БИОЛОГИЯ Том 3 - руководство по общей биологии - 2004
23. НЕПРЕРЫВНОСТЬ ЖИЗНИ
23.9. Регуляция генной активности
После того как был расшифрован генетический код и была установлена структура ДНК, исследования в области генетики ушли далеко вперед. Одна из главных проблем, которая особенно волнует молекулярных генетиков, это вопрос о том, каким образом регуляция активности генов обеспечивает выполнение каждой клеткой единой программы развития и ее собственных функций.
Все соматические клетки данного организма несут один и тот же набор генов, т. е. содержат одинаковое число хромосом, несущих одни и те же аллели. И тем не менее клетки многоклеточного организма очень разнообразны по структуре и функциям. Даже в одной и той же клетке скорость синтеза белковых молекул может быть различной в зависимости от обстоятельств и потребностей. Данные о механизмах, регулирующих активность генов в клетке, были впервые получены при изучении регуляции синтеза ферментов у Е. coli.
В 1961 г. Жакоб и Моно провели ряд экспериментов, желая понять природу индукции синтеза ферментов у Е. coli. Полагают, что в клетках этой бактерии синтезируется около 800 ферментов. Синтез некоторых из них происходит непрерывно, и их называют конститутивными ферментами; другие же образуются только в присутствии надлежащего индуктора, который может и не быть субстратом данного фермента. Такие ферменты, примером которых служит β-галактозидаза, называют индуцибельными ферментами.
Е. coli быстро растет на культуральной среде, содержащей глюкозу. При переносе клеток на среду, содержащую вместо глюкозы лактозу, рост начинается не сразу, а после короткой задержки, но затем продолжается с такой же скоростью, как и на среде с глюкозой. Проведенные исследования показали, что для роста на среде с лактозой необходимо наличие двух веществ, которые Е. coli обычно не синтезирует: β-галактозидазы, гидролизующей лактозу до глюкозы и галактозы, и лактозопермеазы, делающей клетку способной быстро поглощать лактозу из среды. Это служит примером того, как изменение в условиях среды (замена глюкозы лактозой) индуцирует синтез определенного фермента. Другие эксперименты с Е. coli показали, что высокое содержание в среде аминокислоты триптофана подавляет продуцирование триптофансинтазы — фермента, необходимого для синтеза триптофана. Синтез β-галактозидазы служит примером индукции фермента, а подавление синтеза триптофансинтазы — примером репрессии фермента. На основе этих наблюдений Жакоб и Моно предложили механизм, объясняющий индукцию и репрессию — механизм «включения» и «выключения» генов.
23.9.1. Гипотеза Жакоба-Моно
Генетические инструкции, определяющие аминокислотную последовательность упоминавшихся выше белков, заключены в структурных генах. Инструкции для β-галактозидазы и лактозопермеазы тесно сцеплены в одной хромосоме. Активность этих генов регулируется еще одним геном — геном-регулятором, который, как полагают, препятствует переходу структурных генов в активное состояние. Ген-регулятор может находиться на некотором расстоянии от структурных генов. Доказательства его существования получены при изучении мутантных клеток Е. coli, лишенных этого гена и поэтому вырабатывающих β-галактозидазу непрерывно. Ген-регулятор содержит генетическую информацию для синтеза репрессора, который препятствует активности структурных генов. Репрессор действует на структурные гены не прямо, а опосредованно, оказывая влияние на участок, примыкающий к структурным генам и называемый геном-оператором. Оператор и структурные гены в совокупности называют опероном (рис. 23.34).
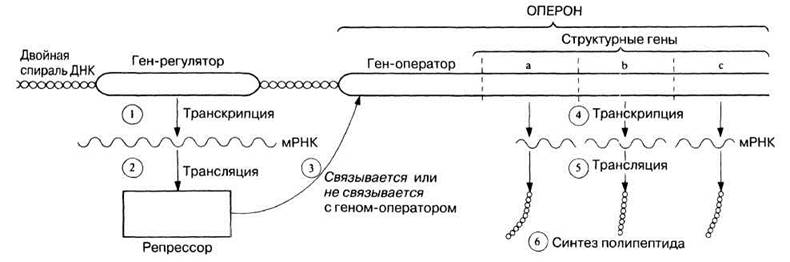
Рис. 23.34. Основные структуры и процессы, участвующие в регуляции белкового синтеза согласно гипотезе Жакоба и Моно. Цифрами указана последовательность событий.
Репрессор представляет собой особый аллостерический белок, который либо связывается с геном-оператором, подавляя его активность («выключает» его), либо не связывается с ним, позволяя ему стать активным («включает» его). Когда оператор «включен», структурные гены осуществляют транскрипцию и образуется мРНК, которую рибосомы и тРНК транслируют в полипептиды. Когда оператор «выключен», мРНК не образуется и кодируемые ею полипептиды не синтезируются (рис. 23.34).
Механизм, от которого зависит, присоединится ли аллостерический белок к гену-оператору или нет, прост, но при этом чувствителен к изменениям внутри клетки. Предполагается, что в молекуле репрессора имеется по меньшей мере два активных участка, к которым может присоединиться либо молекула индуктора, либо молекула репрессора, в зависимости от их концентрации в данное время, как это описано в разд. 23.9.4.
23.9.2. Индукция ферментов
Присоединение молекулы индуктора к ее активному участку на молекуле репрессора изменяет третичную структуру репрессора (аллостерический эффект; см. разд. 4.4.4), так что он не может связаться с геном-оператором и репрессировать его. Ген-оператор становится активным и «включает» структурные гены.
При культивировании Е. coli на среде с глюкозой ген-регулятор продуцирует белок, обладающий свойствами репрессора, который связывается с геном-оператором и «выключает» его. Структурные гены при этом не активируются, и ни β-галактозидаза, ни лактопермеаза не синтезируются. При переносе бактерий на среду с лактозой последняя, как полагают, действует как индуктор синтеза белка, присоединяясь к молекуле репрессора и препятствуя ее соединению с геном-оператором. Структурные гены переходят в активное состояние, продуцируется мРНК и синтезируются белки. Таким образом лактоза индуцирует собственное расщепление.
23.9.3. Репрессия ферментов
Если молекула какого-либо корепрессора связывается своим активным участком с молекулой репрессора, то она усиливает эффект от связывания репрессора с геном-оператором. При этом происходит инактивация гена-оператора, что фактически препятствует «включению» структурных генов.
Е. coli синтезирует аминокислоту триптофан в присутствии фермента триптофансинтазы. При избыточном содержании в клетке триптофана некоторая его часть действует как корепрессор синтеза фермента, связываясь с молекулой репрессора. Молекулы корепрессора и репрессора присоединяются к гену-оператору и подавляют его активность. Структурные гены «выключаются», мРНК не образуется и синтез триптофансинтазы прекращается. Это пример ингибирования по принципу обратной связи на генном уровне.
23.9.4. Регуляция метаболических путей
Описанный выше двойной механизм индукции и репрессии делает возможным взаимодействие между цитоплазмой и ядром для обеспечения тонкой регуляции метаболизма клетки. В случае какого-либо простого метаболического пути исходный субстрат и конечный продукт могут играть роль соответственно индуктора и репрессора. Благодаря этому клетка может синтезировать фермент в таком количестве, которое необходимо в данное время для поддержания количества конечного продукта на нужном уровне. Такой способ регуляции метаболизма чрезвычайно экономичен. Отрицательная обратная связь, осуществляемая путем инактивации первого фермента в результате его связывания с конечным продуктом, быстро блокирует данный метаболический путь, но не препятствует синтезу других ферментов. В модели, предложенной Жакобом и Моно, конечный продукт, присоединяясь к молекуле репрессора, чтобы усилить его ингибирующее действие на ген-оператор, подавляет синтез всех ферментов и выключает данный метаболический путь.
23.9.5. Модификация гипотезы оперона
После того, как в 1961 г. Жакоб и Моно предложили механизм, с помощью которого гены могут включаться и выключаться, были получены другие данные, позволившие прояснить различные аспекты этого механизма. На основе генетических данных было высказано предположение о существовании гена-промотора, расположенного рядом с геном-оператором и действующего между ним и геном-регулятором. Предполагается, что ген-промотор несет две функции. Во-первых, он служит местом, к которому присоединяется РНК-полимераза, прежде чем начать перемещаться вдоль ДНК в процессе транскрипции, т. е. в процессе синтеза мРНК на структурных генах. Это перемещение, конечно, зависит от того, находится ли ген-оператор в активном состоянии или нет. Во-вторых, последовательность оснований гена-промотора определяет, какая из цепей двойной спирали ДНК присоединяет к себе РНК-полимеразу, т. е. служит матрицей для транскрипции мРНК.