БИОЛОГИЯ Том 1 - руководство по общей биологии - 2004
10. ОРГАНИЗМЫ И ОКРУЖАЮЩАЯ СРЕДА
10.7. Популяционная экология
10.7.5. Межвидовые взаимодействия, влияющие на размеры популяций
Исследования популяционной динамики в природных условиях редко удается ограничить пределами одного вида, поскольку он практически всегда взаимодействует с другими видами. Такие взаимодействия называют межвидовыми. Между организмами, относящимися к разным трофическим уровням, действуют отношения хищник-жертва и паразит—хозяин. Возможны и более тонкие взаимосвязи, приносящие выгоду одному или сразу обоим партнерам, — комменсализм и мутуализм. На одном и том же трофическом уровне возможна межвидовая конкуренция, например за пищу и пространство. Иногда она приводит к разделению ресурсов.
Отношения хищник-жертва
Простая модель таких взаимоотношений хорошо иллюстрируется лабораторным экспериментом с двумя видами клещей — хищником Typhlodromus и фитофагом Eotetranychus. На рис. 10.21 показаны циклические колебания их численности, несколько не совпадающие по фазе.
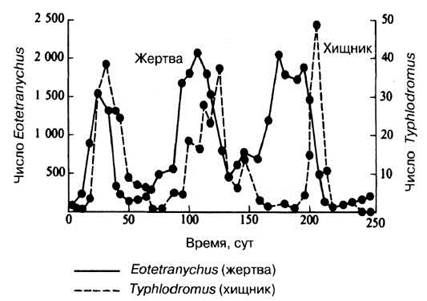
Рис. 10.21. Колебания численности популяций хищного клеща Typhlodromus и его жертвы — растительноядного клеща Eotetranychus в лабораторных условиях. (Из М. К. Sands (1978) Problems in ecology, Mills, Boon Limited.)
Объясняются эти кривые легко: рост числа жертвы, т. е. корма, приводит к усиленному размножению его потребителей — хищников («демографическому взрыву»). Это в свою очередь усиливает гибель фитофагов и вызывает их «демографическую катастрофу», за которой неизбежно следует гибель от бескормицы и хищников. Цикл считается завершенным, когда сниженное число хищников позволяет популяции жертвы снова расти. Каждый такой цикл, очевидно, охватывает несколько поколений.
Хотя хищничество — лишь один из факторов, регулирующих размеры популяций, его важная роль несомненна. Некоторые данные о его значении и благоприятном воздействии на популяции жертв в долгосрочной перспективе получены на плато Кейбаб в Аризоне. В 1906 г. там был создан заповедник, и, чтобы сохранить в нем популяцию оленей, в течение следующих 30 лет проводился отстрел его главных врагов — пум, волков, и койотов. До 1906 г. популяция оленей была стабильной (примерно 4000 голов), а затем, как показано на рис. 10.22, произошел ее демографический взрыв, приведший к превышению «емкости» наличных пастбищ (по оценкам, она соответствовала примерно 30 000 голов). Численность оленей росла до 1924 г. экспоненциально, не выходя на плато, и достигла, по учетным данным, 100 000 особей. Затем недостаток корма и болезни привели к демографической катастрофе. Вместе с тем перевыпас снизил продуктивность пастбищ, и, несмотря на резкое падение нагрузки, их емкость так и не вернулась к уровню 1906 г., составив в конечном итоге 10 000 голов.
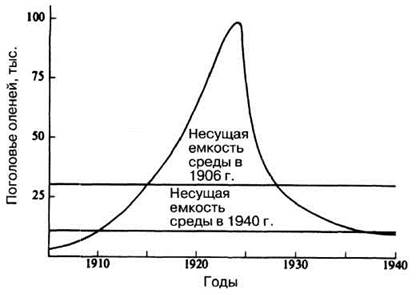
Рис. 10.22. Кривая роста популяции оленей на плато Кейбаб после истребления там хищников.
Отношения паразит-хозяин
Некоторые изученные популяции паразитов и их хозяев (например, при паразитировании одних насекомых на других) демонстрируют сходные, но слегка не совпадающие по фазе циклы.
Межвидовая конкуренция
Конкуренция между популяциями внутри экосистемы может происходить за любые доступные ресурсы — пищу, свет, пространство, убежище и т. п. Если два вида располагаются на одном трофическом уровне и питаются одними и теми же организмами (растительными или животными), то они скорее всего будут конкурентами. С течением времени может произойти пищевая специализация одного или обоих этих видов, так что в рамках одного трофического уровня они займут разные ниши. Этот процесс, сводящий к минимуму конкуренцию, называется разделением ресурсов (см. ниже). Если конкуренты занимают одну и ту же нишу, или ниши сильно перекрываются, то возможны два исхода: либо установится равновесие по типу «боевой ничьей», либо популяция одного из видов окажется менее конкурентоспособной и постепенно снизит свою численность вплоть до полного исчезновения (вымирания). Последний вариант называется конкурентным исключением.
В природных условиях межвидовую конкуренцию изучать сложно, но ее существование было подтверждено классическими работами русского биолога Г. Ф. Гаузе, в 1934 г. работавшего с несколькими видами инфузорий рода Paramecium. Некоторые из его результатов проиллюстрированы на рис. 10.23.
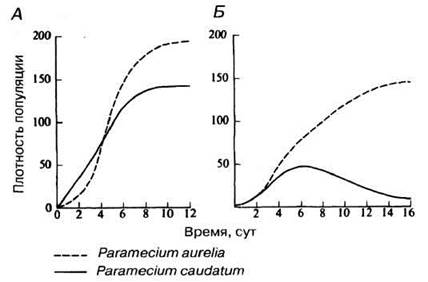
Puc. 10.23. Рост численности популяций двух видов Paramecium: А — при раздельном культивировании; Б — в смешанной культуре.
10.17. Изучив рис. 10.23, ответьте на следующие вопросы:
а) какой тип кривой роста популяции характерен для видов, находящихся в изоляции друг от друга?
б) за какие ресурсы могут конкурировать эти два вида в смешанной культуре?
в) какие факторы обеспечивают Р. aurelia конкурентное преимущество над Р. caudatum?
Когда виды Р. aurelia и Р. caudatum живут вместе, первый из них питается активнее, в результате чего через пять суток численность Р. caudatum начинает снижаться, а спустя примерно 20 суток этот вид полностью исчезает из смешанной культуры («вымирает»). Это и есть так называемое конкурентное исключение. Однако популяция Р. aurelia достигает стационарной фазы роста медленнее, чем в изоляции, так что тоже страдает от конкуренции, несмотря на свои преимущества. Это помогает объяснить давление отбора, заставляющее оба вида разделять ниши. В природных условиях полного вымирания менее конкурентоспособного вида обычно не происходит, вид просто становится редким.
Принцип конкурентного исключения, или принцип Гаузе, с тех пор был подтвержден в экспериментах на различных животных и растениях, например в смешанных культурах разных видов ряски (Lemna).
Изучение конкурентного исключения в природных популяциях затрудняется из-за большого числа взаимодействующих видов, а также из-за колебаний таких переменных, как температура, влажность и доступность пищи, что неизбежно отражается на относительной конкурентоспособности рассматриваемых организмов и не позволяет установиться устойчивому равновесию.
Большое внимание привлекает особая форма конкуренции, связанная с выделением некоторыми видами органических веществ, замедляющих рост других организмов. У растений это взаимодействие называется аллелопатией. Такая химическая борьба особенно развита среди микроорганизмов, выделяющих различные антибиотики и ингибиторы роста, чтобы «приватизировать» питательный субстрат. Хорошо известный пример — пенициллин, образуемый плесневыми грибами рода Penicillium и подавляющий развитие грамположительных бактерий (разд. 12.11.1).
Разделение ресурсов
Чем более специализированную экологическую нишу занимает организм, тем меньше у него шансов встретить серьезных конкурентов. В давно сложившихся, богатых видами сообществах эволюция (коэволюция) привела к разделению ресурсов (ниш), т. е. к специализации его членов на использовании ресурсов, в которых по тем или иным причинам слабо нуждаются другие. Это повышает как устойчивость экосистемы, так и ее продуктивность.
Разделение ресурсов происходит разными путями, например за счет:
1) морфологической и поведенческой специализации на питании только определенным типом корма; так, клювы разных птиц своей формой адаптированы к ловле на лету насекомых, долблению стволов, раскалыванию орехов, разрыванию мяса и т. д.;
2) разделения местообитания по вертикали (стратификации, см. рис. 10.15): одни виды живут в пологе леса, другие в лесной подстилке;
3) разделения местообитания по горизонтали, т. е. распределения по разным микроместообитаниям — ложбинам, кочкам, прогалинам и т. д.
Однако, несмотря на тенденцию каждого вида к специализации, некоторое перекрывание ниш и связанная с этим межвидовая конкуренция все равно сохраняются.