БИОЛОГИЯ Том 3 - руководство по общей биологии - 2004
27. МЕХАНИЗМЫ ВИДООБРАЗОВАНИЯ
27.5. Естественный отбор
Гипотеза естественного отбора, постулированная Дарвином и Уоллесом, основывалась на исторических данных. Дарвин считал, что промежуток времени, необходимый для эволюционного изменения популяции, должен быть слишком большим, чтобы такое изменение можно было наблюдать непосредственно. Происходящие в последнее время изменения, связанные с промышленной, технической и медицинской революцией, создают столь сильные давления направленного и дизруптивного отбора, что теперь мы можем наблюдать резкие изменения в генотипе и фенотипе популяций, происходящие достаточно быстро. Открытие в сороковых годах антибиотиков создало сильное давление отбора в пользу бактериальных штаммов, обладающих генетической устойчивостью к антибиотикам. Бактерии очень быстро размножаются и дают ежедневно много поколений и миллионы особей. В результате случайной мутации может появиться устойчивая клетка, потомки которой будут процветать благодаря отсутствию конкуренции со стороны других бактерий, уничтожаемых данным антибиотиком. В ответ на это приходится создавать новые антибиотики для уничтожения устойчивых бактерий, и цикл продолжается. Селективное давление создается также в результате использования таких веществ, как ДДТ для борьбы с платяной вошью и комарами и антикоагулянт варфарин для уничтожения крыс. После возникновения устойчивости она быстро распространяется по всей популяции.
Классическим примером эволюционного изменения служит реакция некоторых бабочек на направленное селективное давление, создаваемое загрязнением атмосферы в результате промышленной революции. За последние 100 лет у более чем 80 видов бабочек появились темные формы, встречающиеся теперь с различной частотой по всей Великобритании. Это явление известно под названием индустриального меланизма. До 1848 г. все описанные формы березовой пяденицы (Biston betularia) имели бледную кремовую окраску с черными точками и отдельными темными пятнышками (рис. 27.9). В 1848 г. в Манчестере была обнаружена черная форма этой бабочки, а к 1895 г. популяция березовой пяденицы в Манчестере на 98% состояла из черных бабочек. Эта черная «меланистическая» форма появилась в результате повторных случайных мутаций, причем фенотип мутантных особей обладал большим селективным преимуществом в промышленных районах по причинам, выдвинутым и проверенным д-ром Кеттлуэллом.

Рис. 27.9. Полиморфизм у березовой пяденицы (Bistort betularia), А. Нормальная форма Biston betilaria typica. Б. Меланистичеасая форма Biston betularia carbonaria. (Из Е. В. Ford (19 73) Evolution studied by observation and experiment, Oxford Biology Readers, 55, Oxford University Press.)
Березовые пяденицы активны в ночное время, а днем они отдыхают на стволах деревьев. Нормальная форма имеет очень эффективную покровительственную окраску, которая сливается с лишайниками, покрывающими стволы. С развитием промышленной революции диоксид серы, образующийся при сжигании угля, вызвал в промышленных районах гибель лишайников, и в результате обнажилась темная кора деревьев, которая стала еще темнее из-за покрывающей ее сажи (рис. 27.10).

Рис. 27.10. Механистическая и немеланистическая формы Biston betularia на стволах деревьев вблизи Бирмингема (А) и в Дорсете (Б). (С любезного разрешения д-ра Н. В. Kettlewell, Department of Zoology, University of Oxford.)
В пятидесятых годах XX в. Кеттлуэлл пометил определенное число светлых и темных бабочек и выпустил их в двух местах: в загрязненном районе вблизи Бирмингема, где популяция на 90% состояла из темной формы, и в незагрязненном районе в Дорсете, где темная форма встречалась редко. С помощью световой ловушки он провел отлов меченых бабочек и получил следующие результаты:
Бирмингем |
Дорсет |
|
Меченые темные бабочки, % |
34,1 |
6,3 |
Меченые светлые бабочки, % |
15,9 |
12,5 |
С помощью киносъемки Кеттлуэлл показал, что малиновки и дрозды питаются этими бабочками, осуществляя форму естественного отбора, называемую избирательным выеданием, которая в данном случае оказывает селективное давление на распространение меланистической и светлой форм.
Полученные результаты, свидетельствуют о том, что меланистическая форма Biston betularia carbonaria в промышленных районах обладает селективным преимуществом перед светлой формой Biston betularia typica, тогда как светлая форма обладает преимуществом в незагрязненных районах.
Позднее было установлено, что окраска темной формы определяется доминантным меланистическим аллелем. На рис. 27.11 показано распространение этих двух форм на Британских островах в 1958 г.
Наличие темных бабочек в непромышленных районах восточной части Англии объясняется распространением меланистических форм и преобладающими там западными ветрами. После того как в 1956 г. был введен в действие Акт о чистоте воздушного бассейна, доля светлых форм вновь стала значительно выше, так как в промышленных районах элиминация их отбором снизилась.
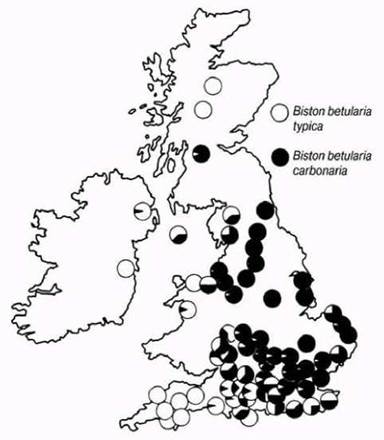
Рис. 27.11. Распространение меланистической и немеланистической форм Biston betularia на Британских островах в 1958 г. (Н. В. D. Kettleweli, Heredity, 1978, 12, 51-72).
27.5.1. Полиморфизм
Полиморфизм играет значительную роль в процессе естественного отбора. Его можно определить как существование в пределах одной популяции двух или нескольких форм данного вида, различающихся по биохимическим, морфологическим или поведенческим признакам. Различают две формы полиморфизма: переходный полиморфизм и сбалансированный, или стабильный, полиморфизм.
Сбалансированный полиморфизм
Сбалансированный полиморфизм создается при сосуществовании в одной популяции различных форм при стабильных условиях среды. Наиболее ярким его примером служит наличие двух полов у животных и растений. Частоты генотипов различных форм сбалансированы, так как обе формы обладают равноценными селективными преимуществами. Примером сбалансированного полиморфизма у человека служат группы крови А, В, АВ и О. Частоты разных генотипов в разных популяциях могут варьировать, однако в данной популяции они остаются постоянными из поколения в поколение. Это объясняется тем, что ни один генотип не обладает селективным преимуществом перед другими. Как показывают статистические данные, у мужчин белой расы с группой крови О ожидаемая продолжительность жизни выше, чем у мужчин с другими группами крови, однако у обладателей группы О чаще, чем у других, развивается язва двенадцатиперстной кишки, которая в случае прободения может привести к смерти. Другие примеры полиморфизма — нормальное зрение и цветовая слепота у человека, касты рабочих, трутней и маток у общественных насекомых и длинно- и короткостолбчатые формы у первоцвета.
Классическое количественное изучение сбалансированного полиморфизма провели Кейн, Карри и Шеферд (Cain, Currey, Shepherd) на обыкновенной наземной улитке Cepaea nemoralis. У этих улиток раковина бывает желтая (если внутри находится живая улитка, то раковина выглядит зеленой), различных оттенков коричневого, в том числе светло-палевая, розовая, оранжевая, и красная. Устье раковины может быть темно-коричневым, розовым или белым, а на самой раковине может быть до пяти темных полос, тянущихся по ее контурам (рис. 27.12). Как цвет раковины, так и характер ее полос детерминированы генетически. Окраска определяется множественными аллелями, причем коричневая окраска доминирует над розовой, и обе они доминируют над желтой. Полосатость — рецессивный признак.

Рис. 27.12. Изменчивость окраски и рисунка полос у раковины Сeраеа nemoralis. А — вид сверху: Б — вид сбоку. Показан постепенный переход от желтой раковины без полос (верхний ряд слева) до темно-коричневой полосатой (нижний ряд справа). (По Tribe. Tallan, Erant (1978), Basic Biology Course, Book 12, Cambridge University Press.)
Улиток поедают дрозды: они переносят улиток на близлежащий камень и, используя его в качестве наковальни, разбивают раковину, чтобы добраться до улитки. Изучая соотношение раковин разных типов, разбросанных вокруг наковален и в местообитаниях улиток в целом, Кейн, Карри и Шеферд показали, что в популяции действуют селективные факторы. В тех местах, где фон был довольно однородный, например на траве или на лесной подстилке, селективным преимуществом обладали желтые и коричневые раковины без полос — вокруг наковален таких раковин было меньше (рис. 27.13). Более темные полосатые раковины имели преимущество там, где фон был пестрым, как, например, на пастбищах с грубыми травами и в живых изгородях. На любых участках дрозды истребляют больше всего хорошо заметные формы. Обширная популяция полиморфных улиток может занимать несколько участков, различающихся по фону. Кроме того, характер и окраска фона могут изменяться со сменой времен года. Хотя выедание хорошо заметных форм происходит непрерывно, ни одна форма не обладает абсолютным селективным преимуществом; поэтому доля каждой формы в популяции из года в год остается довольно постоянной.

Рис. 27.13. Раковины Сераеа nemoralss без полос на фоне листовой подстилки. Раковина, лежащая справа — желтая, вверху — розовая, а две раковины слева — коричневые. (По Е. В. Ford, Evolution studied by observation and experiment. Oxford Biology Reader, 55, Oxford University Press. 1973.)
Баланс численностей разных форм не обязательно определяется только окраской и характером полосатости. Судя по некоторым данным, сохранению равновесия при полиморфизме способствуют также физиологические факторы. 13 некоторых областях с сухой известковой почвой и светлым фоном не всегда доминируют формы с наименее заметными окраской и характером полос. Полагают, что у Сераеа полиморфизм определяется сцеплением генов особого типа: гены окраски и полосатости сцеплены и образуют суперген, действующий и наследуемый как одна генетическая единица. Входящие в него гены определяют признаки, обладающие такими селективными преимуществами, благодаря которым они сохраняются в популяции. Именно разнообразие аллельных форм этих генов, охраняемое вследствие широкого распространения гетерозигот, составляет основу полиморфизма. К этому добавляется сцепление генов, детерминирующих некоторые физиологические функции, что, как полагают, тоже способствует поддержанию сбалансированного полиморфизма. Наличие в одной и той же популяции нескольких обособленных форм, доля которых слишком велика, чтобы их можно было отнести за счет повторных мутаций, называют генетическим полиморфизмом; примером служит Сераеа.
Переходный полиморфизм
Переходный полиморфизм возникает в тех случаях, когда различные формы, или морфы, существуют в популяции, испытывающей сильное давление отбора. Частота фенотипического проявления каждой формы определяется интенсивностью давления отбора, как в случае меланистичеcкой и светлой форм березовой пяденицы. Переходный полиморфизм обычно наблюдается при постепенном замещении одной формы другой.