БИОЛОГИЯ Том 3 - руководство по общей биологии - 2004
26. ЭВОЛЮЦИЯ, ИЛИ ИСТОРИЯ ЖИЗНИ НА ЗЕМЛЕ
26.7. Данные в пользу теории эволюции
Свидетельства, подтверждающие современные представления об эволюции, поступают из разных источников, среди которых главное место занимают данные:
1) палеонтологии,
2) биогеографии,
3) систематики,
4) селекции растений и животных,
5) сравнительной анатомии,
6) адаптивной радиации,
7) сравнительной эмбриологии,
8) сравнительной биохимии.
Большая часть данных, приводимых в настоящей главе, не была доступна Дарвину и Уоллесу в то время, когда они публиковали свои статьи о происхождении видов, что, однако, не помешало Дарвину использовать свою интуицию, как это свойственно великим ученым. Об этом свидетельствует следующее его высказывание:
«В октябре 1838 г., т. е. спустя 15 месяцев после того как я начал свое систематическое исследование, я взялся читать просто для развлечения работу Мальтуса о народонаселении и поскольку длительные наблюдения над жизнью животных и растений вполне подготовили меня к тому, чтобы оценить происходящую повсеместно борьбу за существование, я вдруг понял, что в таких условиях благоприятные изменения должны сохраняться, а неблагоприятные отметаться. Результатом всего этого должно быть формирование новых видов. Итак, я, наконец, получил теорию, с которой можно работать.»
Представленные здесь данные в значительной мере подтверждают теорию эволюции путем естественного отбора, изложенную в разд. 26.5; следует, однако, помнить, что они не содержат никаких доказательств того, что эволюция происходила именно таким путем, равно как и доказательств того, что в этом не участвуют никакие другие механизмы. Эти данные получены в разных областях знания, и их интерпретация во многом зависит от допущения достоверности принятой здесь концепции эволюции. Немало среди них и таких свидетельств, которые в свою очередь требуют подтверждения, а также исключений или сведений, которые можно интерпретировать иначе. Однако концепция эволюции в широком смысле основана на огромном количестве научных фактов. Читая последующие разделы, постарайтесь сами оценить имеющиеся данные и решить, оправданы ли сделанные на их основе заключения. Попытайтесь провести различие между данными, свидетельствующими в пользу эволюции, и данными, свидетельствующими о том, что механизмом эволюции служит естественный отбор. Остальная часть этой главы посвящена главным образом доказательствам существования эволюции, а в гл. 27 представлены данные, свидетельствующие в пользу естественного отбора.
26.7.1. Палеонтология
Палеонтология занимается изучением ископаемых остатков, т. е. любых сохранившихся остатков, предположительно принадлежавших каким-либо живым организмам. Это могут быть целые организмы, твердые скелетные структуры, наружные и внутренние ядра, отпечатки, следы и копролиты (окаменевшие экскременты) (табл. 26.3.).
Таблица 26.3. Типы ископаемых остатков, их образование и примеры
Тип остатков |
Процесс фоссилизации |
Примеры |
Целые организмы |
Вмерзшие в лед в период оледенения |
Мамонты, найденные в вечной мерзлоте в Сибири |
«-» |
Заключенные в янтарь — отвердевшую смолу хвойных деревьев |
Экзоскелеты насекомых в олигоценовых отложениях на Балтийском побережье |
«-» |
Заключенные в асфальт |
«Мумии», найденные в асфальтовых озерах Калифорнии |
«-» |
Захороненные в кислых торфяных болотах, где отсутствие бактерий и грибов препятствует полному разложению |
«Мумии», найденные в болотах и торфяниках на Скандинавском полуострове |
Твердые скелетные структуры |
Захороненные в осадочном песке и глинах, образующих осадочные породы (например, известняк, песчаник и пр.) |
Кости, раковины и зубы (очень часто встречаются на Британских островах) |
Наружные и внутренние ядра |
Твердые объекты, захороненные как указано выше. При литификации осадка части скелета растворяются, оставляя отпечаток своей наружной или внутренней поверхности. Полученная полость может заполниться тонкозернистым материалом, который затвердевает образуя слепок. При этом возможно сохранение мельчайших деталей |
Брюхоногие из портландского камня (юра). Отпечатки гигантских хвощей (каламитов), росших в лесах каменноугольного периода. Отпечатки внутренних поверхностей раковин моллюсков, на которых видны места прикрепления мышц |
Окаменелости |
Постепенное замещение приносимыми водой минеральными веществами такими, как кремнезем, пириты, карбонат кальция или углерод. Медленное замещение этими веществами мягких частей по мере их разложения обеспечивает сохранение тонких структур организма. |
Иглокожее Micraster, структуры тела которого были замещены кремнеземом |
Отпечатки |
Отпечатки организмов или их частей на поверхности тонкозернистых отложений, на которых они погибли |
Перья археоптерикса из верхней юры. Медузы из кембрия Британской Колумбии. Отпечатки листьев из карбона |
Следы |
Следы ног животных, следы ползания и ходы, оставленные в иле, быстро высыхали и заполнялись песком, а затем покрывались другими отложениями |
Следы ног и хвоста динозавра дают представление о размерах и позе животного |
Копролиты |
Экскременты животных, избежавшие разложения, а затем включившиеся в осадочные породы. Часто содержат остатки съеденной пищи, например зубы или чешуи |
Экскременты кайнозойских млекопитающих |
Одних лишь палеонтологических данных недостаточно для обоснования того, что эволюция действительно происходила, однако они подтверждают факт прогрессивного возрастания сложности организмов. Ископаемые остатки были хорошо известны еще до того, как эволюция получила всеобщее признание. Их считали либо остатками существ, сотворенных раньше других, либо артефактами, помещенными в горные породы Богом. Большую часть найденных до сих пор остатков можно распределить по тем же самым таксономическим группам (типам и классам), что и ныне живущие виды, однако вопрос о том, являются ли они предками современных форм, можно обсуждать, но нельзя доказать.
В самых древних породах, содержащих ископаемые остатки, встречаются организмы лишь нескольких типов и все они имеют простое строение. Более молодые породы содержат более разнообразные ископаемые остатки со все более сложным строением. Во всей палеонтологической летописи многие виды, появляющиеся на одном из ранних стратиграфических уровней (уровень в отложениях горных пород) исчезают на более позднем уровне. В эволюционном смысле это истолковывают как возникновение и исчезновение видов в соответствующие эпохи.
Судя по имеющимся данным, на протяжении истории Земли на ней происходили изменения географических областей и климатических условий. Поскольку каждый организм приспособлен к определенной среде, непрерывно изменяющиеся условия могли благоприятствовать возникновению некоего механизма эволюционных изменений, что позволяет объяснить прогрессивные изменения в строении организмов, отраженные в палеонтологической летописи. Экологические соображения тоже согласуются с палеонтологическими данными. Например, растения появились на суше раньше, чем животные, а насекомые — раньше, чем опыляемые ими растения.
Одно из главных возражений против использования ископаемых остатков в качестве свидетельств эволюции — отсутствие непрерывности в палеонтологической летописи. Разрывы в летописи («недостающие звенья») считают веским доводом против теории образования новых форм путем постепенного изменения. Однако существует ряд соображений, позволяющих объяснить неполноту палеонтологической летописи:
1) мертвые организмы быстро разлагаются;
2) мертвые организмы поедаются животными-падальщиками;
3) животные с мягким телом плохо поддаются фоссилизации;
4) лишь небольшая часть организмов погибала в условиях, благоприятных для фоссилизации;
5) обнаружена только часть ископаемых остатков.
Данные в пользу эволюционного процесса пополняются по мере обнаружения все большего числа возможных «недостающих звеньев» — либо ископаемых остатков, таких как сеймурия (амфибии → рептилии), археоптерикс (рептилии → птицы) и циногнатус (рептилии → млекопитающие), либо ныне живущих организмов как, например, перипатус (см. рис. 26.17) и латимерия, близкие по своему строению к вымершим формам.
Возможно также, что новые виды возникали внезапно и, что промежуточных форм не существовало. Эддредж и Гоулд (Eldredge, Gould) описали процесс, названный «прерывистым равновесием», позволяющий объяснить внезапное появление в палеонтологической летописи новых видов. По их мнению, виды остаются неизменными на протяжении длительного времени, а затем за сравнительно короткие периоды времени дают начало новым видам. Предполагается, что это происходит в результате относительно внезапного и существенного изменения условий среды. Следует также иметь в виду, что скорости эволюции способны варьировать, так что новые виды возникают быстро, создавая впечатление неполноты ископаемых данных. Такие кажущиеся «скачки» в эволюционной последовательности породили термин «скачкообразная эволюция». Сам Дарвин обсуждал такую возможность, написав в «Происхождении видов»:
«Я не думаю, что этот процесс [видообразование]... происходит непрерывно; гораздо более вероятно, что каждая форма долгое время остается неизменной, а затем вновь претерпевает модификацию».
Эволюция, следовательно, не всегда должна быть постепенной.
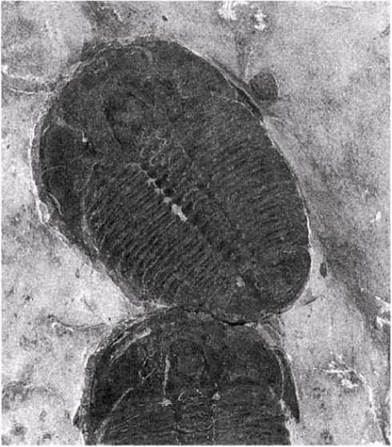
Рис. 26.3. Фотография ископаемых трилобитов из кембрийских отложений.
Эволюционная история лошади
Одним из лучших примеров филогении, основанной на изучении почти полного палеонтологического материала из североамериканских осадочных отложений от нижнего эоцена до настоящего времени, служит эволюция лошади.
Самые первые непарнокопытные появились 54 млн. лет назад; к современным представителям непарнокопытных относятся лошади, тапиры и носороги. Древнейшее животное, остатки которого обнаружены в палеонтологической летописи, принадлежали к роду Hyracotherium, широко распространенному в раннем эоцене по всей Северной Америке и Европе. К началу олигоцена оно вымерло везде, кроме Северной Америки. Это было небольшое субтильное животное, приспособленное к бегу. Его короткие тонкие ноги оканчивались вытянутыми стопами, а пальцы располагались почти вертикально. На передних ногах было по четыре, а на задних — по три пальца. Резцы были небольшие, а у коренных зубов была низкая коронка с округлыми бугорками, покрытыми эмалью.
В предполагаемый эволюционный ряд лошадей от Hyracotherium до Equus входит не менее 12 родов и несколько сотен видов. Главные направления в эволюции лошадей касались передвижения и питания. Они отражали адаптацию к изменявшимся условиям среды и включали следующие тенденции:
1) увеличение размеров тела;
2) удлинение ног и стоп;
3) редукцию боковых пальцев;
4) удлинение и утолщение третьего пальца;
5) выпрямление и увеличение жесткости спины;
6) совершенствование органов чувств;
7) увеличение и усложнение головного мозга, связанное с развитием органов чувств;
8) увеличение резцов в ширину;
9) замещение ложнокоренных зубов коренными;
10) удлинение зубов;
11) увеличение высоты коронки коренных зубов;
12) укрепление зубов благодаря развитию цемента между складками;
13) увеличение поверхности зубов в результате развития складок эмали.
На рис. 26.4 представлено прогрессивное развитие лошади в кайнозое; в качестве примеров выбраны роды, доминирующие в данной геологической эпохе. Однако сведениями, которые позволяли бы утверждать, что эти формы связаны прямым родством, мы не располагаем.
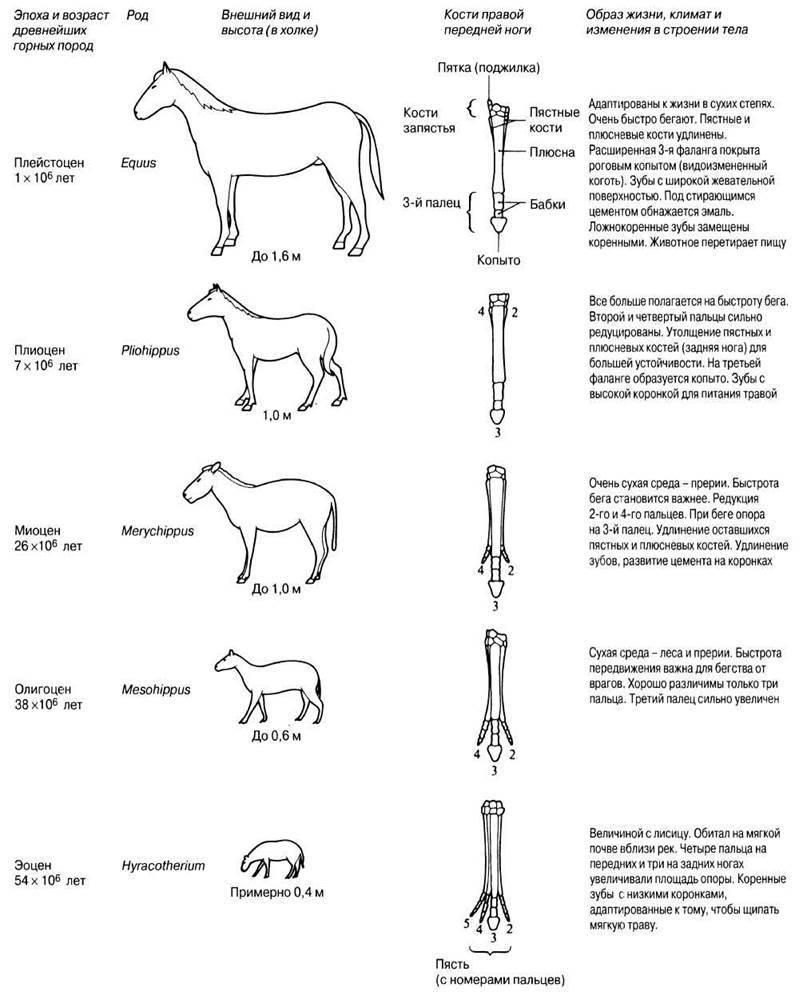
Рис. 26.4. Эволюция предков современной лошади.
Значение последовательного ряда форм, показанного на рис. 26.4, состоит в том, что он служит подтверждением теории прогрессивного развития гомологичных структур, в данном случае конечностей и зубов. Гомологичными структурами называют имеющиеся у разных видов сходные структуры, характеризующиеся общим эволюционным происхождением. Каждый из видов, представленных на рис. 26.4, соответствует некой стадии развития, которая процветала в течение нескольких миллионов лет (судя по обилию ископаемых остатков), прежде чем вымерла. Вымирание одного вида не означало, однако, исчезновения всего семейства. Как показывают ископаемые данные, место вымершего вида всегда занимал другой, близкородственный ему вид. Поскольку все виды, входящие в данную последовательность, обладают структурным и экологическим сходством, это подкрепляет теорию эволюции путем модификаций. Другие ископаемые остатки, обнаруженные в тех же слоях, свидетельствуют об изменявшихся климатических условиях; это вместе с другими данными позволяет считать, что каждый вид был адаптирован к преобладавшим в его время условиям.
В эволюционной линии лошади не наблюдается постепенного перехода, равномерно распределенного во времени и пространстве, да и палеонтологическую летопись нельзя считать полной. По-видимому, от линии, представленной на рис. 26.4, отходило несколько боковых ветвей, но все они вымерли. Современные лошади, очевидно, происходят от Pliohippus. Нынеживущий род Equusвозник в Северной Америке в плейстоцене и мигрировал в Евразию и Африку, где дал начало зебрам и ослам, а также современной лошади. Удивительно, что лошади, просуществовав в Северной Америке миллионы лет, вымерли там несколько тысяч лет назад, в период, совпавший с появлением там человека. Рисунки, сохранившиеся на стенах пещер в других частях земного шара, указывают на то, что древние люди, по-видимому, использовали лошадей в пищу. Вторично лошади были интродуцированы в Северную Америку почти 500 лет назад.