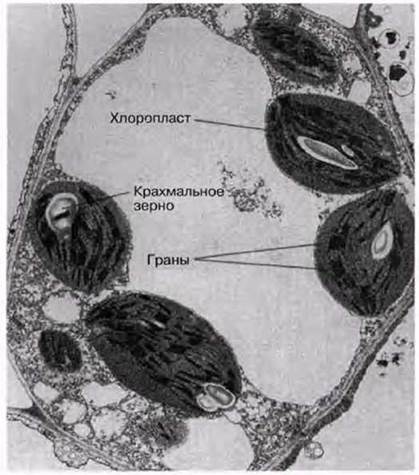
Рис. 7.4. Электронная микрофотография клетки палисадной паренхимы мезофилла (х3000).

Рис. 7.5. Схема поперечного среза листа типичного двудольного растения.
Таблица 7.1. Строение и функции листа двудольного растения
Ткань |
Строение |
Функция |
Верхний и нижний эпидермис |
Толщина в одну клетку. Клетки плоские, в них нет хлоропластов. Наружные стенки покрыты восковой кутикулой, образованной кутином. Эпидермис содержит устьица (поры), более многочисленные в нижнем эпидермисе. Каждое устьице образовано парой замыкающих клеток |
Защитная Кутан водоустойчив, защищает ткани листа от потери влаги и от инфекций. Через устьица происходит газообмен с окружающей средой. Размеры устьиц регулируются замыкающими клетками — специальными эпидермальными клетками, содержащими хлоропласты |
Палисадная паренхима мезофилла |
Столбчатые клетки с большим количеством хлоропластов в тонком слое цитоплазмы |
Основная фотосинтезирующая ткань. Хлоропласты способны перемещаться к свету |
Губчатая паренхима мезофилла |
Клетки неправильной формы, расположены свободно, с большими воздушными пространствами между ними (межклетниками) |
Фотосинтезирующая ткань, но содержащая меньше хлоропластов, чем столбчатая Газообмен осуществляется через большие межклетники и устьица Содержит запасы крахмала |
Проводящая ткань |
Сильно разветвленная сеть по всему листу |
Проводит воду и минеральные соли по ксилеме в лист Отводит продукты фотосинтеза (в основном сахарозу) по флоэме. Служит «скелетом», поддерживающим листовую пластинку (за счет колленхимы средней жилки листа, тургесцентности клеток мезофилла и в некоторых случаях склеренхимы) |
7.2. Перечислите особенности строения листа, которые обеспечивают его успешное функционирование.
И последнее, что следует отметить — это расположение листьев, обеспечивающее их минимальное перекрывание. Такое мозаичное расположение особенно заметно у некоторых растений, например у плюща.
7.4.1. Хлоропласты
У эукариот фотосинтез протекает в органеллах, называемых хлоропластами. Их число может варьировать от одного (как у одноклеточной водоросли Chlorella) до примерно ста (как в клетках палисадной паренхимы). Диаметр хлоропластов составляет 3—10 мкм (в среднем около 5 мкм), поэтому они хорошо видны в световой микроскоп (рис. 5.2 и 7.3). Хлоропласты окружены двойной мембраной, которая образует оболочку хлоропласта. Они всегда содержат хлорофилл и другие фотосинтетические пигменты, расположенные на системе мембран. Мембраны погружены в основное вещество, или строму. Детали строения хлоропластов можно выявить при помощи электронного микроскопа. На электронной микрофотографии низкого разрешения (рис. 5.11, 5.13 и 7.4) показан типичный вид хлоропластов в клетке мезофилла. На рис. 7.6 и 7.8 показаны электронные микрофотографии хлоропластов, а на рис. 7.7 схема строения хлоропласта и его мембранных систем. На мембранах протекают световые реакции фотосинтеза (разд. 7.6.2). Здесь расположены хлорофилл и другие пигменты, ферменты и переносчики электронов. Система состоит из множества заполненных жидкостью плоских мешочков, называемых тилакоидами; тилакоиды образуют стопки, или граны, которые соединены друг с другом ламеллами (одиночными гранами). Каждая грана напоминает стопку монет, а ламелла — пластинку (рис. 7.8). В световом микроскопе граны едва видны в виде мелких зерен.
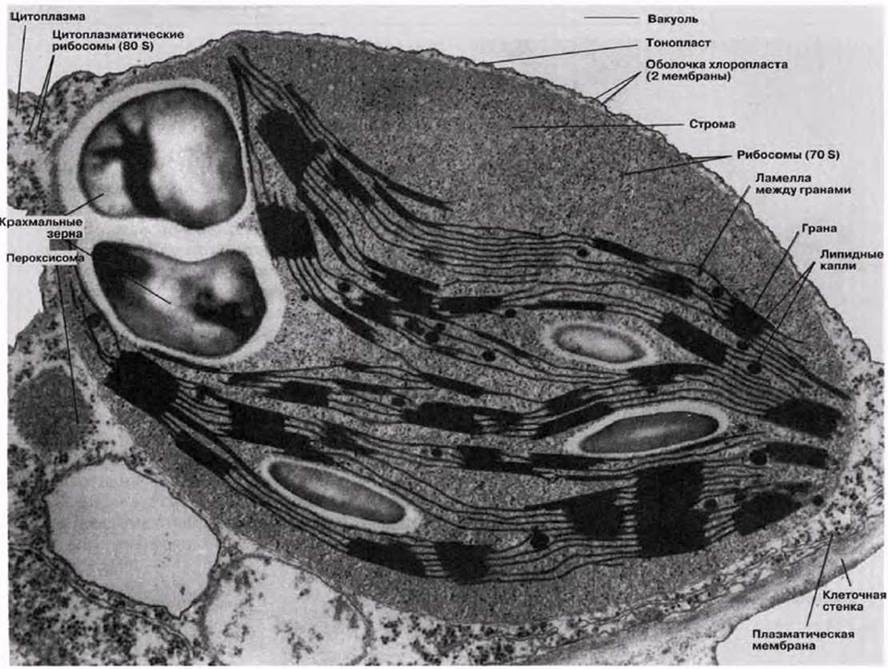
Рис. 7.6. Электронная микрофотография хлоропласта (х15 800).
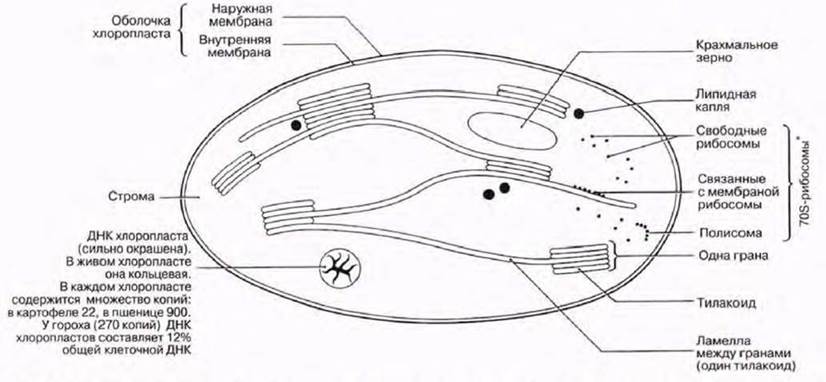
Рис. 7.7. Строение хлоропласта. Для упрощения рисунка система мембран несколько сокращена по своим размерам (* — белоксинтезирующий аппарат, подобный прокариотическому).
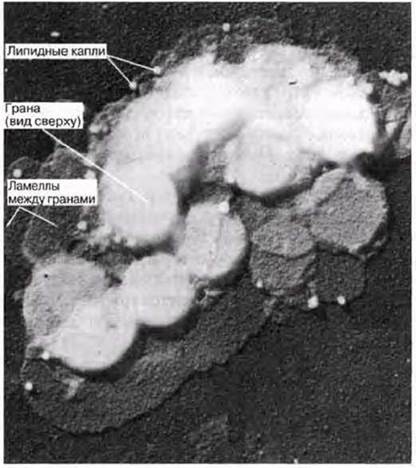
Рис. 7.8. Электронная микрофотография «голого» хлоропласта (с удаленной наружной оболочкой), полученная с помощью сканирующего электронного микроскопа. Вид сверху. Ламеллы и граны показаны в трехмерном изображении. Обратите внимание, что ламеллы плоские, как листы бумаги, и соединяют между собой граны. Для приготовления препарата использовалась реплика с напылением.
В строме происходят темповые реакции фотосинтеза (разд. 7.6.3). Структура стромы напоминает гель; в ней содержатся растворимые ферменты, в частности ферменты цикла Кальвина, а также сахара и органические кислоты. Избыток углеводов, образуемых в процессе фотосинтеза, запасается в виде крахмальных зерен. С мембранами часто связаны сферические липидные капли. Они становятся крупнее по мере разрушении мембран в процессе их старения. По-видимому, в этих каплях аккумулируются липиды из мембран.
Белоксинтезирующий аппарат и теория эндосимбиоза
Интересной особенностью хлоропластов помимо фотосинтеза, является их белоксинтезирующий аппарат. В шестидесятых годах XX в. было показано, что и хлоропласты, и митохондрии содержат ДНК и рибосомы. Это навело на мысль, что хлоропласты и митохондрии, возможно, являются прокариотическими организмами, внедрившимися в эукариотическую клетку на ранних этапах развития жизни. Таким образом, в соответствии с эндосимбиотической теорией эти органеллы представляют крайнюю форму симбиоза. Некоторые данные в пользу этой теории приведены в табл. 7.2.
Таблица 7.2. Сравнение прокариот, хлоропластов и митохондрий с эукариотами
Прокариоты, хлоропласты, митохондрии |
Эукариоты |
|
ДНК |
Кольцевая Не организована в хромосомы Не находится в ядре |
Линейная Организована в хромосомы Находится в ядре |
Рибосомы |
Небольшие (70 S) |
Крупные (80 S) |
Устойчивость к антибиотикам |
Синтез белка ингибируется хлорамфеннколом, но не циклогексимилом |
Синтез белка ингибируется циклогексимидом, но не хлорамфеннколом |
Средний диаметр |
Прокариотическая клетка: 0,5—10 мкм Хлоропласт: 1—10 мкм Митохондрия: 1 мкм |
Эукариотическая клетка: 10-100 мкм |
Фотосинтезирующие бактерии (прокариоты) не содержат хлоропластов. Их фотосинтетические пигменты расположены на мембранах, разбросанных по цитоплазме. Таким образом, целая клетка становится похожей на один хлоропласт, причем она имеет практически такие же размеры. В настоящее время полагают, что хлоропласты являются потомками фотосинтезирующих бактерий (разд. 2.6.1).
Хлоропласты и митохондрии, как было показано, действительно могут синтезировать некоторые собственные белки. При этом для выполнения этой задачи некоторые гены должны переместиться в ядро клетки, где они взаимодействуют с ядерной ДНК. Этим объясняется тот факт, что ни хлоропласты, ни митохондрии не могут более существовать независимо.