СОВРЕМЕННАЯ БОТАНИКА - П. РЕЙВН - 1990
РАЗДЕЛ I. РАСТИТЕЛЬНАЯ КЛЕТКА
ГЛАВА 4. ДВИЖЕНИЕ ВЕЩЕСТВ В КЛЕТКИ И ИЗ КЛЕТОК
Структура клеточных мембран
В настоящее время наибольшим признанием пользуется жидкостно-мозаичная модель мембраны, согласно которой мембрана состоит из бислоя липидов, в который погружены глобулярные белки (рис. 4-7). Эти белки, называемые интегральными, часто выступают из бислоя с одной или обеих сторон мембраны. Участок белковой молекулы, погруженный в бислой, является гидрофобным, участок вне его — гидрофильным. Возможно, но не доказано, что некоторые интегральные белки пронизаны гидрофильными каналами. Такую структуру имеют все мембраны клетки, в том числе плазматическая и внутренние мембраны.
Рис. 4-7. Жидкостно-мозаичная модель мембраны. Мембрана состоит из бислоя (двойного слоя) липидных молекул с гидрофобными хвостами, обращенными внутрь, и крупных белковых молекул. Белки, погруженные в бислой, называют интегральными. С внутренней поверхности мембраны к некоторым интегральным белкам прикрепляются так называемые периферические белки. Часть белковой молекулы, погруженная в липидный бислой, является гидрофобной, часть, выступающая из него, — гидрофильной. Короткие углеводные цепи присоединены к белкам внешней стороны плазматической мембраны. В целом структура жидкая, и считается, что белки плавают в липидном «море»
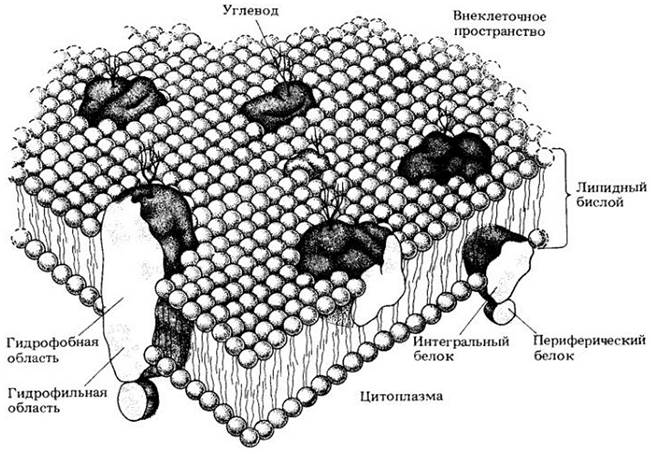
Две поверхности мембраны различаются по химическому составу. Например, два основных типа липидов плазматической мембраны растительных клеток — фосфолипиды (более распространенные) и стеролы — в двух половинах бислоя находятся в разных концентрациях. Более того, интегральные белки ориентированы внутри бислоя определенным образом и участки их молекул, выступающие за пределы бислоя в разные стороны, различаются по аминокислотному составу и третичной структуре. На внутренней поверхности мембраны к интегральным белкам прикрепляются дополнительные, так называемые периферические белки, на внешней — короткоцепочечные углеводы. Считается, что углеводы на внешней поверхности мембраны некоторых эукариотических клеток играют важную роль во взаимодействии между клетками и в модификации их поверхности.
В то время как липидный бислой определяет основную структуру клеточных мембран, за большую часть мембранных функций ответственны белки. Большинство мембран состоит на 40 — 50% (по массе) из липидов и на 50 — 60% из белков, причем количество и тип мембранных белков отражают ее функцию. Некоторые являются ферментами, которые регулируют связанные с мембранами реакции, другие — переносчиками, участвующими в транспорте специфических молекул внутрь клетки или из нее, третьи служат рецепторами для получения и преобразования химических сигналов из окружающей среды. Несмотря на то что некоторые интегральные белки, по-видимому, «заякорены» и остаются на одном месте, липидный бислой весьма подвижен. Некоторые белки более или менее свободно плавают в нем и так же, как молекулы липидов, могут латерально перемещаться, образуя различные ансамбли, или мозаики, структура которых постоянно меняется.
Транспорт через мембраны
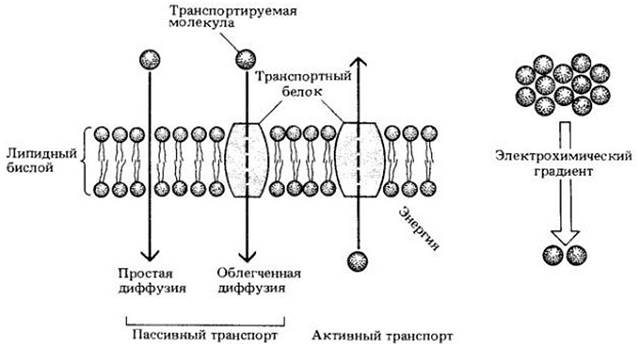
Молекулы проходят через мембраны благодаря трем различным процессам: простой диффузии, облегченной диффузии и активному транспорту. Неполярные (гидрофобные) вещества, в том числе О2, растворимые в липидах, обычно проникают через мембрану путем простой диффузии. (Наблюдение, что гидрофобные молекулы легко диффундируют через плазматические мембраны, послужило первым доказательством липидной природы мембраны.)
Воду и другие полярные (гидрофильные) молекулы липидный бислой мембраны, казалось бы, пропускать не должен, однако гидрофильные молекулы и ионы через него проходят. Как же это осуществляется? В случае воды и некоторых других полярных молекул, таких, как СО2, диффузия через мембрану возможна отчасти потому, что эти молекулы малы и незаряжены.
Диффузия неполярных или мелких незаряженных молекул через мембрану представляет собой пример пассивного транспорта. Его направление определяется только разностью концентраций вещества по обеим сторонам мембраны (градиентом концентрации). Однако если молекула заряжена, то на ее транспорт влияет как градиент концентрации, так и общий электрический градиент поперек мембраны (мембранный потенциал). Оба градиента вместе составляют электрохимический градиент. В растительных клетках обычно существуют электрические градиенты через плазматическую мембрану и тонопласт. Основное вещество заряжено отрицательно как по отношению к водной среде, окружающей клетки, так и по отношению к содержимому (клеточному соку) вакуоли.
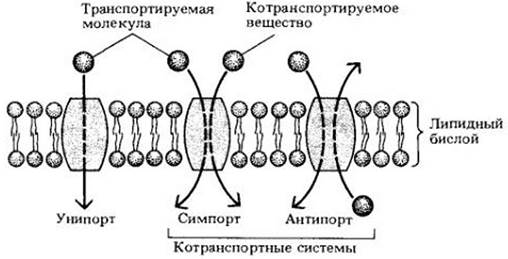
Большинство веществ, необходимых клеткам, полярно и переносится через мембрану с помощью погруженных в нее транспортных белков (белков-переносчиков). Некоторые транспортные белки переносят одно растворенное вещество через мембрану (унипорт). Другие функционируют как котранспортные системы, в которых перенос одного растворенного вещества зависит от одновременного или последовательного переноса второго вещества. Второе вещество может транспортироваться в том же направлении (симпорт) либо в противоположном (антипорт) (рис. 4-8). Все транспортные белки, по-видимому, образуют непрерывный белковый проход через мембрану, поэтому вещества, которые они переносят, непосредственно не контактируют с гидрофобной внутренней частью липидного бислоя.
Различают две основные формы транспорта с помощью переносчиков: облегченную диффузию и активный транспорт. Облегченная диффузия обусловлена градиентом концентрации. и молекулы движутся соответственно этому градиенту. Ни простая, ни облегченная диффузия (две разновидности пассивного транспорта) не способны идти против градиента концентрации (рис. 4-9) или электрохимического градиента. Перенос растворенных веществ против градиента концентрации или электрохимического градиента требует энергии и называется активным транспортом (рис. 4-10). В клетках растений и грибов активный транспорт осуществляется с помощью протонного насоса, использующего энергию АТР, который расщепляется Н+-АТРазой, локализованной в мембране. Фермент генерирует большой электрический потенциал и градиент pH, что создает движущую силу для поглощения веществ, котранспортируемых с Н+.
Рис. 4-8. Схема функционирования транспортных белков. Унипорт — простой перенос какого-либо растворенного вещества через мембрану. В котранспортных системах перенос одного вещества зависит от одновременного или последовательного переноса другого вещества либо в том же направлении (симпорт), либо в противоположном (антипорт)
Транспорт, в котором участвуют переносчики, будь то облегченная диффузия или активный транспорт, высокоизбирателен. Транспортный белок может взаимодействовать с какой-нибудь одной молекулой и не воспринимать другую, почти идентичную первой. Транспортный белок не претерпевает изменений в процессе транспорта. В этом отношении транспортные белки похожи на ферменты. Чтобы подчеркнуть эту особенность, они были названы пермеазами. Однако в отличие от ферментов транспортные белки обычно не вызывают химических изменений веществ, с которыми они временно связываются.
Рис. 4-9. Схема пассивного транспорта по электрохимическому градиенту и активного транспорта против электрохимического градиента. Простая диффузия и облегченная диффузия— пассивные процессы, в то время как для активного транспорта требуется энергия
Натриево-калиевый насос
Одной из важнейших и наиболее изученных систем активного транспорта в клетках животных является натриевокалиевый насос. (Насосами обычно называют механизмы, осуществляющие активный транспорт.) Большинство клеток животных поддерживает разные градиенты концентрации ионов натрия (Na+) и калия (К+) поперек плазматической мембраны: внутри клетки сохраняется низкая концентрация Na+ и высокая концентрация К+. Градиент Na+ используется клетками животных (как и фадиент Н+ клетками растений и грибов) для поглощения котранспортируемых с Na+ веществ. Энергия, необходимая для работы натриевокалиевого насоса, поставляется молекулами АТР, образующимися при дыхании. О значении этой системы для всего организма свидетельствует тот факт, что у находящегося в покое животного более трети АТР затрачивается на обеспечение работы натриево-калиевого насоса.
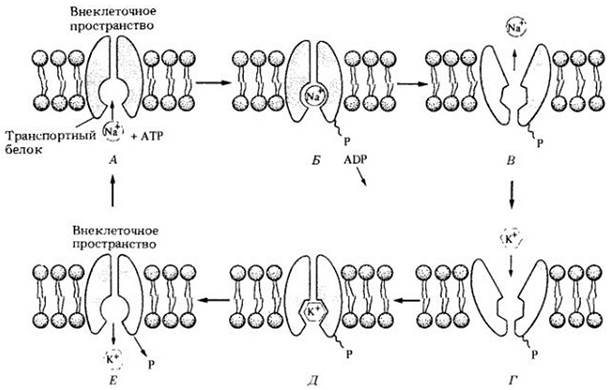
Транспорт Na+ и К+ осуществляется специальным белком, который, но мнению некоторых исследователей, существует в двух альтернативных формах. Одна из них имеет полость, открытую внутрь клетки, в которую может войти ион Na+. Другая имеет полость, открытую наружу, в нее входит ион К+. Как показано на рис. 4-10, вначале ион Na+ связывается с транспортным белком. Затем АТР расщепляется с образованием ADP, а освободившийся фосфат присоединяется к белку (белок фосфорилируется). Это вызывает изменение конформации (формы) белка, и Na+ переносится на внешнюю сторону мембраны и высвобождается. Затем транспортный белок захватывает ион К+, что ведет к дефосфорилированию белка, возвращению его к первоначальной конформации и высвобождению иона К+ внутри клетки. Этот процесс генерирует градиент Na+ и К+ поперек мембраны.
Рис. 4-10. Модель работы натриевокалиевою насоса. А. Ион Na+ в цитоплазме соединяется с молекулой транспортною белка. Б. Реакция с участием АТР, в результате которой фосфатная группа (Р) присоединяется к белку, а ADP (аденозиндифосфат) высвобождается. В. Фосфорилирование индуцирует изменение конформации белка, что приводит к высвобождению Na* за пределами клетки. Г. Ион К+ во внеклеточном пространстве связывается с транспортным белком (Д), который в этой форме более приспособлен для соединения с К+, чем с Na+. Е. Фосфатная группа отщепляется от белка, вызывая восстановление первоначальной формы, а ион К+ высвобождается в цитоплазму. Транспортный белок теперь готов к выносу другого иона Na+ из клетки
Протонный насос растений и грибов сходен с натриевым насосом животных. Растительный белок фосфорилируется в цепи реакций, похожей на описанную выше. Несмотря на сходство в химической структуре и механизме работы фермента, эти два насоса используют разные катионы: Н+ — у растений и грибов. Na+ — у животных.