СОВРЕМЕННАЯ БОТАНИКА - П. РЕЙВН - 1990
РАЗДЕЛ II. ЭНЕРГИЯ И ЖИВЫЕ КЛЕТКИ
ГЛАВА 7. ФОТОСИНТЕЗ
Темновые реакции
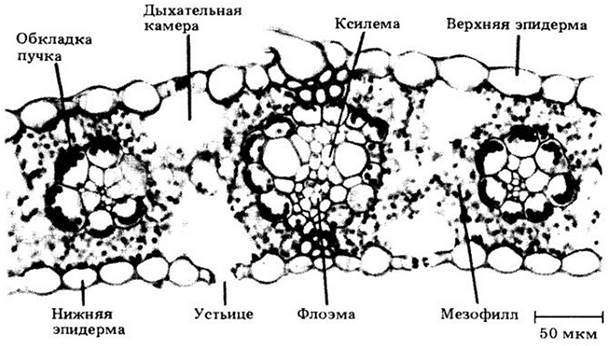
На второй стадии фотосинтеза химическая энергия, запасенная в световых реакциях, используется для восстановления углерода. Углерод, доступный для фотосинтезирующих клеток, поставляется в виде двуокиси углерода. Обнаружено; что водоросли и цианобактерии усваивают двуокись углерода, растворенную в воде. У большинства растений СО2 поступает к фотосинтезирующим клеткам через специальные отверстия, называемые устьицами, которые находятся в листьях и зеленых стеблях (рис. 7-14).
Рис. 7-14. Микрофотография (сканирующий электронный микроскоп) устьиц на нижней поверхности листа тополя (Populus). Углекислота поступает к фотосинтезирующим клеткам через устьица
Цикл Кальвина: С3-путь
Восстановление углерода происходит в строме хлоропласта в цикле реакций, которые известны как цикл Кальвина (названного в честь Мелвина Кальвина, который получил Нобелевскую премию за его открытие). Цикл Кальвина аналогичен циклу Кребса, поскольку в конце цикла происходит регенерация исходного соединения.
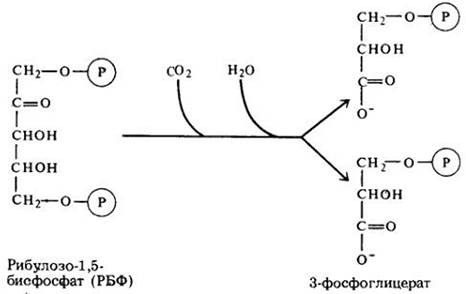
Исходное (или конечное) соединение цикла Кальвина — пятиуглеродный сахар с двумя фосфатными группами — рибулозо-1,5-бисфосфат (РБФ). Процесс начинается, когда двуокись углерода входит в цикл и фиксируется на РБФ. Образуемое при этом соединение затем расщепляется на две молекулы 3-фосфоглицерата (рис. 7-15). (Каждая молекула 3-фосфоглицсрата состоит из трех атомов углерода, отсюда другое название цикла Кальвина — С3-путь.)
Рис. 7-15. Кальвин и его сотрудники проводили кратковременное освещение фотосинтезирующих водорослей в атмосфере радиоактивного углекислого газа (14СО2). Они обнаружили, что радиоактивный углерод сначала включается в рибулозо-1,5-бисфосфат, который после этого немедленно расщепляется на две молекулы 3-фосфоглицерата. Радиоактивный углевод (он обозначен другим цветом) обнаруживается затем в одной из двух образовавшихся молекул. С данной реакции и начинается цикл Кальвина
Рибулозобисфосфаткарбоксилаза — фермент, катализирующий эти ключевые реакции, — самый распространенный в хлоропласте; его содержание составляет 15% от общего количества белка хлоропласта. (Говорят, что это самый распространенный белок в мире, можете вы сказать почему?) Данный фермент локализован на поверхности тилакоидных мембран.
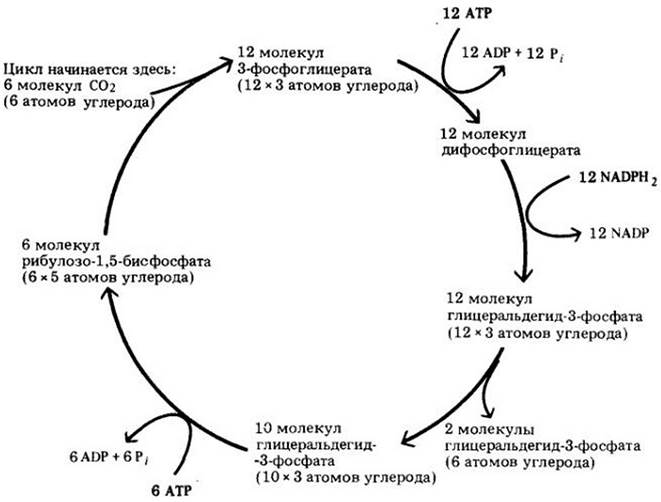
Полный цикл представлен на рис. 7-16. Как и в цикле Кребса, отдельная реакция катализируется специфическим ферментом. В течение каждого оборота цикла одна молекула СО2восстанавливается, а молекула РБФ — регенерируется. Шесть оборотов цикла с поглощением шести атомов углерода необходимо совершить, чтобы образовался шестиуглеродный сахар — глюкоза. Суммарное уравнение синтеза глюкозы можно записать следующим образом:
6СО2 + 12NADPH2 + 18ATP —> 1Глюкоза + 12NADP + 18ADP + 18Рi + 6Н2О.
Промежуточный продукт цикла — глицеральдегид-3-фосфат. Это же соединение образуется в процессе гликолиза при расщеплении фруктозо-1,6-бисфосфата. В аналогичных, но обратных реакциях идет синтез глюкозы из глицеральдегид- 3-фосфата с использованием энергии фосфатных связей.
Рис. 7-16. Общий итог цикла Кальвина. При каждом обороте в цикл входит одна молекула углекислоты. Здесь показан результат шести оборотов (для синтеза двух молекул глицеральдегид-3- фосфата необходимо шесть молекул СО2). Шесть молекул рибулозо-1,5-бис- фосфата, пятиуглеродного соединения, реагируют с шестью молекулами СО2, образуя таким образом двенадцать молекул 3-фосфоглицерата, трехуглеродного соединения, которые затем превращаются в двенадцать молекул глицеральдегид-3-фосфата. Десять из них перераспределяются с образованием шести пятиуглеродных молекул рибулозо-1,5-бисфосфата. Оставшиеся две молекулы глицеральдегид-3-фосфата представляют собой «чистый доход» цикла Кальвина. Энергия, обеспечивающая цикл Кальвина, поставляется в форме АТР и NADPH2, образовавшихся в световых реакциях
С4-нуть фотосинтеза
Цикл Кальвина — не единственный путь фиксации углерода в темновых реакциях. У некоторых растений первый продукт фиксации — не трехуглеродная молекула 3-фосфоглицерата, как в цикле Кальвина, а четырехуглеродное соединение — оксалоацетат (который образуется и в цикле Кребса). Растения, которые осуществляют этот путь, называются С4- растениями в отличие от С3-растений, где функционирует только цикл Кальвина. (С4-путь называют также циклом Хэтча — Слэка в честь австралийских физиологов растений, которые сыграли решающую роль в его открытии.)
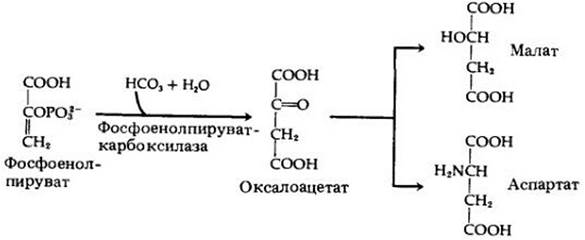
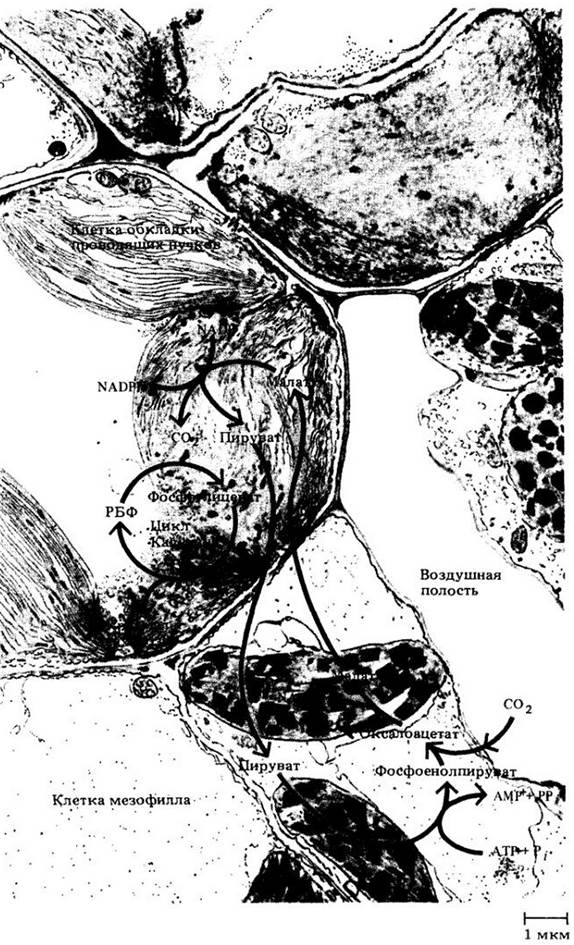
Оксалоацетат образуется, когда СО2 фиксируется на фосфоенолпирувате. Эта реакция катализируется ферментом фосфоенолпируваткарбоксилазой (рис. 7-17). Оксалоацетат затем восстанавливается до малата или превращается с добавлением аминогруппы в аспартат. Эти реакции происходят в клетках мезофилла. Следующий этап поразителен: малат (или аспартат — в зависимости от вида растения) перемещается из клеток мезофилла в клетки обкладок сосудистых пучков листа, где декарбоксилируется с образованием СО2 и пирувата. СО2 затем вступает в цикл Кальвина, реагирует с РБФ с образованием фосфоглицерата и других промежуточных соединений цикла, в то время как пируват возвращается в клетки мезофилла, где реагирует с АТР, образуя фосфоенолпируват (рис. 7-18). Таким образом, анатомия растения способствует пространственному разделению С4- пути и цикла Кальвина в листьях С4-растений.
Рис. 7-17. Фиксация углекислоты по С4-пути. Углекислота взаимодействует с фосфоенолпируватом при участии фермента фосфоенолпируваткарбоксилазы. Образующийся в этой реакции оксалоацетат либо восстанавливается до малата, либо превращается в аспартат в реакции аминирования (добавления NН2-группы). Эти этапы будут рассмотрены дальше вместе с процессом выделения СО2, вступающего в цикл Кальвина
Два первичных карбоксилирующих фермента используют различные формы СО2 в качестве субстрата. Рибулозо- бисфосфаткарбоксилаза использует СО2, который присутствует в клетке в концентрации около 15 — 20 мкМ в равновесии с воздухом. В то же время фосфоенолпируваткарбокси- лаза использует гидратированную форму двуокиси углерода — НСО-3. При pH 8,0 НСО3 обнаруживается в клетке в концентрации 15 — 20 мкМ в равновесии с воздушной фазой. Рибулозобисфосфаткарбоксилаза найдена в хлоропластах, в то время как фосфоенолпируваткарбоксилаза находится в основном веществе цитоплазмы.
Рис. 7-18. Путь фиксации углерода у С4-растений. СО2 на начальном этапе фиксируется в клетках мезофилла с образованием оксалоацетата, который быстро превращается в малат. Затем малат транспортируется в клетки обкладки, где выделяется СО2. Выделяемый СО2 утилизируется в цикле Кальвина; при этом образуются крахмал и сахароза. Пируват возвращается в клетку мезофилла для регенерации фосфоенолпирувата. Здесь показан лист одного из С4-растений, а именно кукурузы (Zea mays)
Обычно листья С4-растений характеризуются особым расположением клеток мезофилла вокруг слоя больших клеток обкладок, поэтому вместе они образуют два концентрических слоя вокруг сосудистого пучка (рис. 7-19). Эта кольцеобразная организация клеток получила название кранц- анатомия (Kranz — по-немецки «венок»).
У некоторых С4-растений хлороиласты клеток мезофилла имеют хорошо развитые граны, в то время как в клетках обкладки граны развиты слабо или отсутствуют совсем (рис. 7-18). Кроме того, в хлоропластах клеток обкладки вовремя фотосинтеза, как правило, образуются более крупные и многочисленные зерна крахмала, чем в хлоропластах мезофилла.
Рис. 7-19. Поперечный срез участка листа кукурузы (Zea mays). Как у типичного С4-растения, сосудистые пучки окружены крупными, содержащими хлоропласты клетками обкладки, которые в свою очередь окружены слоем клеток мезофилла. С4 -путь осуществляется в клетках мезофилла, а цикл Кальвина — в клетках обкладки
Эффективность С4-растений
Фиксация СО2 у С4-растений требует более высоких энергетических затрат, чем у С3-растений. В С4-пути на каждую ассимилированную молекулу СО2 должна образоваться молекула фосфоенолпирувата; при этом тратятся две высокоэнергетические фосфатные группы АТР. С4-растениям нужно затратить пять молекул АТР на фиксацию одной молекулы СО2, а С3-растениям — только три молекулы АТР.
Может возникнуть вопрос, почему у С4-растений в процессе эволюции возник такой сложный и энергетически дорогой способ обеспечения цикла Кальвина углекислотой. Это станет ясно, если уточнить, что фотосинтез у С3-растений всегда сопровождается фотодыханием — процессом, в ходе которого на свету потребляется кислород и выделяется СО2 (рис. 7-20). Фотодыхание — это расточительный процесс. По сравнению с митохондриальным дыханием фотодыхание не сопровождается окислительным фосфорилированием и не дает АТР. Более того, фотодыхание отвлекает восстановитель, образуемый в световой стадии, от биосинтеза глюкозы на восстановление кислорода. При нормальных атмосферных условиях до 50% углерода, ассимилированного при фотосинтезе у С3-растений, может реокисляться до СО2 в процессе фотодыхания. Таким образом, столь активное фотодыхание у С3-растений ограничивает в значительной степени их эффективность, а у С4-растсний оно практически отсутствует.
Основной субстрат, окисляемый при фотодыхании у С3- растений, — это гликолевая кислота. Она окисляется в пероксисомах фотосинтезирующих клеток и образуется при окислительном расщеплении рибулозобисфосфата с помощью рубилозобисфосфаткарбоксилазы — основного фермента, превращающего СО2 в фосфоглицерат. Как это происходит?
Рибулозобисфосфаткарбоксилаза может способствовать взаимодействию РБФ либо с СО2, либо с О2. Когда концентрация СО2 высокая, а кислорода — относительно низкая, то рибулозобисфосфаткарбоксилаза фиксирует СО2 на РБФ с образованием фосфоглицерата. В случае низкой концентрации СО2 и относительно высокой концентрации О2 фермент проявляет оксигеназную активность и «сажает» О2 на РБФ; при этом образуется фосфогликолевая кислота и фосфоглицерат вместо двух молекул фосфоглицерата, образуемых в норме при карбоксилировании. Фосфогликолевая кислота затем превращается в гликолевую кислоту — субстрат фотодыхания.
Высокая концентрация СО2 и низкая О2 ограничивают фотодыхание. Соответственно С4-растения имеют значительное преимущество перед С3-растениями, поскольку СО2, фиксируемый в С4-пути, перекачивается активно из клеток мезофилла в клетки обкладки, тем самым поддерживая высокое отношение СО2/О2 в участках, где сосредоточена активность рибулозобисфосфаткарбоксилазы. Это благоприятствует реакции карбоксилирования. Кроме того, цикл Кальвина и фотодыхание проходят во внутреннем слое клеток обкладок проводящих пучков, и СО2, выделяемый при фотодыхании, может быть рефиксирован в наружном слое клеток мезофилла в С4-пути, который здесь осуществляется. Таким образом, СО2, образующийся при фотодыхании, не выходит из листа. Более того, С4-растения по сравнению с С3-растсниями лучше утилизируют внешний СО2; это обусловлено тем, что фосфоенолпируваткарбоксилаза не ингибируется О2. В результате скорость нетто-фотосинтеза у С4- трав, таких, как кукуруза (Zea mays), сахарный тростник (Saccharum officinale), сорго (Sorghum vulgare), может быть в 2 — 3 раза выше, чем у С3-трав, таких, как пшеница (Triticum aestivum), рожь (Secale cereale), овес (Avena sativa) и рис (Oryza sativa).
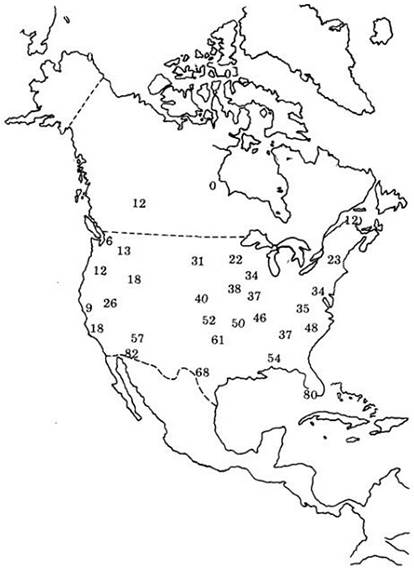
С4-растения появились в процессе эволюции преимущественно в тропиках; особенно хорошо они адаптированы к высокой инсоляции, повышенным температурам и засухе. Оптимальная температура для осуществления фотосинтеза у С4-растений выше, чем у С3-растений; С4-растения процветают даже при температурах, которые были бы детальны для многих С3-видов. С4-растения, обладая более эффективным способом использования СО2, могут достигать той же скорости фотосинтеза, что и С3-растения, но с меньшей потерей воды, поскольку у них может быть открыто меньшее число устьиц. Анализ географического распределения С4- видов в Северной Америке показывает, что в основном они наиболее многочисленны в климатических зонах с высокими температурами. Однако существуют различия между однодольными и двудольными в характере реакции на высокую температуру. Например, С4-злаки преобладают в районах с очень высокой температурой, приходящейся на вегетационный сезон (рис. 7-21). В то же время С4-двудольные широко распространены в тех районах, где вегетационный сезон характеризуется чрезмерной засушливостью.
Рис. 7-20. Реакции, катализируемые рибулозобисфосфаткарбоксилазой. Для реакции 1 благоприятны высокая концентрация СО2 и низкая концентрация кислорода. Реакция 2 обычно протекает при низкой концентрации СО2 и высокой концентрации кислорода (нормальные условия атмосферного воздуха)
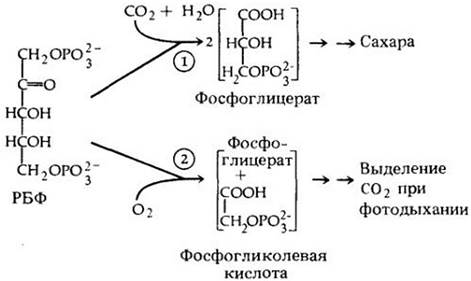
Рис. 7-21. Цифры указывают процент травянистых видов, фиксирующих СО2 по С4-пути, в 32 пунктах Северной Америки. Самый высокий процент характерен для районов с высокой температурой вегетационного периода
Яркую иллюстрацию особого типа роста у С4-растений можно найти на газонах, которые в северных районах США состоят из С3-трав, таких, как мятлик луговой (Роа pratensis) или полевица тонкая (Agrostis tenuis). Росичка кровяная (Digitaria sanguinalis), которая часто подавляет эти темно-зеленые тонколистные травы своими желтовато-зелеными широкими листьями, относится к С4-злакам и растет быстрее в жаркое лето, чем умеренные С3-злаки.
В настоящее время известно, что все растения с С4-фотосинтезом — это цветковые из 19 семейств (3 — однодольных и 16 — двудольных растений.), однако нет семейства, которое состояло бы только из С4-видов. Несомненно, этот путь фотосинтеза возникал в ходе эволюции многократно и независимо.
Метаболизм по типу толстянковых
Метаболизм органических кислот по типу толстянковых, обозначаемый как CAM-метаболизм1), возник в процессе эволюции у многих суккулентных растений, включая кактусы (Cactaccae) и толстянки (Crassulaceae). Растения относят к типу САМ, если их фотосинтезирующие клетки могут фиксировать СО2 в темноте с помощью фосфоенолпируваткарбоксилазы, образуя яблочную кислоту, которая запасается в вакуолях. В течение последующего светового периода яблочная кислота декарбоксилируется и СО2 переносится на РБФ цикла Кальвина в пределах той же клетки. Таким образом, CAM-растения, подобно С4-растениям, используют и С3- и С4-путь, но отличаются от С4-растений тем, что для них характерно разделение этих путей только во времени, но не в пространстве, как у С4-растений.
1По начальным буквам «Crassulacean acid metabolism» — Прим. ред.
CAM-растения сильно зависят от накопления углерода для фотосинтеза в ночные часы, поскольку их устьица закрыты в течение дня, чтобы предотвратить потерю воды. Это создает очевидное преимущество в условиях высокой инсоляции и нехватки воды, где обитают САМ-растения. Если все поглощение атмосферного СО2 у САМ-растений происходит ночью, то эффективность использования воды у этих растений может быть выше, чем у С3- или С4-растений. В течение периодов продолжительной засухи некоторые САМ-растения могут держать свои устьица закрытыми и ночью, и днем, поддерживая низкую скорость метаболизма за счет рефиксации СО2. При этом ночью фиксируется значительное количество СО2 в составе яблочной кислоты, а на следующий день СО2 высвобождается и вновь фиксируется в цикле Кальвина.
CAM-метаболизм более распространен среди сосудистых растений, чем С4-фотосинтез. Он характерен по крайней мере для 23 семейств цветковых растений, главным образом двудольных, включая такие привычные домашние растения, как каланхоэ Дегремона (Kalanchoë daigremontiana), восковое дерево (Hoya carnosa), сансевьера цейлонская (Sansevieria zeylanica). Но не все САМ-растения суккуленты; два примера меньшей суккулентности — ананас (Ananas comosus) и бородатый мох (Tillandsia usneoides); оба относятся к семейству бромелиевых (однодольные). Есть данные, что некоторые нецветковые растения проявляют признаки САМ-метаболизма, включая экзотическое голосеменное растение, Welwitschia mirabilis (см. рис. 18-35), полушник (Isoetes, рис. 17-19, 17-20) и некоторые папоротники.
Адаптивное значение фотосинтетических механизмов
Из приведенного выше обсуждения С3-, С4- и САМ-типов фотосинтеза становится очевидным, что фотосинтетический механизм не является единственным фактором, определяющим местообитание растения. Хотя это чрезвычайно важный фактор, все три типа фотосинтетических механизмов имеют свои преимущества и недостатки, и растение может успешно бороться за существование только в том случае, когда выгода того или иного типа фотосинтеза будет перевешивать другие факторы. Например, хотя С4-растения в целом более устойчивы к высоким температурам и засухе, чем С3-виды, при температуре ниже 25° С они не могут успешно конкурировать с ними, отчасти из-за большей чувствительности к холоду. Далее, CAM-растения лучше адаптированы к суровой засухе; они запасают воду, закрывая устьица в течение дня. Однако это свойство значительно уменьшает их способность поглощать и ассимилировать СО2. Поэтому CAM-растения растут медленно и слабо конкурируют с С3- и С4-видами при менее экстремальных условиях. Таким образом, каждый тип фотосинтеза растений в какой-то мере становится жертвой собственного механизма.