СОВРЕМЕННАЯ БОТАНИКА - П. РЕЙВН - 1990
РАЗДЕЛ III. ГЕНЕТИКА
ГЛАВА 8. ХИМИЯ И НАСЛЕДСТВЕННОСТЬ
Регуляция транскрипции генов
Транскрипция генов находится под тщательным контролем клетки. Даже у бактерий некоторые системы переключения действуют так, чтобы регулировать транскрипцию определенных генов в нужный момент и в необходимой степени. Эукариоты, по-видимому, обладают многими возможностями бактерий, хотя детали механизма регуляции транскрипции генов у эукариот менее известны.
Благодаря интенсивным исследованиям некоторых бактериальных генов мы много знаем о том, как регулируется их транскрипция. Пожалуй, лучше всего изучен механизм метаболизма лактозы у бактерии Escherichia coli. В норме эта бактерия не встречается с дисахаридом лактозой и поэтому не синтезирует ферментов, необходимых для ее метаболизма. Если лактоза добавлена в культуральную среду, то бактерии начинают в большом количестве синтезировать фермент β-галактозидазу, которая расщепляет дисахарид на глюкозу и галактозу (рис. 8-11).
Рис. 8-11. У бактерии Escherichia coli для расщепления молекулы лактозы на галактозу и глюкозу требуется фермент β-галактозидазаб β-Галактозидаза — индуцируемый фермент, т. е. его образование регулируется индуктором, в этом случае субстратом — дисахаридом лактозой
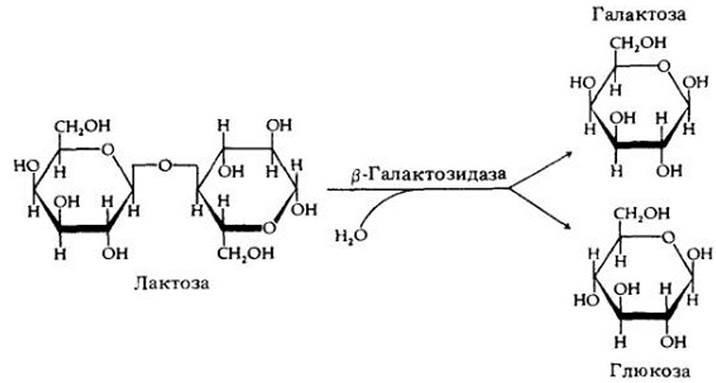
Таким образом получается, что лактозный субстрат вызывает, или индуцирует, образование фермента, необходимого для его расщепления. Известно много случаев подобной индукции синтеза ферментов энергетически богатым субстратом. В других случаях фермент, участвующий в синтезе определенной аминокислоты или другого метаболита, не образуется в клетке в присутствии этой аминокислоты; в подобной репрессии синтеза фермента ключевыми метаболитами участвует та же система контроля, что и в индукции.
Оперон
Франсуа Жакоб и Жак Моно, работавшие во Франции, разработали концепцию оперона, объясняющую, каким образом бактериальная клетка регулирует биосинтез ферментов. За это исследование в 1965 г. они были удостоены Нобелевской премии. Оперон — это группа структурных генов, расположенных вдоль одного фрагмента ДНК; он является единицей трансляции; экспрессия всех его генов согласована, поскольку они транскрибируются вместе с образованием одной молекулы мРНК. Например, lac-оперон Е. coli содержит ген β-галактозидазы и два других гена, продукты которых участвуют в метаболизме лактозы.
Одновременная работа ферментов, закодированных всеми генами laс-оперона, необходима, чтобы бактерия утилизировала лактозу. В каждой клетке, растущей на среде, содержащей лактозу, обнаруживается около 3000 молекул β-галактозидазы, что составляет около 3% всего клеточного белка. В отсутствие лактозы не происходит экспрессии генов laс-оперона, поскольку особый белок, называемый репрессором, блокирует ее. Репрессор связывается с ДНК в определенном месте, а именно между самими генами и областью присоединения РНК-полимеразы к ДНК. Когда в клетку поступает лактоза, она связывается с белком-репрессором, меняя его форму таким образом, что он теряет сродство к ДНК. Это позволяет полимеразе беспрепятственно перемещаться вдоль ДНК, транскрибируя гены lac-оперона, кодирующие ферменты.
У эукариот не обнаружено оперонов, т. е. групп связанных генов, которые транскрибируются сообща с образованием одной молекулы мРНК, как у бактерий. Каждая цитоплазматическая мРНК эукариотической клетки несет информацию о синтезе только одного белка. Однако у растений встречаются системы регуляции. Мутации, проявляющиеся в неограниченном образовании групп ферментов, были обнаружены у кукурузы (Zea mays) и ослинника (Oenothera).
Ингибирование по типу обратной связи
Кроме генетических механизмов, которые изменяют функции клеток, контролируя синтез отдельных ферментов, известен ряд физиологических контролирующих систем, осуществляющих прямое ингибирование посредством обратной связи, иными словами, путем контроля активности ферментов.
Тонко отрегулированные контролирующие системы, например, ингибирование конечным продуктом, служат примером аллостерических взаимодействий. Связывание особой молекулы (аллостерического эффектора) с определенным участком белка может так повлиять на слабые взаимодействия, детерминирующие его третичную структуру (см. гл. 3), что она изменится. Когда молекула белка претерпевает конформационные изменения, то может быть затронут и изменен участок, который позволяет белку функционировать в качестве фермента. Подобные взаимодействия могут контролировать уровень специфической активности клеточных ферментов, тем самым оказывая влияние на функциональную активность клетки. Связывание лактозы с белком- репрессором, обсуждавшееся выше, приводит к аллостерическим изменениям этого белка. Аллостерическим эффектором в некоторых случаях может быть конечный продукт того метаболического пути, который начинается с реакции, катализируемой аллостерическим ферментом.
Контроль развития растений
Те генетические процессы, которые мы обсудили, в конечном итоге ответственны за проявление отдельных признаков организма. Однако становлению признаков, характерных для взрослого организма, непременно предшествуют дифференцировка и развитие. Дифференцировка — это процесс, благодаря которому неспециализированные клетки или ткани постепенно изменяются, начинают выполнять специфические функции и приобретают характерную структуру. Под развитием подразумевают организацию сложного набора тканей, которые образуют взрослый организм. Взрослый организм представлен сложной совокупностью многих различных тканей, которые находятся в определенных физиологических и морфологических отношениях друг с другом. В конечном счете все они происходят из одной и той же клетки. У диплоидных организмов — это оплодотворенная яйцеклетка, или зигота. По мере ее развития происходит организация все более сложных тканей, что требует и более тщательного контроля.
Процессы развития животных и растений радикально отличаются. Развитие животных происходит по определенному плану в точной последовательности; порядок и согласованность во времени — решающие факторы для достижения конечного результата. На каждой стадии осуществляется строгий контроль экспрессии генов. У животных внешние условия, как правило, не сильно влияют на ход развития; по достижении взрослого состояния оно у них заканчивается.
У растений, напротив, процесс развития непрерывен: растения развиваются на протяжении всей своей жизни. На их развитие непосредственно влияют как внешние факторы (что минимально у животных), так и внутренние. Тканеспецифичная дифференцировка растений управляется гормонами, синтез которых чувствителен к изменениям условий окружающей среды (см. гл. 24). Способность растений реагировать на внешние факторы способствует адаптации растений к конкретным местообитаниям. Это важный фактор, если иметь в виду неспособность растений «искать» более благоприятные условия, перемещаясь с места на место.
У растений почти всякая дифференцировка обратима при условии, если дифференцированная клетка жива, в протопласте сохранилось ядро и не образовалась вторичная оболочка. С этим связано отсутствие у растений жесткого плана развития. У животных такая обратимость возможна достаточно редко.
Обратимость развития растений может быть проиллюстрирована экспериментальными данными. Немецкий ботаник Готлиб Габерландт еще в 1902 г. высказал мысль, что все живые растительные клетки тотипотентны: это значит, что каждая из них обладает потенциальной возможностью стать любой другой растительной клеткой. Габерландт считал, что один кусочек растительной ткани или даже отдельная клетка способны образовать целое взрослое растение, но он не смог это практически осуществить. Прошло более половины столетия, прежде чем эта гипотеза была четко доказана.
В целом проблема свелась к незнанию тех веществ, которые надо добавить в ростовую среду. В 1950-х годах физиолог растений Корнеллского университета Ф. К. Стьюард изолировал маленький кусочек флоэмы корня моркови (Daucus carota) и поместил его в жидкую культуральную среду во вращающейся колбе. Среда содержала сахарозу и минеральные элементы, необходимые для роста растений (см. гл. 26), а также некоторые витамины (органические вещества, которые растение синтезировать не может). Однако для роста и дифференцировки требовались еще какие-то вещества. Стьюард обнаружил их в кокосовом молоке, которое известно богатым содержанием ростовых веществ, хотя их природа к тому времени не была выяснена.
Во вращающейся колбе индивидуальные клетки постоянно отделялись от растущей клеточной массы и свободно плавали в среде. Эти одиночные клетки были способны расти и делиться, образуя небольшие группы. Еще давно Стьюард заметил, что многие из этих новых групп клеток образовывали корни. Если клетки оставить во вращающейся колбе, они перестают дифференцироваться, но при переносе на твердую среду (в данных опытах на агар) в некоторых группах образовывался побег (рис. 8-12). После перенесения на почву маленькие растения покрываются листьями, цветут и образуют семена.
Рис. 8-12. Две почки, образовавшиеся на недифференцированной ткани (каллусе) герани после обработки двумя гормонами — ауксином и цитокинином. Каллусы некоторых растений продолжают расти либо как недифференцированная ткань, либо как корни или почки в зависимости от соотношения гормонов

Приведенные результаты показывают, что по крайней мере некоторые клетки дифференцированной ткани флоэмы содержат всю генетическую информацию для развития целого растения, хотя она и не проявляется. Опыты свидетельствуют и о том, что такие дифференцированные клетки способны экспрессировать часть своего ранее не проявлявшегося генетического материала, когда из окружающей среды поступают соответствующие сигналы. (Гормональный контроль такого способа дифференцировки обсужден в гл. 24.) Получив данные результаты, Стьюард, наконец, подтвердил гипотезу Габерландта.
Форма и структура взрослого организма определена программой, заложенной в его генах. Однако признаки такого организма полностью проявляются только в ходе сложных процессов дифференцировки и развития. Процессы контролируются взаимодействием продуктов, транслируемых в ходе развития организма.
Влияние цитоплазмы на дифференцировку
У многих организмов компоненты цитоплазмы играют непосредственную роль в процессе клеточной дифференцировки; среди них органеллы, например, пластиды и митохондрии, которые содержат собственную ДНК (см. гл. 2). Если в ходе клеточного деления эти органеллы распределяются между дочерними клетками неравномерно, то судьба последующих «поколений» клеток может сильно различаться. Этим объясняются, в частности, многие из «материнских эффектов», известных в генетической литературе, — различия идентичных гибридов, обусловленные влиянием материнского организма.
Сходным образом важную роль в дифференцировке играют химические градиенты, т. е. изменения концентрации веществ в различных частях клетки. Например, у бурой водоросли Fucus (см. рис. 15-9,5) градиент запасенных нерастворимых пищевых частиц, по-видимому, устанавливается в зиготе под действием силы тяжести или, может быть, электрохимических сил. Этот градиент определяет положение веретена в первом делении зиготы, а, следовательно, и «судьбу» двух клеточных поколений, которым дает начало это деление. Неравное клеточное деление может быть весьма существенным для распределения различных элементов цитоплазмы и определения судьбы клеточных поколений.
В клетках животных и растений многие вещества непрерывно диффундируют с неодинаковой скоростью в различных направлениях и разного типа ткани часто расположены в непосредственной близости одна от другой. Поэтому эффект, обнаруженный у Fucus, может быть выявлен и у других более просто устроенных организмов. К чрезвычайно тонкому контролю процесса развития могут привести не только градиенты веществ в отдельных клетках и тканях; дифференцировка любой клетки способна в большой мерс определяться ее положением в теле развивающегося растения или животного. Некоторые пути взаимодействия гормонов и других факторов в развитии растений обсуждены в разд. VI.