БИОЛОГИЯ Том 1 - руководство по общей биологии - 2004
10. ОРГАНИЗМЫ И ОКРУЖАЮЩАЯ СРЕДА
10.3. Экосистемы и поток энергии
10.3.5. Эффективность переноса энергии: продуктивность
Изучение продуктивности различных компонентов экосистемы фактически означает прослеживание различных путей переноса в ней энергии. Энергия поступает в биоту через продуцентов, и скорость, с которой она запасается ими в виде пригодного в пищу органического вещества, называется первичной продуктивностью. Этот параметр очень важен, поскольку он определяет общее количество энергии, доступное всем организмам сообщества, т. е. суммарную биомассу экосистемы.
10.7. Какие еще группы организмов, кроме растений, обеспечивают первичную продуктивность экосистемы?
Как указывалось в разд. 10.3.1, количество солнечной радиации, достигающей поверхности Земли, зависит от географической широты местности и особенностей ландшафта, например высоты над уровнем моря и экспозиции участка. Количество этой радиации, ассимилируемое растениями, определяется ее качеством и структурой растительного покрова. В Британии среднее количество энергии, достигающее растений, соответствует примерно 1 · 106 кДж · м-2 · год-1. Из этого количества 95—99% сразу же теряется за счет отражения, нагревания и испарения. Остающаяся лучистая энергия (1—5%) поглощается хлорофиллом и используется для синтеза органических молекул. Накопленная растениями суммарная химическая энергия называется валовой первичной продукцией (ВПП). Примерно 20—25% ВПП расходуется самим растением на дыхание и фотодыхание, а остальное дает прирост его биомассы, называемый чистой первичной продукцией (ЧПП). Именно эта энергия потенциально доступна следующему трофическому уровню.
Потребляя другие организмы, гетеротрофы переносят вещество и энергию с одного трофического уровня на другой. Однако не вся пища идет на увеличение биомассы консументов. Часть энергии утрачивается в процессе дыхания; к числу других потерь относятся конечные продукты метаболизма (экскреты); кроме того, корм переваривается не полностью и эта непереваренная часть утрачивается в виде фекалий (экскрементов).
Потребленная пища = Рост + Дыхание + Экскременты + Экскреты
Впрочем, если брать экосистему в целом, то отходов в ней не образуется: то, что выделяют одни, служит пищей для других — редуцентов и детритофагов.
Некоторые из членов приведенного равенства нетрудно измерить у домашних животных или в лаборатории у диких животных. Рост измеряется как увеличение биомассы, или лучше, как увеличение энергетической ценности организма в единицу времени. Количество потребленных за этот же период кормов, а также образовавшихся фекалий и мочи легко взвесить. Зная эти величины, можно простыми арифметическими действиями рассчитать, сколько энергии затрачено на дыхание.
То, что остается гетеротрофам после всех неизбежных потерь и обеспечивает их рост, восстановление тканей и размножение, называется вторичной продукцией (независимо от трофического уровня).
На рис. 10.10 ясно показано, что энергия теряется каждым звеном пищевой цепи, и длина ее, очевидно, зависит от масштабов этих потерь. Наименее эффективен первый этап — преобразование солнечной радиации в первичную продукцию. В дальнейшем при переходе энергии с одного трофического уровня на другой потери энергии намного меньше. Средняя эффективность ее переноса от автотрофов к фитофагам близка к 10%, а от одного животного к другому — к 20%. В целом растительноядные хуже усваивают пищу, чем хищники, поскольку растения содержат много целлюлозы и других веществ, которые обычно не перевариваются и выводятся из организма в составе фекалий, не превращаясь во вторичную продукцию.
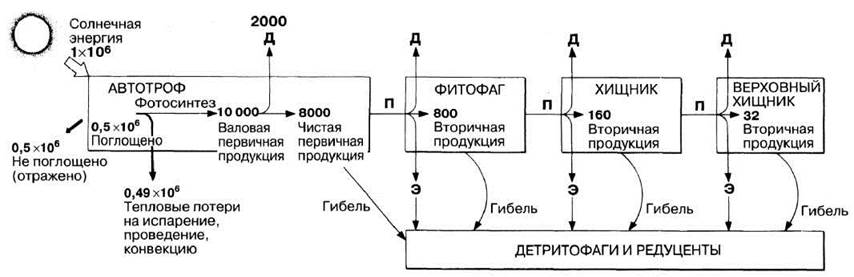
Рис. 10.10. Поток энергии в пастбищной пищевой цепи. Все величины выражены в кДж · м-2 · год-1. Д — потери энергии при дыхании; Э — потери энергии при переходе из пастбищной пищевой цепи в детритную с экскретами (моча и т. п.) и экскрементами. П — потребление организмами более высокого трофического уровня.
Энергия, затраченная на дыхание, уже недоступна другим организмам. Однако та ее часть, которая соответствует экскретам и фекалиям, для экосистемы не потеряна: ее получают редуценты и детритофаги. Сходным образом любой мертвый организм, опавшие листья, ветки и сучья представляют собой основу для возникновения новых детритных пищевых цепей. Детритные цепи часто очень сложны и гораздо менее изучены, чем традиционно рассматриваемые пастбищные. Тем не менее в смысле передачи энергии они зачастую имеют более важное значение.
Доля ЧПП, поступающей непосредственно в детритную пищевую цепь, зависит от типа экосистемы. В лесу основная масса первичной продукции поступает в детритную пищевую цепь, а не в пастбищную: опад, подстилка и перегной в почве — основные зоны активности лесных гетеротрофов, хотя эти организмы, как правило, незаметны. А вот в приповерхностном слое океана или на интенсивно используемом пастбище в пастбищную пищевую цепь может поступать более половины ЧПП. В самых интенсивных сельскохозяйственных системах игнорируется потенциальная роль производства продовольствия на основе детрита.
Цифры, приводимые в этом разделе, рассчитаны для периода в один год. Если экосистема равновесна, т. е. не меняется в ходе сукцессии (разд. 10.6.1), то ее общая биомасса по истечении такого срока остается прежней. Вся энергия, перешедшая в первичную продукцию, поступает на следующие трофические уровни и по пути рассеивается, так что ЧПП не остается. Однако такое равновесное состояние экосистем — большая редкость. Например, молодой лес к концу года подрастает, т. е. запасает часть солнечной энергии в биомассе деревьев. Годовой период как основу для оценки энергетических аспектов экосистемы удобно использовать потому, что год охватывает все сезонные изменения, происходящие в экосистеме. Например, первичная продуктивность, как правило, повышается весной и летом, когда активизируется рост автотрофов, а потом начинает преобладать вторичная продуктивность.
Одна из причин пристального внимания к энергетике экосистем состоит в том, что эта область экологии очень тесно связана с получением людьми пищи и топлива. Она позволяет анализировать эффективность сельскохозяйственных систем и предлагать пути их совершенствования. Поскольку энергия теряется на каждом трофическом уровне, очевидно, что для всеядных животных, вроде человека, вегетарианство — более рентабельный способ пищевой эксплуатации экосистем (табл. 10.2). Однако, при анализе эффективности питания следует учитывать и другие факторы. Например, животный белок обычно дает нам больше незаменимых аминокислот, чем растительный, хотя ряд бобовых, в частности соя, ненамного отстают от него в этом плане. Кроме того, животные ткани легче перевариваются, поскольку клетки их не окружены, как у растений, стенками, которые прежде необходимо разрушить. Наконец, во многих экосистемах животные концентрируют в себе вещество, потребляемое на обширной территории, не пригодной для земледелия. Примерами тому служат — выпас скота на низкокачественных пастбищах (овец в Британии, северных оленей в Скандинавии, канн в Восточной Африке) или рыболовство.
Таблица 10.2. Эффективность сельскохозяйственных пищевых цепей в Великобритании
Пищевая цепь |
Пример |
Энергоотдача для человека, кДж · 103 га-1 |
Выход белка для человека, кг · га-1 · год-1 |
А. Зерновые → человек |
Монокультуры пшеницы или ячменя |
7800-11000 |
42 |
Б. Зерновые → скот → человек |
Откорм ячменем бычков или беконных свиней |
745-1423 |
10-15 |
В. Интенсивно используемое пастбище →скот → человек |
Интенсивный выпас крупного рогатого скота на окультуренном пастбище |
||
Мясо |
339 |
4 |
|
Молоко |
3813 |
46 |
|
Г. Пастбище и зерновые → скот → человек |
Молочная ферма стойлово-пастбищного типа |
||
Молоко |
1356 |
17 |
|
Данные из: Duckham A. N., Mansfield G. В. (1970) Farming systems of the world, Chatto and Windus. |
|||