СОВРЕМЕННАЯ БОТАНИКА - П. РЕЙВН - 1990
РАЗДЕЛ III. ГЕНЕТИКА
ГЛАВА 9. ГЕНЕТИКА ЭУКАРИОТ
Как детерминируются признаки?
На рис. 9-8 изображена кривая изменчивости длины початков растений кукурузы определенного сорта, но подобная кривая может отражать и распределение растений по весу, например, у выбранного наугад сорта пятнистых бобов, или распределение по высоте деревьев дуба. Факторы внешней среды, например, пролившийся на кукурузу дождь, могут изменить абсолютные величины, по которым строится такой график, но лишь редко могут повлиять на форму кривой.
Такого рода кривая, имеющая форму колокола, называется «нормальной», а характер изменчивости — непрерывным. Плавная симметричная форма кривой говорит о том, что популяция не может быть расчленена на ряд резко различающихся форм. Рост человека наследуется таким же образом. Большинство вариаций, наблюдаемых в природе, носят именно непрерывный характер, и те ученые, которые первыми пытались понять, как наследуются признаки растений и животных, были этим фактом озадачены. У растений наиболее важные для сельскохозяйственного производства признаки — размер, вес и высота — имеют непрерывную изменчивость. Теперь мы знаем, что такое наследование зависит от взаимодействия многих генов и в принципе поддается анализу путем установления индивидуальной роли каждого гена.
Рис. 9-8. Распределение длин початков кукурузы (Zea mays) сорта Black Mexican. Длина початка — пример фенотипического признака, который детерминируется взаимодействием нескольких генов. Этот признак обнаруживает непрерывную изменчивость; кривая изменчивости имеет форму колокола, а средняя величина признака приходится на середину кривой

Гениальность Грегора Менделя отчасти состояла в том, что он стал изучать не непрерывную изменчивость, которая нередко встречается в природе, а дискретные качественные признаки, определяемые одним геном. Такие признаки часто встречаются у тех растений, которые и предоставили Менделю прекрасный исследовательский материал; для растений вообще характерны самоопыляющиеся разновидности; скрещивания между ними произвести просто, и вырастить большое гибридное потомство не представляет особого труда (в отличие от многих групп животных). Имея удачный экспериментальный материал, Мендель мог решить те сложные проблемы, которые ставили в тупик его современников, включая Чарльза Дарвина. Мендель работал в саду августинского монастыря св. Фомы в австрийском городе Брюнне (столице Моравии; теперь город Брно в Чехословакии) с 1856 по 1863 гг. Он тщательно выбирал свой главный объект для опытов — горох огородный (Pisum sativum). С юных лет Мендель собирал семена гороха с разными признаками и к 1856 г. уже хорошо знал, какие растения будут скрещиваться друг с другом, а какие — нет.
Мендель обратил внимание на четкие признаки — окраску цветков и форму семян. Затем он провел большое количество экспериментальных скрещиваний и проанализировал численные соотношения признаков среди потомства. Вероятно, немаловажным было и то, что Мендель изучал потомство не только первого поколения, но и последующих генераций. Кроме того, прежде чем скрестить растения гороха, он получал чистые линии по каждому интересовавшему его признаку; это было легким делом, поскольку самоопыление у гороха происходит автоматически (рис. 9-9). Ученые, изучавшие процессы наследственности до Менделя, не исследовали такие четкие признаки и не подготавливали столь тщательно свой экспериментальный материал, а поэтому и результаты их исследований оставались сомнительными и с трудом поддавались анализу.
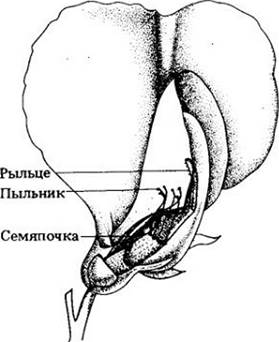
Рис. 9-9. В цветке пыльца образуется в пыльниках, а яйцеклетки — в семяпочках. В результате опыления пыльцевые зерна, нанесенные на рыльце пестика, прорастают по направлению к семяпочке, где происходит оплодотворение. У большинства видов в этом процессе участвует пыльца одного растения, попавшая на рыльце другого растения; это явление называют перекрестным опылением. Оплодотворенная яйцеклетка, или зигота, развивается в зародыш внутри семяпочки; созревшие семяпочка и зародыш образуют семя. В цветке гороха огородного рыльца и пыльники полностью скрыты лепестками. Поскольку цветок гороха в противоположность цветкам многих других растений не раскрывается, пока не произойдут опыление и оплодотворение, то в норме растение самоопыляется. Мендель в своих опытах по перекрестному опылению открывал бутон до того, как созреет пыльца, и пинцетом удалял пыльники. Затем проводил искусственное опыление, нанося на рыльце пыльцу, собранную с другого растения, которое обладало интересовавшими его признаками
Интерпретировать результаты опытов Менделя было удивительно просто. В табл. 9-1 перечислены семь признаков растений гороха, на которых ученый сосредоточил свое внимание. Скрещивания между растениями, различающимися только по одному признаку (как в опытах Менделя), называют моногибридными, а по двум признакам — дигибридными. Когда Мендель скрещивал растения с альтернативными признаками, он наблюдал, что всегда один из этих признаков не проявлялся в первом поколении гибридов (F1). Например, семена всех потомков от скрещивания растений с желтыми и зелеными семенами были желтыми, как и у одного из родительских растений. Признак желтой окраски семян, так же, как и другие признаки, проявляющиеся в первом поколении, Мендель назвал доминантными, а те, которые не проявляются в первом поколении, — рецессивными. В результате самоопыления растений первого поколения рецессивные признаки вновь проявлялись во втором поколении в соотношении три доминантных к одному рецессивному (табл. 9-1).
Таблица 9-1. Опыты Менделя с растениями гороха
Признак |
Доминантный |
Рецессивный |
Поколение F2 |
|
Доминантный |
Рецессивный |
|||
Форма семян (7)1 |
Круглая |
Морщинитая |
5474 |
1850 |
Окраска семян (1) |
Желтая |
Зеленая |
6022 |
2001 |
Положение цветка (4) |
Пазушное |
Терминальное |
621 |
207 |
Окраска цветка (1) |
Красная |
Белая |
705 |
224 |
Форма боба (4) |
Выпуклая |
Плоская |
882 |
299 |
Окраска боба (5) |
Зеленая |
Желтая |
428 |
152 |
Длина стебля (4) |
Длинный |
Короткий |
787 |
277 |
1Цифры в скобках указывают хромосому, на которой локализован ген данного признака.
Полученные результаты легко понять, имея представления о мейозе, но, когда Мендель проводил свои исследования, мейоз еще не был известен. Признаки диплоидного организма определяются взаимодействиями между аллелями. Аллель — одна, две или более альтернативных форм одного и того же гена. Аллели занимают одинаковые места (сайты, или локусы) на гомологичных хромосомах.
Рассмотрим скрещивание между растениями с белыми и красными цветками. Аллель белой окраски цветка, которая является рецессивным признаком, обозначим маленькой буквой w, а аллель красной окраски цветка (доминантного признака) — большой буквой W. В чистых линиях гороха, с которыми работал Мендель, растения с белыми цветками имели генотип ww. Растения с красными цветками имели генотип Особи, у которых два гена, определяющих данный признак, идентичны, т. е. особи с идентичными аллелями, называются гомозиготами (гомозиготными). При скрещивании растений с данными генотипами все растения в F1 получают аллель W от материнского растения с красными цветками и аллель лу — от растения с белыми цветками и, таким образом, имеют генотип Ww. Такие особи называются гетерозиготными по гену окраски цветка.
Во время мейоза у гетерозиготных особей образуются в равном количестве два вида гамет — W и w. Как показано на рис. 9-10, А, в результате рекомбинации гамет при скрещивании двух гетерозиготных растений в среднем на каждые четыре потомка одна особь будет иметь генотип WW, одна — ww и две особи — Ww. По внешнему виду, или фенотипу, гетерозиготные Ww растения не будут отличаться от гомозиготных WW растений, т. е. тоже будут иметь красные цветки. Продуктов аллеля от растения с красными цветками достаточно, чтобы замаскировать продукты аллеля от растений с белыми цветками, и это является обоснованием фенотипического соотношения три к одному, которое получил Мендель (табл. 9-1).
Рис. 9-10. А. Скрещивание растения гороха с двумя доминантными аллелями красной окраски цветка (WW) с растением с двумя рецессивными аллелями белой окраски цветка (ww). Гибриды первого поколения имели красные цветки, но заметьте, что их генотип был Ww. Поколение F2 показано внизу. Аллель W, будучи доминантным, определяет внешний вид, или фенотип, цветка. Только когда потомство получает аллель от каждого родителя (генотип ww), проявляется рецессивный признак (белые цветки). Отношение доминантного фенотипа к рецессивному в поколении F2 составляет 3:1, отношение генотипов — 1:2:1. Б. Анализирующее скрещивание растения с красными цветками с растением с белыми цветками. Хотя растения, имеющие красные цветки, фенотипически тождественны растениям с генотипом результаты анализирующего скрещивания показали, что они гетерозиготны (Ww) по гену окраски цветка; только гетерозиготный генотип мог дать наблюдавшиеся в потомстве генотипы. В этом случае отношение генотипов составило 1WW к 1Ww
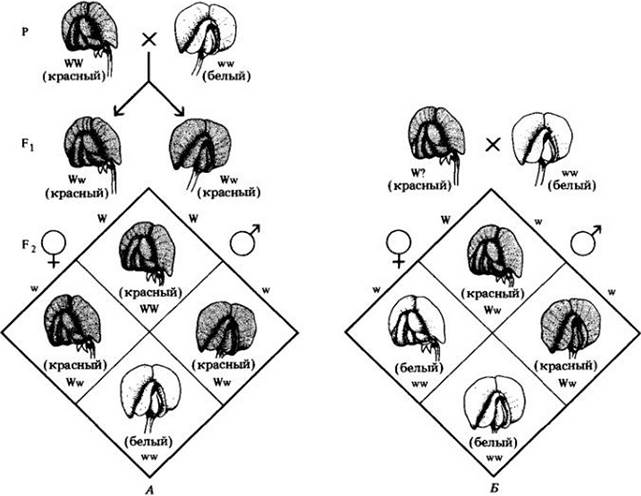
Как можно узнать, каков генотип растений с красными цветками — WW или Ww? Как показано на рис. 9-10, Б, это легко определить, скрестив такое растение с растением, имеющим белые цветки, и подсчитав потомство от этого скрещивания. Именно такой опыт и проделал Мендель. Скрещивание особи с доминантным признаком с гомозиготной рецессивной особью поэтому же признаку называют анализирующим.
Правило расщепления
Закономерность, установленную в описанных выше опытах, иногда называют первым законом Менделя или правилом расщепления. Согласно ему, наследственные признаки детерминируются дискретными факторами (т. е. генами), которые представлены парами, причем каждый ген пары унаследован от разных родителей. Во время мейоза пары генов разъединяются, или расщепляются. Следовательно, каждая гамета, образующаяся у зрелого потомка, содержит только один ген из той пары, которой обладал потомок. Данная концепция дискретности факторов объясняет, как признаки могут предаваться из поколения в поколение, не смешиваясь с другими признаками, и каким образом — «исчезать» и вновь появляться в следующих поколениях. Поэтому открытие правила расщепления имело огромное значение для понимания многих аспектов генетики и эволюции.
Неполное доминирование
В приведенных выше примерах присутствие доминантного аллеля маскировало присутствие второго, рецессивного аллеля. Однако доминантные и рецессивные признаки не всегда так четко разграничены. В случаях неполного доминирования, например, какой-либо признак проявляется у гетерозиготного растения в промежуточной между двумя гомозиготами форме, поскольку действие одного аллеля неполностью маскирует действие другого.
Например, у львиного зева при скрещивании растений с красными и белыми цветками получаются потомки с розовыми цветками. В потомстве от скрещивания растений первого поколения между собой эти признаки снова расщепляются в следующем соотношении: одно растение с красными цветками (гомозиготное), два растения с розовыми цветками (гетерозиготные) и одно растение с белыми цветками (гомозиготное) (рис. 9-11). Таким образом, и случаи неполного доминирования подтверждают менделевское правило расщепления.
Рис. 9-11. При скрещивании растений львиного зева с красными цветками с растениями, имеющими белые цветки, получается потомство с розовыми цветками (Ww). Результаты скрещивания растений с розовыми цветками показывают, что гены остаются дискретными и расщепляются согласно отношению 1:2:1 и в генотипе, и в фенотипе

Независимое комбинирование
Когда изучается одновременное наследование двух признаков, то передача их потомкам зависит от локализации генов. Сначала мы рассмотрим сравнительно простой случай, когда гены локализованы на разных хромосомах, а затем более сложный, когда они локализованы на одной хромосоме.
Мендель изучал гибриды, полученные при скрещивании растений, различавшихся по двум четким признакам (т. е. проводил дигибридное скрещивание). Например, он скрещивал растения двух линий гороха; одни имели круглые и желтые семена, а другие — морщинистые и зеленые. Данные табл. 9-1 показывают, что признаки «круглый» и «желтый» — доминантны, а «морщинистый» и «зеленый» — рецессивны. Все растения первого поколения имели круглые желтые семена. Когда семена гибридов первого поколения были высеяны, а цветки беспрепятственно самоопылялись, то всего в поколении F2 было получено 556 растений. Из них 315 имели два доминантных признака, т. е. круглые желтые семена, 32 — два рецессивных, т. е. морщинистые зеленые семена. Все остальные растения дали семена, непохожие на имеющиеся у родителей: 101 — морщинистых желтых, 108 — круглых зеленых. Проявилась совершенно новая комбинация признаков.
Рис. 9-12 иллюстрирует опыты Менделя. Когда в скрещивании участвуют две пары доминантных и рецессивных аллелей, а каждая пара находится в разных хромосомах, фенотипы гибридов второго поколения распределяются в соотношении 9:3:3:1. При этом 9 — доля растений, имеющих оба доминантных признака, 3 — два возможных сочетания доминантных и рецессивных признаков и 1 — оба рецессивных. В приведенном примере одно родительское растение несло оба доминантных признака, а другое — оба рецессивных. Предположим, что каждое родительское растение имело один доминантный и один рецессивный признак. Будет ли результат таким же? Если вы не уверены в ответе, — попробуйте построить диаграмму возможных сочетаний, как сделали мы (рис. 9-12). Такие диаграммы называют решетками Пеннетта; они названы по имени английского генетика Реджиналда Пеннетта. Пеннетт был одним из ученых, в начале нашего столетия возродивших работу Менделя.
Рис. 9-12. Независимое расщепление. В одном из своих опытов Мендель скрестил растение с круглыми (RR) и желтыми (УУ) семенами с растением, которое имело морщинистые (rr) и зеленые (уу) семена. Растения первого поколения имели круглые и желтые семена. Однако в поколении как показано на схеме, рецессивные признаки вновь проявились. Более того, они проявились в новых комбинациях. В скрещиваниях, подобно этому включающих две пары аллелей в различных хромосомах, в поколении расчетное соотношение фенотипов 9:3:3:1. Соотношение генотипов: 1rrуу:2ггУу:1гrУУ:2Rrуу:1RRуу:4RrУу: 2RrУУ.2RRУу: 1RRУУ

На основании своих опытов Мендель сформулировал правило независимого комбинирования, которое гласит, что наследование пары факторов по одному признаку не зависит от одновременного наследования других признаков; эти факторы распределяются так, как будто другие факторы не существуют.
Сцепление
Когда Мендель проводил свои опыты, не было известно о существовании хромосом. Зная, что гены локализованы на хромосомах, легко представить, что, если разные гены, контролирующие некие признаки, локализованы на одной паре хромосом, они, как правило, не будут расщепляться независимо. В этом случае в поколении F2 не получится соотношения 9:3:3:1.
Либо по счастливой случайности, либо благодаря осторожному подбору признаков Мендель избежал явления сцепления, которое вряд ли смог бы объяснить. Горох огородный имеет семь пар хромосом; четыре главных признака, изученных Менделем, детерминируются генами, локализованными на четырех различных хромосомах, как показано в табл. 9-1. К тому же гены, локализованные на одной и той же хромосоме, так далеко отстоят друг от друга, что, за единственным исключением, распределяются независимо друг от друга. Только гены, определяющие форму боба и длину стебля, находятся в 4-й хромосоме так близко друг к другу, что Мендель должен был заметить эффект сцепления между этими двумя признаками. Однако он не сообщал об этом дигибридном скрещивании, и поэтому мы не знаем, проводил ли он его вообще.
Сцепление генов в 1905 г. открыл английский генетик Уильям Бэтсон, работая с душистым горошком (Lathyrus odoratus). Он и его сотрудники скрестили линию душистого горошка, гомозиготную по двум рецессивным признакам — красные лепестки и круглая пыльца, — с другой линией (похожей на дикую форму), имеющей пурпурные лепестки и продолговатую пыльцу. При этом в поколении F2 они обнаружили следующие признаки:
284: пурпурные + продолговатая,
21: пурпурные + круглая,
21: красные + продолговатая,
55: красные + круглая.
Если бы два гена (окраски лепестков и формы пыльцы) были локализованы на одной хромосоме, потомство бы получилось двух типов; если на двух разных хромосомах, то соотношение фенотипов составило бы 216:72:72:24 или 9:3:3:1. Ясно, что гены «родительского» типа наследовались вместе, как если бы они были сцеплены друг с другом.
Сцепление генов и связанное с ним явление кроссинговера стало понятным после работ Т. X. Моргана и его исследовательской группы в Колумбийском университете в начале 1900-х гг. Морган работал с плодовой мушкой дрозофилой. Drosophila — очень удобный для генетических исследований организм: ее популяции легко скрещивать и поддерживать; новые поколения мушек можно получать каждые две недели; каждая самка откладывает сразу сотни яиц; наиболее распространенный вид имеет только четыре пары хромосом. Самка плодовой мушки имеет две гомологичные половые хромосомы XX, тогда как самец имеет только одну X хромосому и гораздо более мелкую Y хромосому, в которой отсутствуют многие аллели X хромосомы.
В опытах Моргана была обнаружена необычная мушка с белыми глазами. Поскольку этот рецессивный признак всегда проявлялся у самцов, которые имеют две разные хромосомы — X и Y, Морган сделал вывод, что ген, определяющий цвет глаз, локализован у Drosophila на X хромосоме. Самки, несущие две X хромосомы, имели белые глаза только в том случае, когда были гомозиготны по аллелю белых глаз. Изучая наследование другого, «сцепленного с полом» признака — желтой окраски тела, Морган получил аналогичные результаты и решил, что ген данного признака тоже должен быть локализован на X хромосоме и у самца Drosophila представлен только одним аллелем. Поэтому его действие всегда должно проявляться, как если бы он принадлежал гаплоидному организму. Действительно, X хромосома самца Drosophila присутствует в гаплоидном состоянии.
Морган скрестил самцов мухи (XY), имеющих два рецессивных, сцепленных с полом признака, с самками «дикого типа» (XX). Как и ожидалось, все потомство первого поколения, и самки, и самцы, по фенотипу было «диким». В потомстве от анализирующего скрещивания между самкой первого поколения и двойным рецессивным самцом-родителем ожидалось расщепление между диким типом и белоглазым желтотелым. Однако часть потомства имела белые глаза и тело дикого типа, тогда как другая — глаза дикого типа и желтое тело.
Эти результаты объясняются тем, что гены «сцеплены» в одной хромосоме, но иногда происходит обмен между гомологичными хромосомами посредством кроссинговера. Теперь известно, что кроссинговер происходит в первой профазе мейоза (см. рис. 9-4). Чем больше расстояние между генами на хромосоме, тем больше вероятность кроссинговера. Чем ближе расположены гены друг к другу, тем больше вероятность их совместного распределения в мейозе и, таким образом, тем сильнее они сцеплены. Вычисляя частоту кроссинговера, можно построить генетические карты, которые дают приблизительное представление о порядке расположения генов на хромосоме.
Полигенное наследование
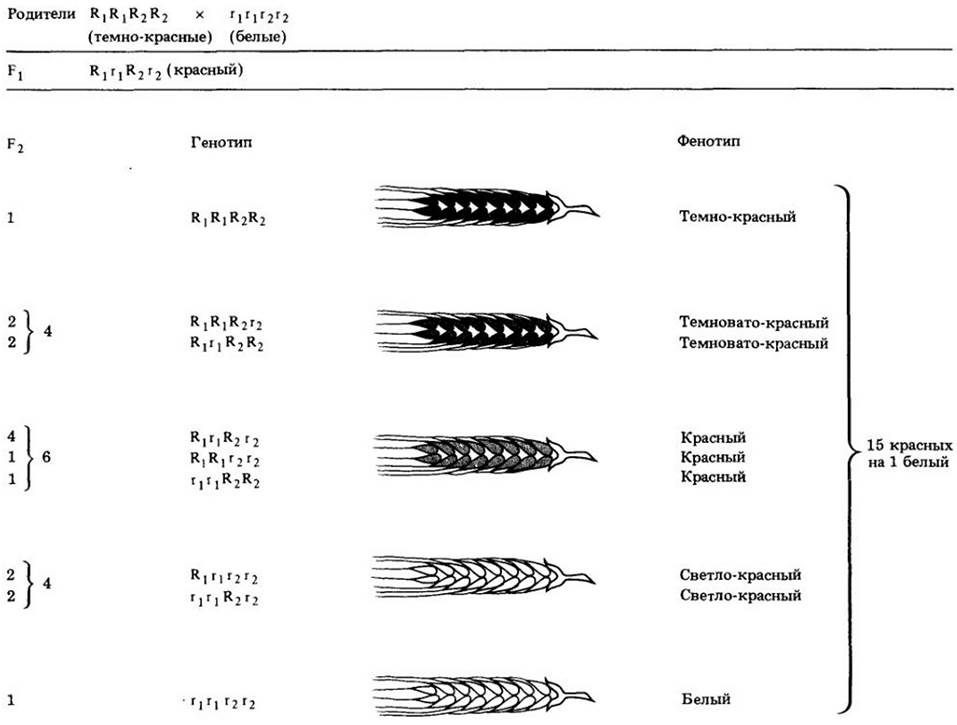
Первый опыт, показывающий, что в создании непрерывного характера изменчивости растений (в данном случае у пшеницы) могут участвовать несколько генов, был предпринят шведским ученым Г. Нильсоном-Эле. Табл. 9-2 иллюстрирует фенотипический эффект различных комбинаций двух генов (каждый представлен двумя аллелями), которые совместно контролируют интенсивность окраски зерна пшеницы. Благодаря подобным исследованиям мы начинаем понимать, как много генов могут взаимодействовать между собой и детерминировать сложные признаки растений и животных. Правила, впервые открытые Менделем, лишь через добрых сто лет смогли быть применены к анализу подобных признаков, над механизмом наследования которых ломали головы первые исследователи наследственности.
Таблица 9.2. Генетический контроль окраски зерен у пшеницы