СОВРЕМЕННАЯ БОТАНИКА - П. РЕЙВН - 1990
РАЗДЕЛ III. ГЕНЕТИКА
ГЛАВА 8. ХИМИЯ И НАСЛЕДСТВЕННОСТЬ
Заключение
Ключом к пониманию процессов наследственности стали исследования, проведенные на молекулярном уровне в начале 1950-х годов и открывшие роль ДНК. Ряд фактов убедительно свидетельствует о том, что генетическая информация заключена именно в ДНК, а в 1953 г. Джеймс Уотсон и Фрэнсис Крик создали ее молекулярную модель. Это открытие обеспечило быстрый прогресс в области молекулярной генетики, и тайная работа генетического механизма клетки теперь может быть описана довольно подробно.
В процессе репликации ДНК на каждой исходной цепи синтезируется комплементарная дочерняя цепь. Разнообразные ферменты действуют согласованно, раскручивая витки ДНК, расплетая двойную спираль и добавляя новые основания к каждой из двух строящихся дочерних цепей.
Генетическая информация передается с помощью информационной (матричной) РНК. Длинные молекулы мРНК синтезируются по принципу комплементарности на одной цепи ДНК, а затем переносятся на цитоплазматические рибосомы. Этот процесс называют транскрипцией; он находится под жестким генетическим контролем. Последовательность из трех нуклеотидов в молекуле мРНК кодирует специфическую аминокислоту.
На рибосомах мРНК взаимодействует с маленькими молекулами — транспортными РНК, которые связаны с определенными аминокислотами. Каждая тРНК имеет последовательность из трех оснований (антикодон), комплементарную кодону мРНК. Молекула тРНК связывается с помощью комплементарного антикодона с мРНК, а доставленная ею аминокислота связывается с концом растущей полипептидной цепи. Таким образом, аминокислота теперь соединена с белковой цепью пептидной связью, образованной с помощью специальных ферментов. Процесс синтеза белка называют трансляцией. Каждая из 20 аминокислот кодируется триплетом оснований (кодоном) в мРНК. Последовательность аминокислот в белке определена последовательностью кодонов в молекуле мРНК, которая и управляет синтезом этого конкретного белка. В конечном счете последовательность кодонов мРНК зависит от последовательности оснований ДНК, с которой мРНК была транскрибирована. Большинству аминокислот соответствуют три или четыре альтернативных кодона, каждому из которых соответствует своя тРНК.
Не весь генетический материал несет информацию об аминокислотной последовательности белков. Большая часть генетической информации ядерной мРНК эукариот транскрибирована с фрагментов ДНК, названных нитронами; эти фрагменты вырезаются из мРНК до того, как она попадет в цитоплазму. Остальные фрагменты мРНК, транскрибированные с участков ДНК, называемых экзонами, сшиваются в ядре до выхода мРНК в цитоплазму.
Регуляция экспрессии некоторых систем бактериальных генов, например, lac-оперона Escherichia coli, проста: одна система активирует транскрипцию в присутствии потенциального субстрата — «индуктора» (лактозы), тогда как другая выключает транскрипцию в присутствии избытка продукта — глюкозы. У эукариот последовательность процессов развития более сложна.
У растений транскрипция генов тесно связана с процессом развития и организована поэтапно; каждому этапу соответствует свой особый путь клеточной дифференцировки. На выбор конкретного пути влияют окружающие условия, и он обратим. В принципе любая дифференцированная клетка, сохранившая протопласт с ядром, может быть дедифференцирована и стимулирована к образованию целого растения.
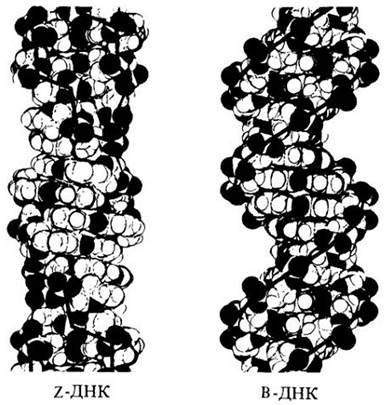
Приложение 1. Правозакрученная и левозакрученная
Непрерывная двойная спираль ДНК, описанная Дж. Уотсоном и Ф. Криком, закручивается в правую сторону (на рисунке справа). Однако в конце 1970-х годов Алекс Рич и его коллеги в Массачусетском технологическом институте обнаружили вторую устойчивую конфигурацию ДНК, а именно закрученную в левую сторону (на рисунке слева). Открытие двух форм ДНК стало возможным благодаря усовершенствованию методов химического анализа. На изображенных моделях «остов» полинуклеотидных цепей ДНК показан черной линией. В левозакрученной ДНК остов зигзагообразен, и поэтому такую форму называют Z-ДНК. В определенных условиях Z-ДНК и В-ДНК (правозакрученная форма) могут переходить одна в другую. Показано, что присутствие Z-ДHK коррелирует с регулируемой транскрипцией определенных генов.
Приложение 2. Контроль многоклеточной дифференцировки
Дифференцировка — это процесс развития, в течение которого относительно неспециализированные клетки или ткани претерпевают постепенные изменения и становятся более специализированными по функции и структуре. Примером хорошо изученной системы, иллюстрирующей процесс дифференцировки, может служить клеточный слизевик Dictyosteliumdiscoideum. Его жизненный цикл позволяет проследить, как воздействия внешних факторов на развивающиеся клетки или ткани могут сказываться в конечном счете на признаках достигшего зрелости организма.
Клеточные слизевики обычно существуют в виде свободноживущих амебовидных клеток, или миксамеб, которые питаются бактериями, окружая и поглощая их (см. рис. 4-11). Они размножаются делением и проявляют слабую морфологическую дифференцировку, пока не исчерпается имеющийся запас бактерий. В ответ на голодание клеточный слизевик образует споры. Отдельные клетки сначала группируются, образуя подвижную массу, похожую на слизевик и называемую псевдоплазмодием. Перед тем как образовать и рассеять споры, псевдоплазмодий мигрирует на новое место. Эта удивительная стадия жизненного цикла предотвращает попадание спор на старое место, где уже больше нет бактерий.
Агрегация миксамеб происходит в тот момент, когда одна или несколько голодающих клеток начинают выделять в среду циклический аденозинмонофосфат (сАМР). Это вещество диффундирует и создает градиент концентрации, который вызывает движение окружающих клеток по направлению к той, которая выделяет с АМР. Секретирующие клетки в свою очередь побуждаются к выделению новой порции сАМР после пятиминутного интервала, и, таким образом, привлекаемые клетки поступают по крайней мере тремя волнами. Когда клетки скапливаются у центра агрегации, их плазматические мембраны становятся липкими и склеиваются друг с другом, что приводит к образованию псевдоплазмодия, окруженного целлюлозной оболочкой.
А. Питание амеб. Светло-серые области в центре каждой клетки — ядра, а белые области — сократительные вакуоли. Б. Агрегация амеб. Направление их движения указано стрелкой. В. Мигрирующие псевдоплазмодии, состоящие из большого количества амеб. Каждая, подобная слизню, масса клеток откладывает вокруг себя толстый чехол из слизи, который уплотняется. Г. По окончании миграции псевдоплазмодий начинает расти вертикально, дифференцируясь на ножку и многочисленные споры, собранные в виде капли (Д)
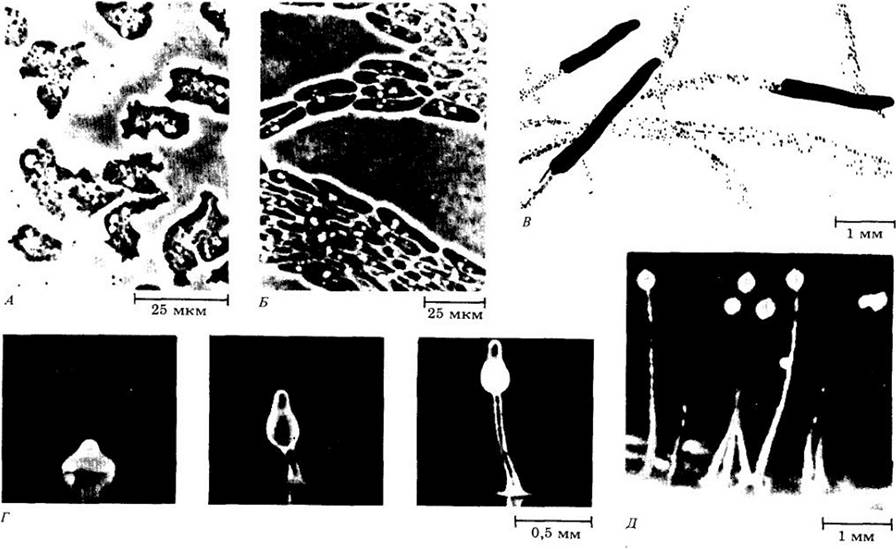
Конечная судьба отдельной клетки определяется в ходе развития ее положением в конгломерате клеток. Первые объединившиеся клетки обычно дают начало передней части псевдоплазмодия, тогда как вступившие в ассоциацию последними образуют его основание. Когда миграция прекращается, клетки апикальной области формируют ножку развивающегося «плодового тела». Затем клетки задней части псевдоплазмодия перемещаются к верхушке ножки и становятся спорами. Наконец базальный диск и клетки ножки погибают и споры рассеиваются. При попадании на теплый влажный субстрат они прорастают. Из каждой споры развивается одна миксамеба, и цикл повторяется снова (см. также гл. 14).
Итак, даже в этой относительно простой эукариотической системе мы встречаемся с миграцией определенных типов клеток, локальной гибелью клеток и распознаванием «тканей». Гораздо более сложные системы с более точным контролем характеризуют клеточную дифференцировку у растений.