БИОЛОГИЯ Том 2 - руководство по общей биологии - 2004
16. КООРДИНАЦИЯ И РЕГУЛЯЦИЯ У РАСТЕНИЙ
16.2. Ростовые вещества растений
16.2.2. Ауксины и геотропизм
Нетрудно заметить, что корням свойствен положительный геотропизм, т. е. они растут вниз, а побегам (по крайней мере главным) — отрицательный, т. е. для них характерна тенденция расти вверх. То, что стимулом, вызывающим такую реакцию, является земное тяготение, или гравитация, можно продемонстрировать с помощью прибора, называемого клиностатом (рис. 16.11). При вращении камеры сила тяжести одинаково и поочередно воздействует на все части проростков. Чтобы избежать одностороннего действия силы тяжести и заставить побеги и корни расти горизонтально, достаточно четырех оборотов камеры в час. Неподвижные контрольные проростки сохраняют нормальный ответ на гравитационную силу, т. е. побег у них растет вверх, а корни вниз. На протяжении всего опыта очень важно обеспечить равномерное освещение со всех сторон (или проводить опыт в темноте), исключив таким образом направленную реакцию на свет.

Рис. 16.11. Клиностат с проростками конских бобов, которые вращались в нем несколько суток.
Роль ауксинов в геотропизме можно продемонстрировать в эксперименте, представленном на рис. 16.12 с использованием методики, предложенной Вентом. Ауксин движется от расположенной горизонтально верхушки колеоптиля, но по пути «стекает» вниз. Повышение концентрации ауксина на нижней стороне интактного колеоптиля приведет к усиленной стимуляции растяжения клеток в этом месте, и в результате рост колеоптиля будет направлен вверх.
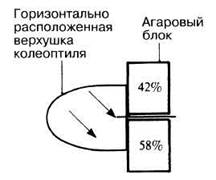
Рис. 16.12. Влияние гравитации на движение ауксина в горизонтально расположенной верхушке колеоптиля. Распределение фитогормона (%) изучали с помощью биотеста Вента.
Удаление кончика корня делает его нечувствительным к силе тяжести, однако в данном случае продемонстрировать движение ауксина труднее из-за его очень низкой концентрации в корне, что не позволяет получить убедительных результатов в биотестах, подобных описанному выше. Тем не менее весьма интересные результаты получены в опыте, показанном на рис. 16.13.

Рис. 16.13. Влияние асимметричного распределения ауксина на рост декапитированных колеоптилей и корней.
Подобного рода наблюдения привели к созданию гипотезы, схематично представленной на рис. 16.14. В соответствии с этой гипотезой противоположные ростовые реакции корней и побегов на ауксин обусловлены их разной чувствительностью к нему. Несколько позже мы обсудим различные модификации этой гипотезы с учетом данных, полученных в последние годы.

Рис. 16.14. Гипотеза, объясняющая перераспределение ауксина в горизонтально расположенном проростке.
Иной чувствительностью корней к ауксину (см. рис. 16.10) можно также объяснить наблюдаемый у них в ряде случаев отрицательный фототропизм. Повышенное накопление ауксина на затененной стороне подавляет рост, и в результате этого клетки на освещенной стороне растягиваются быстрее, заставляя корень удаляться от света. Таким образом, был выявлен важный аспект гуморальной регуляции роста растений: реакция зависит не только от природы фитогормона (качественный контроль), но и от его концентрации (количественный контроль).
Механизм гравитационной чувствительности
Четко выраженный геотропизм у растений заставил ученых еще до открытия ауксинов задумываться над механизмом такой реакции. Еще Дарвин показал, что удаление корневого чехлика — группы крупных паренхимных клеток, которые защищают кончик корня во время роста в почве, подавляет его положительный геотропизм. На срезе корневого чехлика в его клетках видны крупные крахмальные зерна, лежащие в а мило пластах (бесцветных пластидах) (рис, 16.15).
Еще в 1900 г. было высказано предположение, что эти клетки ведут себя как статоциты, т. е. как рецепторы гравитации, а крахмальные зерна — играют роль статолитов — структур, перемещающихся под действием силы тяжести. Согласно так называемой гипотезе крахмальных статолитов, крахмальные зерна оседают в нижней части клеток (рис. 16.15). Это каким-то образом влияет на распределение ростовых веществ, которые иногда образуются в кончике корня, иногда — в корневом чехлике, а иногда — в обеих этих структурах одновременно. В пользу такой гипотезы говорит многое. Все чувствительные к гравитации органы растения содержат статоциты. Они присутствуют, например, в обкладке проводящих пучков стеблей (крахмалоносное влагалище). Растения, у которых тем или иным способом разрушены крахмальные зерна, утрачивают гравитропизм, однако он восстанавливается, если дать им возможность накопить крахмал.

Рис. 16.15. Электронная микрофотография среза корневого чехлика; видны скопившиеся в нижней части клеток амилопласты, содержащие крахмальные зерна.
16.11. В чем сходны механизмы восприятия силы тяжести у растений и животных?
Современные гипотезы гравитропизма
Как сказано выше, гравитропизм колеоптилей, по-видимому, опосредован действием ауксинов, но у большинства побегов геотропическая реакция наблюдается даже после удаления верхушки, и поэтому до сих пор неясно, связана ли она с движением ауксинов. В корнях действительно происходит перераспределение ауксинов, но оно, по всей видимости, не настолько велико, чтобы полностью объяснить наблюдаемые изменения скорости роста. Было показано, что какой-то ингибитор и в самом деле транспортируется из корневого чехлика в зону растяжения, но это может быть и не обязательно ауксин. Несколько групп ученых не смогли обнаружить ауксин в корневых чехликах кукурузных проростков, зато выделили из них хорошо известный ингибитор роста абсцизовую кислоту. Возможно, какую-то роль играет еще один ингибитор роста — этилен. И наконец, было обнаружено, что концентрация гиббереллинов (стимуляторов роста) на быстро растущей стороне стеблей и корней при геотропической реакции намного превышает норму.
16.12. Какой вывод можно сделать из экспериментов, проиллюстрированных рис. 16.16? В контроле (с необработанным агаром) изгиба не наблюдается. Если вместо абсцизовой кислоты использовали ИУК, заметного изгиба тоже не было.
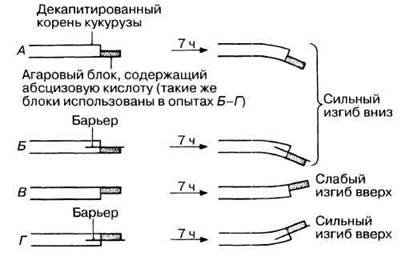
Рис. 16.16. Влияние абсцизовой кислоты на геотропизм декапитированных корней (данные экспериментов Pilet, 1975).