БИОЛОГИЯ Том 1 - руководство по общей биологии - 2004
10. ОРГАНИЗМЫ И ОКРУЖАЮЩАЯ СРЕДА
10.7. Популяционная экология
10.7.3. Увеличение размеров (рост) популяции и кривые роста
Процессы увеличения и уменьшения размеров популяции подчиняются определенным законам. Увеличение размеров определяется репродуктивным (биотическим) потенциалом организмов и сопротивлением среды. Максимальный репродуктивный потенциал — это теоретическая скорость размножения при неограниченной доступности ресурсов среды. Он зависит от возрастной структуры популяции и соотношения в ней самцов и самок (в случае полового размножения).
Сопротивление среды соответствует всей совокупности лимитирующих (т. е. ограничивающих возможности организмов) биотических и абиотических факторов, которые препятствуют реализации максимального репродуктивного потенциала. Сюда относятся как внешние по отношению к популяции воздействия (хищничество, доступность пищи, тепло, свет, пространство и т. п.), так и ее внутренние регулирующие механизмы, например внутривидовая конкуренция и территориальность. Все эти факторы связаны механизмами отрицательной обратной связи. Например дефицит какого-либо ресурса (скажем, пищи) обостряет внутривидовую конкуренцию, а это сокращает популяцию, приводя к тому, что его снова хватает всем выжившим.
Равновесие между биотическим потенциалом и сопротивлением среды соответствует так называемой несущей емкости среды, т. е. количеству особей данного вида, способных выжить при существующей доступности ресурсов.
Кривые роста
Выделяют две основные формы кривых роста — чисто экспоненциальную (J-образную) и сигмоидную (S-образную).
Сигмоидная, или S-образная, кривая описывает ситуацию, при которой в новом для популяции местообитании ее плотность сначала возрастает медленно (лаг-фаза, соответствующая периоду адаптации к условиям), а затем быстро, почти экспоненциально. По прошествии некоторого времени скорость роста замедляется и становится в конечном итоге нулевой: рождаемость полностью уравновешивается смертностью (рис. 10.19, А). Говорят, что кривая выходит на плато. Замедление роста популяции объясняется увеличением внутривидовой конкуренции за ресурсы, например пищу или места гнездования. В результате по механизму отрицательной обратной связи повышается смертность особей и замедляется их размножение (меньший процент спаривающихся животных, рост числа вызванных стрессом выкидышей и т. п.). Другими словами, растущее сопротивление среды уравновешивает биотический потенциал.
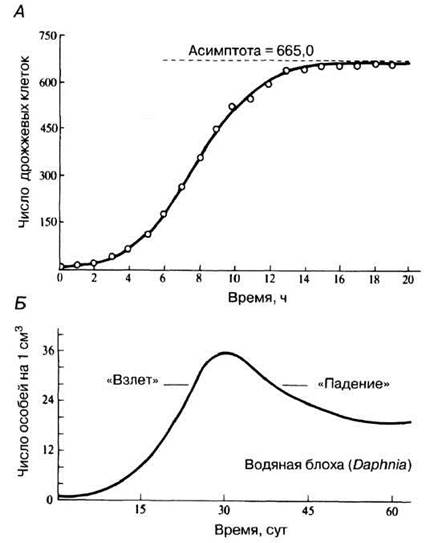
Рис. 10.19. Типы кривых роста популяции. А. Сигмоидная (S-образная) кривая роста культуры дрожжей. Простой случай, когда сопротивление среды (здесь — накопление метаболитов, выделяемых самими организмами) прямо пропорционально плотности популяции. Б. Кривая, соответствующая росту популяции дафнии (водяной блохи) в культуре. Такая форма соответствует периодам «взлета» и «падения» популяционной плотности.
Такой тип популяционного роста называют зависимым от плотности, поскольку для данного набора ресурсов скорость роста определяется числом особей в ограниченном пространстве, занимаемом популяцией. Обилие организмов, соответствующее выходу кривой на плато (нулевая скорость роста), — это максимальная несущая емкость среды для данного вида.
S-образные кривые роста численности свойственны популяциям многих микроорганизмов, растений и животных как в лабораторных, так и в полевых условиях. Наглядным примером может служить рост бактерий на свежей культуральной среде (рис. 12.8). S-образный рост численности фитопланктона может наблюдаться весной в озерах и океанах. Он наблюдается также у насекомых (например, мучных хрущаков) и клещей, интродуцированных в новое местообитание с изобилием пищи и отсутствием хищников.
Экспоненциальный рост без выхода на плато (J-образная кривая) соответствует ситуации, при которой после начального адаптационного периода (лаг-фазы) численность особей резко возрастает, но затем рост внезапно прекращается, когда начинает проявляться отсутствовавшее прежде сопротивление среды (рис. 10.19, Б). Такой рост популяции называют независимым от плотности, поскольку он ничем не сдерживается до последнего момента, за которым следует массовая гибель особей. Эта гибель в свою очередь может вызываться либо наступлением неблагоприятного сезона, либо окончанием сезона размножения самих организмов или их основных жертв. Иногда гибель запрограммирована в жизненном цикле особей на момент, непосредственно следующий за размножением (у однолетних растений, лососевых), или вызывается внешним вмешательством, например инсектицидной обработкой посевов для уничтожения насекомых-вредителей. В целом для таких популяций типичны циклические «взлеты» и «падения» численности (демографические взрывы и катастрофы), наблюдаемые, скажем, у некоторых прожорливых насекомых-фитофагов или связанные с водорослевыми «цветениями».