СОВРЕМЕННАЯ БОТАНИКА - П. РЕЙВН - 1990
РАЗДЕЛ IV. РАЗНООБРАЗИЕ
ГЛАВА10. КЛАССИФИКАЦИЯ ЖИВЫХ СУЩЕСТВ
Половое размножение
ДНК у бактерий не дуплицирована, как у диплоидных эукариот; несмотря на это, и у прокариот известно несколько способов генетической рекомбинации. Как более подробно описывается в гл. 11, часть их генетического материала может быть перенесена из одной клетки в другую, но механизма, сравнимого с мейозом, посредством которого она затем регулярно передавалась бы по наследству вместе с генетическим материалом клетки-реципиента, здесь нет.
Единственным способом устойчивой передачи новой генетической информации у прокариот является встраивание полученного фрагмента в молекулу бактериальной ДНК. Этот способ не точен и с трудом воспроизводим.
Половое размножение с регулярным чередованием мейоза и сингамии имеет высокое селективное преимущество, поскольку служит основным механизмом возникновения и поддержания изменчивости природных популяций эукариот. Изменчивость, как мы узнаем из гл. 28, — важнейшая предпосылка приспособляемости живых организмов к окружающей среде в процессе эволюции.
Эволюция диплоидии
Первые эукариотические организмы были, вероятно, гаплоидными и размножались бесполым путем, однако с распространением среди них полового размножения началась эволюция диплоидии. По-видимому, она возникла первоначально при объединении двух гаплоидных клеток в диплоидную зиготу. Такое эволюционное событие скорее всего имело место неоднократно. Можно полагать, что зигота затем немедленно мейотически разделилась, восстановив гаплоидное состояние (рис. 10-10 и 10-11, А). У организмов с простым типом жизненного цикла зигота является единственной диплоидной клеткой.
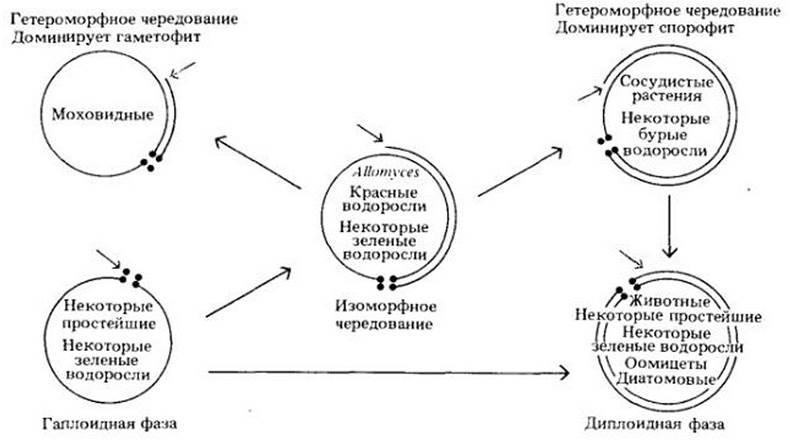
Рис. 10-10. Эволюция генетических систем. Каждый из кругов соответствует определенному типу жизненного цикла. Наиболее примитивные эукариоты были, несомненно, гаплоидными на протяжении большей части своего жизненного цикла (круг, очерченный одной линией, внизу слева). В этом случае мейоз (обозначен четырьмя жирными точками) происходит сразу после оплодотворения (показано цветной стрелкой). Другие жизненные циклы отличаются от гаплоидного моментом оплодотворения и протяженностью диплоидной фазы (обозначена внешней линией круга). Внутри кругов указаны некоторые современные группы организмов, характеризующиеся данным жизненным циклом
«Случайно» (а такой «случай» произошел в нескольких разных эволюционных линиях) некоторые из этих зигот поделились не мейотически, а митотически, дав в результате начало организму, состоящему из диплоидных клеток, вступающих в мейоз позднее. У животных подобный отсроченный мейоз ведет к образованию гамет — яйцеклеток и спермиев. Гаметы затем сливаются, что непосредственно восстанавливает диплоидное состояние (рис. 10-10 и 10-11, Б). Таким образом, у животных гаметы являются единственными гаплоидными клетками.
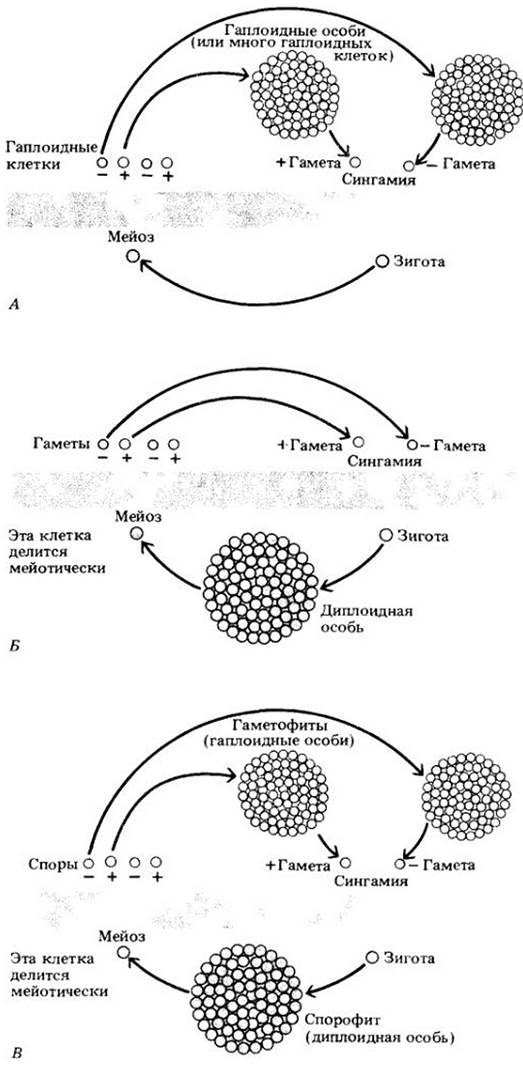
Рис. 10-11. Схемы основных типов жизненных циклов. Диплоидная фаза показана снизу, а гаплоидная сверху от широкой полосы. Четыре белые стрелки указывают на продукты мейоза; одиночная белая стрелка — на оплодотворенную яйцеклетку. A. При зиготическом мейозе зигота делится мейотически, образуя четыре гаплоидные клетки, делящиеся затем митотически с образованием новых гаплоидных клеток или многоклеточной особи, которая в конечном итоге дает гаметы за счет дифференцировки. Этот тип жизненного цикла встречается у хламидомонады и ряда других водорослей. Б. При гаметическом мейозе диплоидная особь образует путем мейоза гаплоидные гаметы, которые сливаются, давая диплоидную зиготу. В результате деления последней возникает новый диплоидный организм. Этот тип жизненного цикла характерен для большинства животных, некоторых протистов (оомицетов), а также для бурой водоросли Fucus. B. При спорическом мейозе спорофит, или диплоидная особь, в результате мейоза образует гаплоидные споры, не функционирующие как гаметы, а делящиеся митотически. При этом возникают многоклеточные гаплоидные организмы (гаметофиты), формирующие в конечном итоге гаметы, которые сливаются с образованием диплоидной зиготы. Та в свою очередь дает начало диплоидным особям. Этот тип жизненного цикла (чередование поколений) характерен для растений и многих водорослей. Сходный цикл наблюдается у хитридиомицета Allomyces и еще одного близкого к нему рода, а также у некоторых других групп протистов, не включенных в эту книгу
У растений мейоз приводит к образованию спор, а не гамет. Споры — это клетки, способные делиться митотически, давая многоклеточный гаплоидный организм; этим они отличаются от гамет, которые могут развиваться только после слияния друг с другом. Многоклеточные гаплоидные особи, чередующиеся с диплоидными формами, характерны для растений, некоторых бурых, красных и зеленых водорослей, двух близкородственных родов хитридиомицетов и других групп протистов, не рассматриваемых в этой книге. Такой процесс известен как чередование поколений (рис. 10- 10 и 10-11, В). Образующее гаметы гаплоидное поколение у растений называется гаметофитом, а диплоидное спорообразующее — спорофитом. Эта терминология используется также в случае водорослей, а иногда и применительно к другим группам.
У ряда водорослей (большинство красных, многих зеленых и отдельных бурых) диплоидная и гаплоидная формы внешне одинаковы. Такие типы жизненных циклов называют изоморфным чередованием поколений (рис. 10-10).
Имеются и другие жизненные циклы, в которых диплоидные и гаплоидные формы не идентичны. В ходе эволюции некоторых групп происходили мутации, которые проявлялись только в одном поколении, хотя соответствующие аллели были, очевидно, представлены и у диплоидных, и у гаплоидных особей. В жизненных циклах такого типа гаметофит и спорофит стали заметно отличаться друг от друга — возникло гетероморфное чередование поколений, характерное для растений и некоторых бурых водорослей (рис. 10-10).
У моховидных (мхов, печеночников, антоцеротовых) гаметофит доминирует; он независим в своем питании и обычно крупнее спорофита, который может быть структурно более сложным. У сосудистых растений, напротив, доминирует спорофит, который намного крупнее и сложнее гаметофита, почти у всех групп зависимого в своем питании от спорофита.
Как уже упоминалось раньше, диплоидия позволяет накапливать больше наследственной информации и гораздо тоньше регулировать проявление генетического материала организма в ходе его развития. Возможно, в этом причина того, что спорофит у сосудистых растений — крупное, сложное, независимое в отношении питания поколение. Одна из наиболее четких эволюционных тенденций у этой группы, преобладающей в большинстве наземных местообитаний, — усиление доминирования спорофита и подавление гаметофита. У цветковых растений женский гаметофит — микроскопическое тельце всего из семи клеток, а мужской — только из трех клеток. Оба они полностью зависят от спорофита в смысле питания.