СОВРЕМЕННАЯ БОТАНИКА - П. РЕЙВН - 1990
РАЗДЕЛ III. ГЕНЕТИКА
ГЛАВА 9. ГЕНЕТИКА ЭУКАРИОТ
Мейоз
Мейоз протекает в специализированных диплоидных клетках и только в определенные моменты жизненного цикла данного организма. В результате мейоза и цитокинеза из одной диплоидной клетки образуются четыре гаплоидные — гаметы или споры. Гамета — это клетка, которая, сливаясь с другой гаметой, образует диплоидную зиготу. Зигота затем может делиться либо мейотически, образуя четыре гаплоидные клетки, либо митотически, давая начало многоклеточному диплоидному организму. Гаплоидные клетки могут функционировать и как самостоятельные одноклеточные организмы. Если сформировался многоклеточный диплоидный организм, то в большинстве случаев рано или поздно он будет образовывать гаплоидные споры или гаметы посредством мейоза. Спора — это клетка, которая может развиться в целый организм без объединения с другой клеткой. Споры часто делятся митотически, и в результате формируется гаплоидный многоклеточный организм, у которого путем митоза образуются гаметы (см. рис. 10-11).
Первое деление мейоза
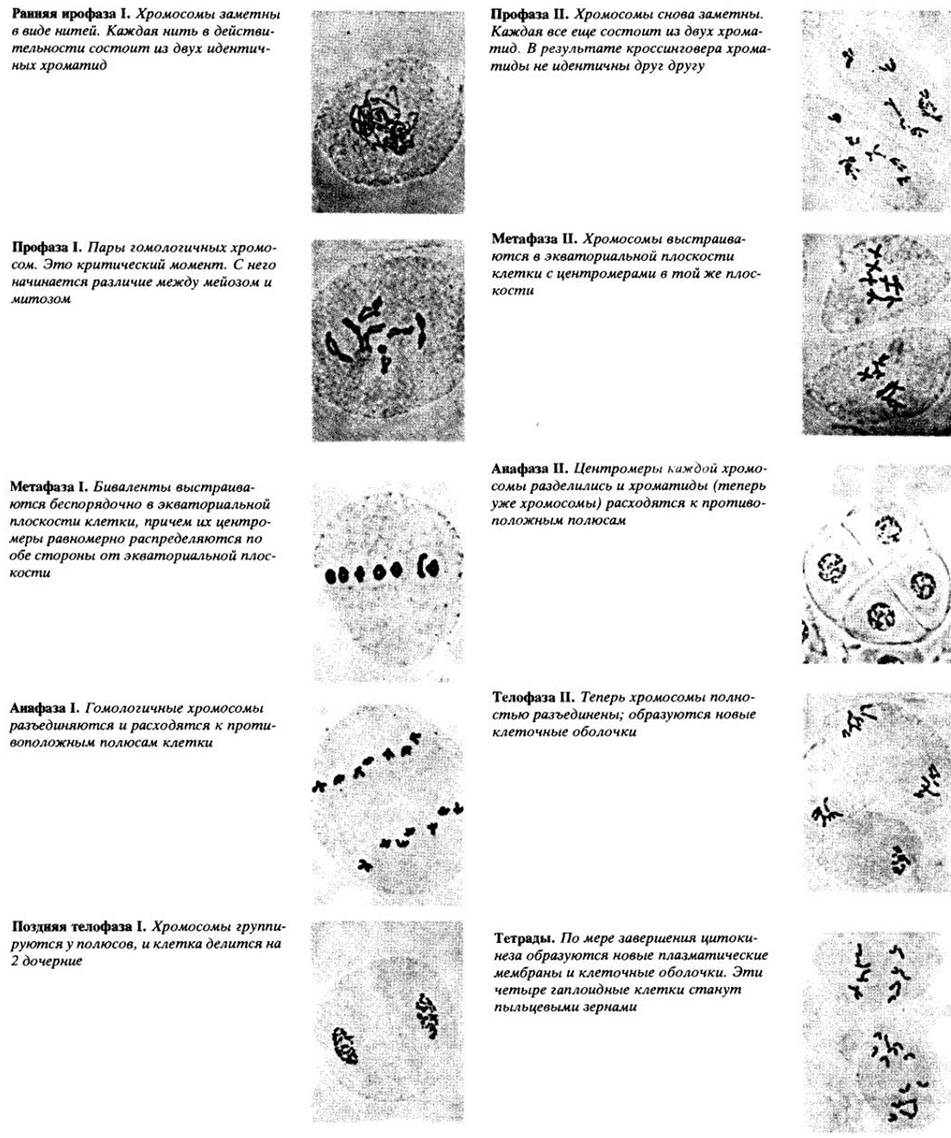
Мейоз состоит из двух последовательных делений ядра. Рис. 9-6 поможет нам проследить процессы, описанные в следующих параграфах.
В профазе I (профазе первого мейотического деления) хромосомы принимают вид длинных тонких нитей. Как и в митозе (см. гл. 2), хромосомы уже удвоились во время предшествующей интерфазы. Следовательно, в начале профазы I каждая хромосома состоит из двух идентичных хроматид, скрепленных центромерой. Однако на этой ранней стадии мейоза каждая хромосома выглядит как единое целое.
Прежде чем станут заметны хроматиды, гомологичные хромосомы спариваются (конъюгируют) друг с другом. Конъюгация происходит очень точно, начинаясь с одного или нескольких участков, она продолжается вдоль всей хромосомы подобно застегиванию молнии. Каждый гомолог происходит от одного родителя и состоит из двух идентичных хроматид. Таким образом, гомологичные пары состоят из четырех хроматид. Конъюгация гомологичных хромосом является обязательной частью мейоза; в гаплоидных клетках она происходить не может, потому что в них отсутствуют гомологи. Сам процесс спаривания называют синопсисом, а объединившиеся пары гомологичных хромосом — бивалентами.
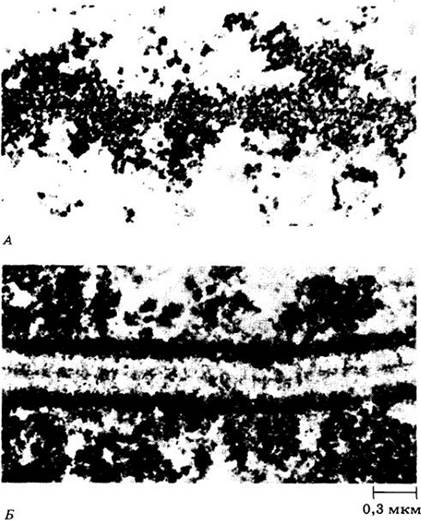
Во время профазы I хромосомы укорачиваются и утолщаются. Под электронным микроскопом в каждой хромосоме можно видеть темноокрашенное осевое белковое тело (рис. 9-3, А). В середине профазы осевые тела гомологичных хромосом сближаются друг с другом до предельного расстояния 0,1 мкм, образуя синаптонемальный комплекс (рис. 9-3, Б).
Рис. 9-3. А. Фрагмент хромосомы Lilium в профазе 1 до конъюгации. Обратите внимание на плотное осевое тело, состоящее главным образом из белков. Возможно, оно организует генетический материал, готовя его к спариванию и генному обмену. Б. Синаптонемальный комплекс в биваленте Lilium. Из четырех хроматид видны только две
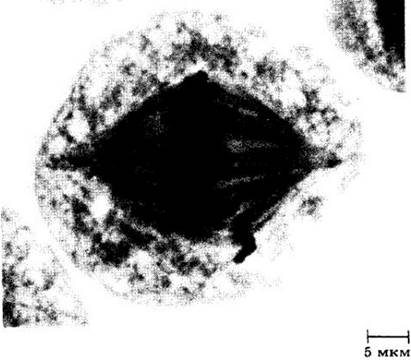
На удачно выбранном материале в это время можно видеть, что каждый бивалент состоит из четырех хроматид, по две в каждой хромосоме. В синаптонемальном комплексе происходит обмен участками между гомологичными хроматидами. Этот процесс, называемый кроссинговером, приводит к тому, что хроматиды теперь имеют иной состав генов. Рис. 9-4 наглядно иллюстрирует процесс кроссинговера; видимая Х-образная структура называется хиазмой.
Рис. 9-4. Конъюгирующие хромосомы прямокрылого Chorhippus parallelus. Можно видеть вариации в числе хиазм

К концу профазы I синаптонемальный комплекс распадается. Затем разрушается ядерная оболочка и обычно исчезают ядрышки, так как приостанавливается синтез РНК. Гомологичные хромосомы разъединяются, однако хроматиды удерживаются вместе в хиазмах (местах контакта) и расходятся очень медленно. По мере расхождения хроматид некоторые хиазмы смещаются к концу плеча хромосомы. В каждом плече хромосомы можно встретить одну или более хиазм, или одну на весь бивалент; структура отдельных бивалентов может быть самой разнообразной в зависимости от числа хиазм (рис. 9-4).
В метафазе I становится заметной веретеновидная структура, образуемая микротрубочками (рис. 9-5). В ходе мейоза к центромерам хромосом каждого бивалента прикрепляются отдельные микротрубочки. Затем пары хромосом перемещаются в экваториальную плоскость клетки, где выстраиваются в случайном порядке. Центромеры гомологичных хромосом располагаются в противоположных сторонах от экваториальной плоскости; в метафазе митоза, как мы уже видели, напротив, центромеры отдельных хромосом располагаются в экваториальной плоскости.
Рис. 9-5. Веретено в материнской клетке пыльцы пшеницы (Тriticum aestivum) во время метафазы I мейоза
Анафаза I начинается с расхождения гомологичных хромосом и движения их в направлении полюсов. (Отметьте опять отличие от митоза. В анафазе митоза центромеры делятся и идентичные хроматиды расходятся.) В анафазе I мейоза центромеры не делятся, хроматиды остаются вместе, а разъединяются гомологичные хромосомы. Однако из-за обмена фрагментами в результате кроссинговера хроматиды не идентичны, как в начале мейоза.
В телофазе I спирализация хромосом ослабевает, они удлиняются и снова становятся неразличимыми. По мере того как телофаза постепенно переходит в интерфазу, из эндоплазматического ретикулума возникает ядерная оболочка. Наконец вновь образуется ядрышко и возобновляется синтез белка. Однако у многих организмов интерфаза отсутствует, в этом случае хромосомы более или менее непосредственно переходят от телофазы I к профазе II второго мейотического деления.
Второе деление мейоза
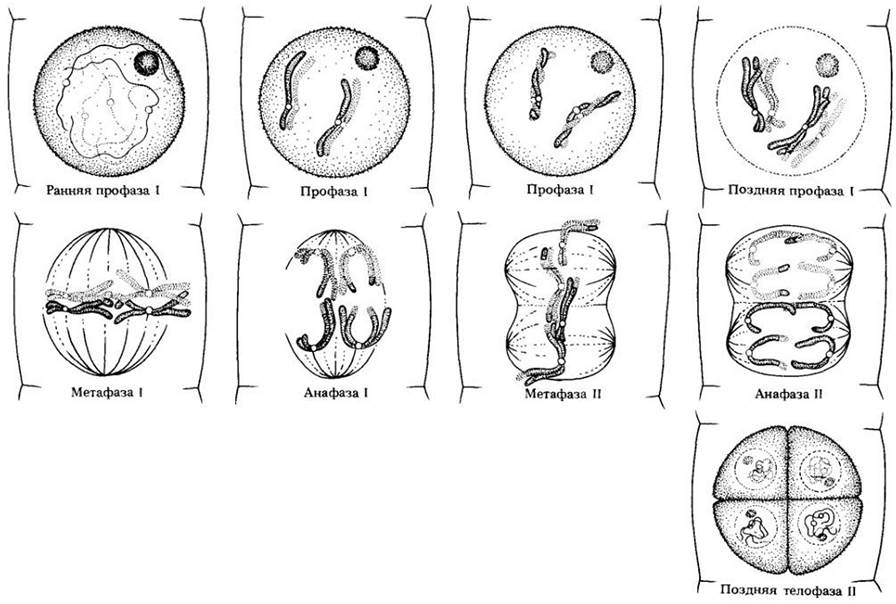
В начале второго мейотического деления хроматиды все еще связаны центромерами. Это деление похоже на митоз: если в телофазе I образовывалась ядерная оболочка, то теперь она разрушается, и к концу профазы II исчезает ядрышко. В метафазе // снова можно увидеть веретено и хромосомы, состоящие из двух хроматид. Хромосомы прикреплены центромерами к нитям веретена и выстраиваются в экваториальной плоскости. В анафазе II центромеры делятся и расходятся, а сестринские хроматиды, ставшие теперь хромосомами, движутся к противоположным полюсам (см. рис. 8-1). В телофазе II образуются новые ядерные оболочки и ядрышки, сжатие хромосом ослабевает и в интерфазном ядре они становятся невидимыми. Общая схема мейоза показана на рис. 9-6.
Рис. 9-6. Схематическое изображение мейоза с двумя парами хромосом. Показаны не все стадии.
Профаза I: хромосомы становятся заметными в виде длинных нитей; гомологичные хромосомы сближаются в пары, пары обвивают друг друга, и конъюгирующие хромосомы становятся очень короткими.
Метафаза I: спаренные хромосомы располагаются в экваториальной плоскости, их центромеры направлены в обе стороны от нее.
Анафаза I: конъюгирующие хромосомы разъединяются и движутся к противоположным полюсам.
Второе деление мейоза по-существу представляет собой митоз.
Метафаза II: хромосомы выстроились в экваториальной плоскости; их центромеры лежат здесь же.
Анафаза II: центромеры делятся, хроматиды расходятся и движутся к противоположным полюсам.
Телофаза II: хромосомы закончили движение; образовались четыре новых ядра, каждое с гаплоидным числом хромосом.
Мейоз у пырея (Аgropyron cristatum), n = 7, показан далее
Значение мейоза
Конечный результат мейоза таков, что генетический материал, присутствующий в диплоидном ядре, удваивается только один раз, но делится дважды. Поэтому в каждой дочерней клетке оказывается только половина того числа хромосом, которое присутствовало в диплоидном ядре. Более важны генетические последствия мейоза. В метафазе I ориентация бивалентов случайна, т. с. хромосомы одного родителя случайно распределяются между двумя новыми ядрами. Кроме того, благодаря кроссинговеру каждая хромосома часто состоит из фрагментов хромосом обоих родителей. Если исходная диплоидная клетка имела две пары гомологичных хромосом, п=2, то существуют четыре возможности, согласно которым хромосомные пары могут выстроиться в метафазе, и соответственно четыре варианта их распределения между гаплоидными клетками. Если n = 3, то имеется 8 возможностей; если n = 4, то 16. Общая формула — 2n. У человека n = 23, поэтому число возможных комбинаций составляет 223, что равно 8 388 608. У многих организмов число хромосом еще больше, чем у человека.
С увеличением числа хромосом все меньше вероятность восстановления набора хромосом, идентичного бывшему в исходном диплоидном ядре. Кроме того, наличие по крайней мере одной хиазмы в каждом биваленте делает практически невозможным получение в результате мейоза клетки, генетически тождественной любой из тех, слияние которых привело когда-то к формированию родительского организма. Три принципиальных отличия мейоза от митоза следующие (см. рис. 9-7):
Рис. 9-7. Сравнение мейоза и митоза
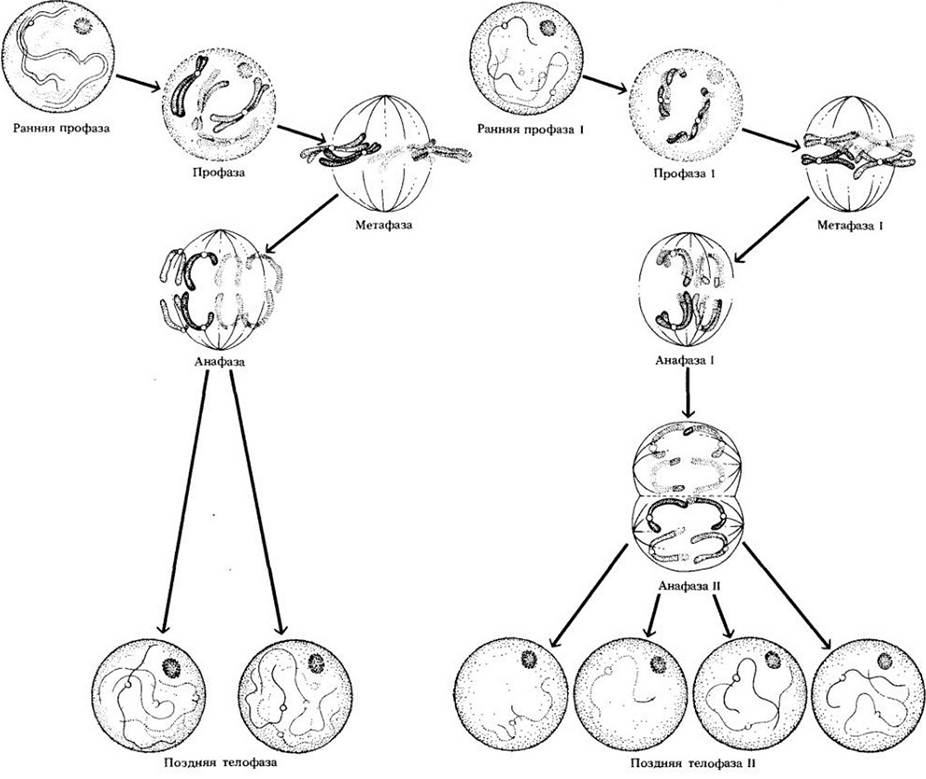
1. Несмотря на то что в процессе мейоза генетический материал удваивается только один раз, происходят два деления, приводящие к образованию четырех ядер.
2. Каждое из четырех ядер гаплоидно, т. е. содержит только половину числа хромосом, характерного для исходного диплоидного ядра.
3. Ядра, образованные в результате мейоза, содержат новые комбинации хромосом.
В результате мейоза образующиеся ядра отличаются от исходного в противоположность митозу, когда наборы хромосом дочерних ядер и материнского идентичны. Поведение хромосом в мейозе имеет глубокие генетические и эволюционные последствия. Благодаря мейозу и сингамии природные популяции диплоидных организмов далеко не однородны; они состоят из особей, которые различаются между собой по многим признакам.