БИОЛОГИЯ Том 3 - руководство по общей биологии - 2004
26. ЭВОЛЮЦИЯ, ИЛИ ИСТОРИЯ ЖИЗНИ НА ЗЕМЛЕ
26.7. Данные в пользу теории эволюции
26.7.2. Географическое распространение
Все организмы в большей или меньшей степени адаптированы к своей среде обитания. Если абиотические и биотические факторы, имеющиеся в определенном местообитании, могут обеспечить существование какого-то вида, то можно было бы ожидать, что этот вид будет обнаружен в аналогичном местообитании и в другой сходной географической области, например львы, обитающие в африканских саваннах, могли бы жить и в южноамериканских пампасах. Однако на самом деле это не так. Распространение животных и растений на земном шаре носит прерывистый характер. Это нередко обусловлено экологическими факторами, однако данные об успешном заселении новых местообитаний животными и растениями, интродуцированными в них человеком, позволяют думать, что наряду с экологической адаптацией в этом участвуют и какие-то иные факторы. Кроликов в Австралии не было, но быстрое увеличение их численности после того, как они были завезены туда человеком, указывает на то, что австралийские местообитания им подходят. Сходными примерами служат факты распространения человеком домашних животных и культурных растений, таких как овцы, кукуруза, картофель и пшеница. Рациональное объяснение прерывистого распространения организмов основано на концепции, согласно которой виды возникают в какой-то данной области, а затем расселяются из нее. Масштабы расселения зависят от того, насколько успешно данный организм сможет обосноваться в новых местах, от эффективности механизма расселения и от наличия или отсутствия естественных преград, таких как океаны, горные хребты и пустыни. По-видимому, наиболее хорошо адаптированы к распространению через сушу и моря споры и семена, переносимые ветром, и летающие животные.
В противоположность этому, несмотря на то, что организмы, как правило, ограничены в своем распространении определенными частями земного шара, во многих случаях близкородственные формы обитают в сильно удаленных друг от друга областях; например, три современных вида двоякодышащих рыб встречаются порознь в тропических областях Южной Америки (Lepidosiren), в Африке (Protopterus) и в Австралии (Neoceratodus); верблюды и ламы живут в Северной Африке, Азии и Южной Америке; еноты широко распространены в Северной и Южной Америке, но обитают также в небольшом районе Юго-Восточной Азии. Судя по ископаемым данным, в прошлом эти животные были распространены гораздо шире, чем в наши дни.
Все эти сведения не имеют непосредственного отношения к эволюционной теории, однако они указывают на то, что распределение суши на земном шаре не всегда было таким, как в наше время.
Прежде считалось, что материки и океаны всегда занимали такие положения, как теперь. Первые геологи, такие как Хаттон (Hutton) и Лайель (Lyell); см. табл. 26.1), полагали, что осадочные породы образовались в результате периодических поднятий и опусканий уровня моря. Позднее возникло представление, что в прошлом на земном шаре существовали два больших континента — Лавразия в Северном полушарии и Гондвана в Южном; эти два массива суши соединялись обширными мостами, по которым могли мигрировать и расселяться растения и животные. Позднее результаты геологических исследований несколько изменили эту точку зрения, и в настоящее время общепринятой стала теория дрейфа континентов, основанная на тектонике литосферных плит. Гипотезу о дрейфе континентов впервые выдвинул Снайдер (Snider) в 1858 г., а затем в конце XIX в. ее развили Тейлор (Taylor) в США и Вегенер (Wegener) в Германии. Вегенер считал, что в карбоне Лавразия и Гондвана составляли единый массив суши, названный им Пангеей (от греч. pán — все; gaia — земля), который «плавал» на более плотном расплавленном внутреннем веществе Земли. В настоящее время, однако, принято считать, что континенты раздвинулись под действием глубинных конвективных течений, направленных вверх и в стороны и тянущих за собой плиты, на которых плавают континенты. Это позволяет объяснить непрерывное перемещение массивов суши и современное распространение таких животных, как, например, двоякодышащие рыбы (рис. 26.5).
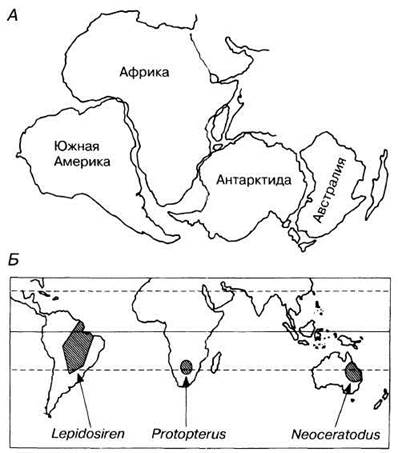
Рис. 26.5. А. Относительное расположение Южной Америки, Африки и Австралии на ранних стадиях дрейфа континентов, указывающее на близость областей, в которых, возможно, возникли двоякодышащие рыбы. Б. Современные ареалы двоякодышащих.
Что же касается верблюдов и лам (семейство Camelidae), то, как полагают, они происходят от общего предка, который, судя по ископаемым находкам, возник в Северной Америке. В плейстоцене этот предок распространился на юг, перейдя по Панамскому перешейку в Южную Америку, и на север, проникнув в Азию до того, как в результате подъема уровня моря она отделилась от Северной Америки (рис. 26.6). Считают, что на протяжении всего этого времени в группе Camelidae происходили прогрессивные изменения, приведшие к образованию двух родов — Camelus и Lama — в краевых областях их плейстоценовой миграции. Формы, промежуточные между современными верблюдами и ламами, были обнаружены в ископаемых остатках по всей Северной Америке. Как показывают палеонтологические данные, другие представители этого семейства в других частях земного шара вымерли в конце последнего оледенения.

Рис. 26.6. Карта современного распространения представителей семейства Camelidae: верблюдов в Азии и в Северной Африке, а лам — в Южной Америке. Судя по ископаемым находкам в плейстоцене Camelidaeбыли распространены по всей Северной и Южной Америкам, в значительной части Азии и в Северной Африке. Черными линиями со стрелками показаны возможные пути миграции. (По Matthewes, 1939, Climateof Evolution, vol. 1, 2nd ed., N.Y. Acad. of Sci.)
Еще одним примером прерывистого распространения, возникшего в результате географической изоляции, служат однопроходные и сумчатые Австралийской области. Австралия откололась от остальных массивов суши в конце юры сразу после появления примитивных млекопитающих. Млекопитающих делят на три подкласса: однопроходных (Monotremata), сумчатых (Marsupialia) и плацентарных (Eutheria). В Австралийской области развивались только две первые группы. Здесь они сосуществовали, претерпев адаптивную радиацию, в результате которой сложилась характерная австралийская фауна, представленная однопроходными Tachyglossus и Zaglossus (ехидны и проехидны) и Omithorhynchus (утконос) и 45 родами сумчатых. В других областях земного шара возникли более высокоразвитые плацентарные млекопитающие. Видимо, они по мере своего распространения по континентам вытесняли более примитивных однопроходных и сумчатых из их экологических ниш и остановились лишь там, где их дальнейшему расселению помешали географические преграды, как на пути в Австралию.
Все изложенное выше можно вкратце свести к следующему:
1) виды возникали в какой-то определенной области;
2) они расселялись за пределы этой области;
3) большинство видов могли расселяться только в том случае, если массивы суши располагались достаточно близко один от другого;
4) отсутствие в какой -либо области более высокоорганизованных форм обычно указывает на то, что данная область отделилась от родины этих форм до возникновения последних.
Ни один из приведенных выше фактов не позволяет объяснить механизм возникновения видов, однако все они указывают на то, что разные группы возникали в разное время и в разных областях. Палеонтологическая летопись дает представление о направлениях, в которых происходило постепенное изменение этих организмов, но она опять-таки не содержит никаких указаний на возможный механизм такого изменения.
О вероятном механизме возникновения видов в результате естественного отбора можно судить на основе распространения животных и растений на океанических островах. И Уоллеса, и Дарвина поразило огромное разнообразие видов, обитающих на таких островах, как Гавайские или Галапагосские. Как показывают геологические данные, эти острова образовались в результате вулканической активности, поднявшей их со дна океана, так что они никогда не имели прямой географической связи ни с одним массивом суши. Найденные на этих островах растения были, по всей вероятности, занесены на них ветром в виде спор и семян или по воде в виде плавучих семян или растительной массы. Водные и полуводные животные, как полагают, были принесены морскими течениями, а наземные организмы приплыли на бревнах или на плавучих скоплениях растений. Птицам, летучим мышам и летающим насекомым заселить эти острова было значительно проще.
Галапагосские острова находятся в Тихом океане, на экваторе, на расстоянии почти 1200 км к западу от Эквадора, и образуют архипелаг, описанный более подробно в разд. 27.8.3. В 1835 г., когда Дарвин впервые посетил эти острова, его внимание привлекло сходство обитающих на них видов с видами, населяющими ближайший континент; аналогичное сходство островных и материковых форм он обнаружил на островах Зеленого мыса, лежащих вблизи побережья Западной Африки. Однако на океанических островах растения и животные были в большинстве случаев заметно крупнее. Это можно объяснить отсутствием здесь крупных, более высокоорганизованных доминантных видов, с которыми более мелким родичам островных видов приходилось конкурировать на континенте. Например, слоновая черепаха (Geohelone elephantopus), которая питается имеющейся на островах обильной растительностью, достигла, вероятно, своих гигантских размеров — почти 2 м длины и 260 кг веса — благодаря отсутствию конкуренции со стороны различных млекопитающих, населяющих континент. Дарвин заметил также, что обитающие на Галапагосских островах многочисленные игуаны опять-таки были значительно крупнее своих континентальных родичей. Ящерицы обычно живут на суше, однако из обнаруженных на Галалагосских островах двух видов игуан один обитает в море. Водная галапагосская игуана (Amblyrhyncus cristatus) питается морскими водорослями и адаптирована к передвижению в воде благодаря уплощенному с боков хвосту и хорошо развитым перепонкам между пальцами на всех четырех конечностях (рис. 26.7). Предполагают, что конкуренция между наземными игуанами за пищу, жизненное пространство и брачного партнера создала давление отбора в пользу тех особей, у которых намечались изменения, благоприятные для жизни в водной среде. Этот механизм воздействия факторов среды на изменчивый генотип называют естественным отбором, и он уже был описан выше. Возможно, что именно в результате такого отбора постепенно сформировался морской вид игуаны. Однако не исключено, что самое сильное влияние на мысли Дарвина о механизме возникновения видов оказало разнообразие адаптивных признаков у 13 видов вьюрков, обитающих на островах. В Эквадоре, т, е, на континенте, обитали вьюрки только одного типа, клюв которых был приспособлен для дробления семян. У вьюрков Галапагосских островов обнаружены клювы шести основных типов, каждый из которых адаптирован к определенному способу питания. Различные группы вьюрков, особенности их питания и число видов в каждой группе представлены на рис. 26.8.

Рис. 26.7. Гигантская морская игуана Галапагосских островов (под водой).


Рис. 26.8. А. Адаптивная радиация дарвиновых вьюрков (по Lack). Б. Самец кактусового вьюрка (Geospiza scandens).
Дарвин предположил, что острова были колонизованы группой вьюрков с континента. Здесь они хорошо прижились, и неизбежная конкуренция, возникшая в результате роста их численности, а также наличие незанятых экологических ниш благоприятствовали освоению разнообразных ниш формами, имевшими соответствующие адаптивные вариации. Видовые отличия сводятся к небольшим различиям в размерах тела, окраске оперения и форме клюва. Несколько видов встречаются на всех крупных островах. Земляные и славковые вьюрки, которых считают самыми примитивными, живут на большей части островов. Насекомоядные древесные вьюрки и растительноядный древесный вьюрок отсутствуют на периферийных островках, а дятловые вьюрки обитают только на островах центральной группы. Современное распространение видов представляет большой интерес; согласно Лэку, его можно объяснить адаптивной радиацией и географической изоляцией. Например, на центральных островах обитает много видов, принадлежащих к разным группам (земляные, древесные, славковые и дятловые), а не к одной какой-то группе. Даже там, где обитают несколько видов одной группы (как на периферийных островах), все они различаются по своим экологическим требованиям. Это соответствует принципу конкурентного исключения Гаузе (см. разд. 10.7.5), согласно которому два или более близкородственных видов могут занимать одну и ту же область только в том случае, если их экологические требования различны.