СОВРЕМЕННАЯ БОТАНИКА - П. РЕЙВН - 1990
РАЗДЕЛ IV. РАЗНООБРАЗИЕ
ГЛАВА 13. ГРИБЫ
Основные группы грибов
Отдел Zygomycota
Большинство зигомицетов (всего описано около 600 видов) обитает в почве на разлагающихся остатках растений и животных, некоторые из них — паразиты растений, насекомых или небольших почвенных животных; ряд форм вызывает серьезные заболевания человека и домашних животных. Термин «зигомицеты» связан с основной особенностью этих грибов — образованием специфических половых структур, называемых зигоспорангиями, в результате слияния двух многоядерных гаметангиев (рис. 13-9). Внутри зигоспорангия гаметы (в данном случае просто ядра) сливаются в одно или более диплоидных ядер (зигот). Зигоспорангий иногда называют зигоспорой, однако мы предпочитаем этого не делать, так как иногда он содержит много таких ядер-зигот. Бесполое размножение с помощью спор, образуемых в одном или нескольких особых спорангиях на гифах, известно почти у всех зигомицетов. У большинства представителей этого отдела ценоцитные (многоядерные, не разделенные септами) мицелии, внутри которых часто заметно быстрое течение цитоплазмы.
Рис. 13-9. Плесень Rhizopus stolonifer поселяется на поверхности влажных субстратов в воздушной среде. А. Гаментагии — структуры, где образуются гаметы. Б. Зигоспорангий — покоящаяся стадия, связанная с половым размножением (темная масса в центре.). Содержит одну или несколько диплоидных зигот
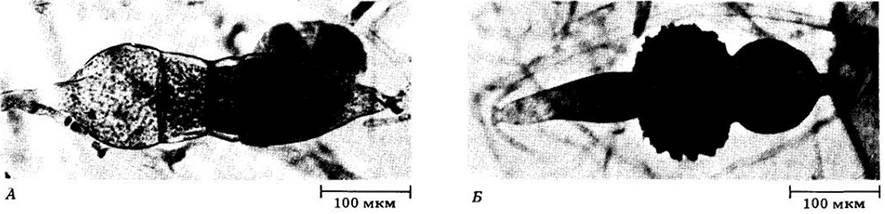
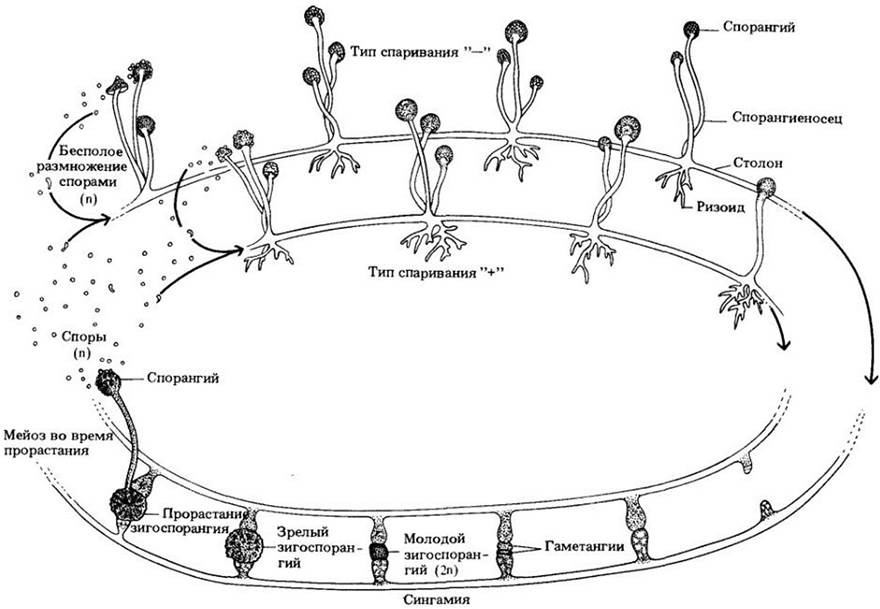
Один из наиболее распространенных зигомицетов — Rhizopus stolonifer, образующий похожую на вату черную плесень на поверхности влажной, богатой углеводами пищи или других аналогичных продуктов, оставленных на воздухе. Этот вид представляет серьезную угрозу для хранящихся овощей и фруктов, так как способен довольно быстро их испортить. Жизненный цикл R. stolonifer представлен на рис. 13-10. Мицелий образован тремя различными типами гаплоидных гиф. Основная их масса — быстро растущая ценоцитная. Эти гифы в основном находятся внутри субстрата, поглощая питательные вещества. От них отходят дуговидные гифы, называемые столонами, которые образуют ризоиды в местах контакта их верхушек с субстратом. Спорангии формируются на верхушках спорангиеносцев, в свою очередь возникающих в виде вертикальных ответвлений прямо над ризоидами. Спорангий сначала имеет вид вздутия с большим числом ядер внутри, затем отделяется от спорангиеносца септой, его цитоплазма фрагментируется по числу ядер, и вокруг каждой споры образуется клеточная оболочка. По мере созревания спорангии, чернеют, что обусловливает характерный цвет плесени. Каждая спора после рассеивания способна прорасти, образовав новый мицелий.
Рис. 13-10. У Rhizopus stolonifer в половом размножении участвуют только штаммы различных типов спаривания, обычно обозначаемых значками «-» или «+» (хотя такие штаммы внешне не отличаются, здесь они показаны двумя различными цветами). Когда два штамма тесно сближаются, образуются гормоны, вызывающие у верхушек гиф формирование особых выростов, которые, вступая в контакт, превращаются в гаметангии и отделяются от остального мицелия септами (см. рис. 13-9, А). Клеточные стенки между двумя соприкасающимися гаметангиями растворяются, и два многоядерных протопласта объединяются. Плюс- и минус-ядра сливаются попарно, и образуется молодой зигоспорангий с несколькими диплоидными ядрами. Затем он покрывается толстой шероховатой оболочкой черного цвета и остается в состоянии покоя, часто в течение нескольких месяцев. Мейоз происходит во время прорастания. Зигоспорангий вскрывается с образованием спорангия, сходного с возникающим при бесполом размножении, и жизненный цикл начинается заново
Одна из важнейших групп зигомицетов, включая род Glomus и несколько близких к нему форм, населяет корни растений, образуя микоризу, которая будет описана ниже.
Отдел Ascomycota
К аскомицетам относят около 30 тыс. видов, в том числе широко распространенных и экономически важных, в частности сизо-зеленые, красные и бурые плесени, вызывающие порчу пищевых продуктов, включая оранжево-розовый грибок Neurospora, сыгравший важную роль в развитии современной генетики. Аскомицеты вызывают ряд серьезных заболеваний растений, например, мучнистую росу (в основном на листьях), каштановую гниль (вызывается грибом Endothia parasitica, случайно завезенным в Америку из северного Китая), голландскую болезнь вяза (возбудитель Ceratocystis ulmi, гриб европейского происхождения). К этому же отделу относятся разнообразные дрожжи, съедобные сморчки и трюфели (рис. 13-11). В целом он изучен относительно слабо, и тысячи новых видов (в том числе, безусловно, экономически важных) ждут научного описания.
Рис. 13-11. Аскомицеты. А. Сморчок Morchella esculenta, один из наиболее популярных съедобных грибов. Грибники начинают искать его весной, когда дубовые листочки становятся «размером с мышиное ухо». В 1983 г. сморчки впервые получены в культуре, но в рыночных масштабах их пока еще не выращивают. Б. Sarcoscypha coccínea.
красивый гриб с открытым аскокарпом (апотецием). В. Высокоценимый съедобный аскокарп черного трюфеля (Tuber melanosporum). У трюфелей спорообразующие структуры формируются в почве и остаются в закрытом виде, высвобождая аскоспоры только после сгнивания аскокарпа или его разрушения роющими животными. Трюфели образуют микоризу (в основном с дубами). Их разыскивают с помощью специально тренированных собак или свиней, а недавно в небольших количествах начали разводить на продажу, инокулируя корни всходов соответствующего растения спорами гриба

Признаки аскомицетов
Аскомицеты, за исключением одноклеточных дрожжей, имеют нитевидное тело. Их гифы, как правило, септированы, т. е. разделены поперечными перегородками, имеющими поры, через которые могут проходить цитоплазма и ядра. Клетки вегетативного мицелия могут быть одно- или многоядерными. Некоторые виды аскомицетов гомоталломны (способны к самооплодотворению, т. е. половому процессу в пределах одного штамма), другие — гетероталломны и требуют для полового размножения присутствия плюс- и минус-штаммов.
Большинство аскомицетов размножается бесполо, образуя особые споры конидии (от греческого слова, означающего «мелкая пыль»), обычно многоядерные и возникающие из клеток (рис. 13-12), которые отшнуровываются на верхушках модифицированных гиф, называемых конидиеносцами.
Половое размножение аскомицетов всегда включает образование аска, или сумки, с гаплоидными аскоспорами.
Рис. 13-12. Конидии — типичные споры, образующиеся при бесполом размножении аскомицетов. На этой электронной микрофотографии представлены стадии образования конидий у гриба Nomuraea rileyi на гусенице совки. А. Фотография в сканирующем электронном микроскопе различных стадий развития конидий. Б. Фотография конидий в электронном просвечивающем микроскопе. У этого вида образование конидий происходит от верхушки к основанию

Эти образования отличают аскомицеты от всех прочих грибов (рис. 13-13). Аски формируются обычно внутри сложной структуры из плотно переплетенных гиф, аскокарпа, во многих случаях макроскопических размеров. Аскокарп может быть открытым в виде чаши (апотеций, рис. 13-11, Б), замкнутым и округленным (клейстотеций, рис. 13-13, Б), колбообразным, с мелкой порой для выхода аскоспоры (перитеций, рис. 13-13, В). Аски обычно развиваются на внутренней поверхности аскокарпа, и их слой называется гимением,или гимениальным (рис. 13-14).
Рис. 13-13. А. Электронная микрофотография асков Ascodesmus nigricans с созревающими аскоспорами. Б. Аскокарп Erysiphe aggregate с закрытыми асками и аскоспорами. Этот полностью замкнутый тип аскокарпа называется клей- стотецием. В. Аскокарп Chaetomium erraticum с закрытыми асками и аскоспорами. Заметна маленькая пора на верхушке. Такая разновидность аскокарпов с небольшим отверстием называется перитецием
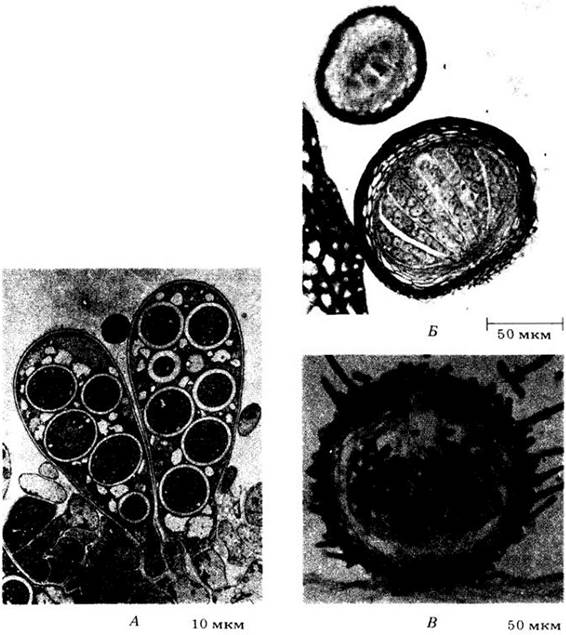
Рис. 13-14. Срез через гимениальный слой у сморчка (Morchella); видны аски с аскоспорами

Рис. 13-15. Типичный жизненный цикл аскомицета. Бесполое размножение связано с образованием особых, обычно многоядерных спор, называемых конидиями. Половой процесс включает формирование асков и аскоспор, возникающих в результате мейоза, который происходит в аске сразу же после кариогамии

На рис. 13-15 показан характерный для аскомицета жизненный цикл. Образование мицелия начинается с прорастания аскоспоры; вскоре он уже формирует конидиеносцы. В течение сезона роста образуется огромное число конидий; именно они отвечают за расселение гриба.
Аски возникают на том же мицелии, что и конидии; этому предшествует формирование многоядерных гаметангиев — антеридия и аскогона. Мужские ядра антеридия проходят в аскогон через вырост последнего, трихогину. Таким образом, имеет место плазмогамия — слияние двух протопластов.
Мужские ядра в пределах общей цитоплазмы находят себе парные, генетически отличные женские ядра, не сливаясь с ними. Затем из аскогона начинают расти и удлиняться аско- генные гифы, куда по мере их развития мигрируют пары ядер, причем в этих гифах и аскогоне проходят синхронные митотические деления. Клеточное деление в развивающихся аскогенных гифах ведет к образованию дикарионов, т. е. клеток с двумя гаплоидными ядрами.
Аск сначала закладывается на верхушке развивающейся аскогенной гифы, состоящей из дикарионов. При этом одна из ее двуядерных клеток разрастается в форме крючка, и два ядра в ней делятся таким образом, что их веретена располагаются примерно вдоль оси гифы параллельно друг другу. Два дочерних ядра в верхушке гифы сближаются, еще одно оказывается у «острия» крючка, а оставшееся лежит около септы в его основании. Затем образуются две перегородки, делящие клетку-крючок на три клетки, средняя из которых и превращается в аск. Именно в ней происходит кариогамия: два ядра сливаются, образуя диплоидное ядро (зиготу), единственное в жизненном цикле аскомицетов (за исключением парасексуального процесса). Вскоре после кариогамии «молодой» аск начинает удлиняться. Диплоидное ядро затем делится мейотически и обычно еще раз митотически. Если этот митоз происходит, зрелый аск содержит восемь спор, если митоз отсутствует — четыре. Образовавшиеся гаплоидные ядра с прилежащими участками цитоплазмы отделяются друг от друга, давая аскоспоры. У большинства аскомицетов зрелый аск разбухает и в конце концов лопается, выстреливая свои споры в воздух. Обычно они отлетают примерно на 2 см, но у некоторых видов дальность рассеивания достигает 30 см.
Одноклеточные аскомицеты: дрожжи
Дрожжи — в основном одноклеточные организмы, размножающиеся бесполо делением или почкованием (рис. 13- 16, A), реже с помощью спор. Половой процесс представляет собой слияние двух клеток или аскоспор с образованием зиготы. Зигота может давать диплоидные почки или функционировать как аск, делясь мейотически и образуя четыре гаплоидных ядра. Затем может происходить митоз. Внутри зиготы (т. e. теперь уже аска) вокруг ядер формируются оболочки, и возникают четыре или восемь аскоспор, высвобождающихся при разрушении стенки аска (рис. 13-16, Б). Аскоспоры либо бесполо почкуются, либо сливаются с другими клетками, чтобы повторить половой процесс.
Дрожжи играют важную роль в жизни человека из-за своей способности сбраживать углеводы, расщепляя глюкозу с образованием этилового спирта и СO2. В результате они используются в виноделии (источник этанола), хлебопечении (источник СО2) и пивоварении, где необходимы оба эти соединения. Многие полезные в хозяйстве штаммы дрожжей были получены путем отбора и скрещивания. Сейчас для их дальнейшего улучшения путем введения полезных генов из других организмов используются методы генной инженерии (см. гл. 30). В современном виноделии к относительно стерильному виноградному соку добавляют чистые штаммы дрожжей; раньше для получения вина применялись дикие штаммы, обитавшие на гроздьях (см. рис. 16-17). Иногда букет вина определяется непосредственно виноградом, но в большинстве случаев он зависит от использованного грибка. В пивоварении среда сначала стерилизуется нагреванием, а потом сбраживается чистой культурой дрожжей. Как правило, в производстве вина, сидра, сакэ и пива используются штаммы одного вида Saccharomyces cerevisiae, хотя находят применение и другие. Фактически только этот вид используется сейчас в хлебопечении (рис. 13-16,4). Некоторые дрожжи патогенны для человека, вызывая такие болезни, как молочница и криптококкоз.
Рис. 13-16. Дрожжи. А. Почкование клеток пекарских дрожжей (Saccharomyces cerevisiae). Б. Аски с аскоспорами у Schizosacchаrоmуces octosporus
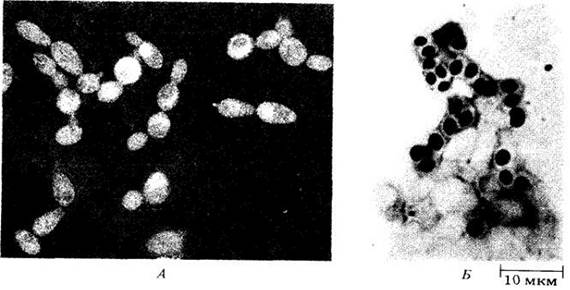
Рис. 13-17. Pénicillium и Aspergillus — два широко распространенных рода несовершенных грибов. А. Культура Pénicillium notatum — гриба, образующего пенициллин; видны различия в окраске, растущей и спороносящей частей мицелия. Б. Культура Aspergillus fumigatus — гриба, вызывающего заболевание дыхательных путей у человека. Хорошо заметен концентрический тип роста за счет последовательных «импульсов» спорообразования

Ряд видов, особенно Saccharomyces cerevisiae, — ценный объект генетических исследований. Как и бактерия Escherichia coli, этот вид стал излюбленным организмом для изучения метаболизма, генетики и развития, в данном случае у эукариотических клеток. Недавно ученые сумели даже синтезировать функциональную дрожжевую микрохромосому, передаваемую в ходе последовательных митотических циклов; такого еще не удавалось в случае других эукариот. Детальное изучение дрожжей при легкости экспериментирования с их генетическим материалом, несомненно, намного повысит значение этих организмов в промышленности будущего (см. рис. 13-3).
Большинство дрожжей — аскомицеты, однако несколько их родов относят к базидиомицетам, например, Cryptococcus, вызывающий заболевание криптококкоз, а также некоторые непатогенные виды Candida. Упрощенная и обычно одноклеточная структура дрожжей — очевидно, результат эволюции более сложных мицелярных грибов. Редукция исходного строения этих организмов зашла так далеко, что трудно определить родственные связи дрожжей с другими группами. Однако, насколько известно, все они произошли от многоклеточных предков, причем одноклеточные формы возникали у грибов не один раз даже среди аскомицетов. К дрожжам относят 39 родов и около 350 видов. Они распространены в самых разнообразных сухопутных и водных местообитаниях, где находят подходящий источник углерода.
Fungi imperfecti (несовершенные грибы)
Несовершенные грибы, или дейтеромицеты, включают около 25 000 описанных видов, для которых неизвестен половой процесс (рис. 13-17, 13-18), что объясняется их недостаточной изученностью или утратой полового размножения в процессе эволюции. Большинство таких «несовершенных» грибов — по сути дела аскомицеты, образующие только конидии, но некоторые по характерным септам и пряжкам можно отнести к базидиомицетам (см. рис. 13-29). Пара сексуальные циклы, широко распространенные среди несовершенных грибов, поддерживая изменчивость, вероятно, частично компенсируют отсутствие у них полового процесса. В целом классификация здесь основана на способе образования конидий (рис. 13-12, 13-18). Несовершенные грибы — искусственная группа, объединяющая разнообразные организмы и сохраняемая в основном для удобства определения форм. Ее можно рассматривать как класс, не эквивалентный по своей сути другим классам грибов.
Рис. 13-18. Конидиеносцы несовершенных грибов используются в их классификации: ветвистые у Penicillium (А) и собранные в плотные пучки у Aspergillus (Б)

Многие из несовершенных грибов имеют большое экономическое значение. Например, некоторые представители рода Pénicillium придают определенным сортам сыра вид, запах и вкус, высоко ценимые гурманами. Одна из таких плесений — P. roquefortii — впервые найдена в пещерах вблизи французской деревни Рокфор. Согласно легенде, крестьянский мальчик забыл свой завтрак — кусочек обычного свежего сыра — в одной из пещер, а вернувшись через несколько недель, нашел на его месте острый ароматный сыр с цветными разводами. Название «Рокфор» могут иметь только сыры, изготовленные в этом районе Франции. Другой вид этого рода — P. camembertii — придает специфические свойства сыру «Камамбер». На Востоке, сбраживая соевые бобы с Aspergillus oryzae, получают соевое тесто (мисо), сбраживая их со смесью A. oryzae и A. soyae — соевый соус (сею). При этом молочнокислые бактерии и дрожжи используются для придания окончательных свойств продукту (соевый творог, тофу, и темпе, сходная богатая белком пища в странах тропической Азии, приготовляются путем сбраживания соевых бобов с зигомицетами родов Мисоr и Rhizopus соответственно). А. oryzae важен также для первых этапов производства сакэ — традиционного в Японии алкогольного напитка; дрожжи S. cerevisiae используются на следующих стадиях этого процесса. В промышленности лимонную кислоту в больших количествах получают из колоний Aspergillus, выращенных в сильно кислой среде. Сейчас в Европе и США исследуют проблему обогащения белком кормов для скота путем сбраживания их с А. oryzae.
Антибиотики — особые вещества, синтезируемые несовершенными грибами, подавляют рост других организмов (например, бактерий) и применяются при лечении многих заболеваний человека. Первый антибиотик был открыт в 1928 г. Александером Флемингом, заметившим, что Pénicillium, попавший в растущую на питательной агаровой среде культуру стафилококка, полностью подавил рост этой бактерии. Десять лет спустя Говард Флори и его коллеги из Оксфордского университета выделили чистый пенициллин, а позднее в США развернули широкомасштабное производство этого лекарственного препарата. Во время второй мировой войны спрос на него был так велик, что производство увеличилось с нескольких миллионов единиц в 1942 г. до 700 млрд. единиц в 1945 г. Пенициллин эффективен при лечении широкого спектра заболеваний, вызываемых грамположительными бактериями, включая пневмонию, скарлатину, сифилис, гонорею, дифтерит, ревматизм и многие др. Многие антибиотики, используемые как лекарства, несомненно, играют и важную экологическую роль в природе, позволяя синтезирующим их организмам побеждать в конкуренции с другими существами.
Далеко не все соединения, образуемые несовершенными грибами, полезны для человека. Например, афлатоксины, группа очень близких между собой вторичных метаболитов видов Aspergillus flavus и A. parasiticus — высокотоксичны и канцерогенны. Иногда эти грибы поселяются на пище при ее хранении, поэтому они постоянно находятся в центре внимания здравоохранения. Другая группа грибных ядов — трихотецены, образуемые некоторыми родами несовершенных грибов. Они подавляют синтез белка у эукариот.
Одна из групп несовершенных грибов — дерматофиты (от греческого слова dermatos — кожа и phyton — растение) — включает возбудителей стригущего лишая, эпидермофитии стопы и других кожных заболеваний. Эти болезни особенно распространены в тропиках. Патогенные стадии здесь соответствуют бесполому размножению, но большинство таких грибов сейчас отнесено к видам аскомицетов, хотя классификация продолжает основываться на их болезнетворных формах. Ступни поражаются в теплых и влажных условиях, но болезнь, как правило, быстро проходит, если человек меняет закрытые ботинки или туфли на сандалии и держит ноги сухими. Хотя обычно на 1 см2 подошвы стопы обитает около 2 млн. бактерий, конкурирующих с дерматофитами за питательные вещества, грибы побеждают в борьбе с ними, так как прорастают прямо в клетки эпидермиса и отравляют бактерий выделяемыми антибиотиками. Грибы гораздо активнее используют кератин, прочный волокнистый белок, в который превращается содержимое эпидермальных клеток перед тем, как они слущиваются. Однако среди бактерий могут развиваться устойчивые штаммы, способные при активном размножении вызвать тяжелую инфекцию.
Во время второй мировой войны из южной части Тихого океана в связи с кожными заболеваниями эвакуировалось больше солдат, чем из-за ранений. Многие грибы — возбудители специфических инфекций; например, Candida albicans, дрожжеподобный при определенных условиях вид, вызывает молочницу и другие поражения слизистых оболочек. Споры грибов постоянно вдыхаются людьми, и некоторые из них являются причиной внутренних болезней, иногда очень тяжелых и даже смертельных, особенно если затрагиваются легкие.
В последние годы микозы распространились у людей значительно шире, чем прежде. Некоторые соединения, в частности циклоспорин (см. с. 184), впервые полученный из несовершенного гриба Tolypocladium inflatum, сейчас обычно вводятся пациентам при трансплантациях для подавления нормальных иммунных реакций, грозящих отторжением пересаженных органов или тканей. Однако при этом усиливается восприимчивость к грибковым и другим заболеваниям. Некоторые химические вещества, например, применяемые в случае острой лейкемии, также снижают устойчивость организма к грибковой инфекции. В результате внимание к проблеме ее профилактики и лечения постоянно возрастает.
Лишайники
Лишайники — это симбиотические ассоциации между аскомицетами и некоторыми родами зеленых водорослей или цианобактерий1. Их автотрофные компоненты, поставляющие питательные вещества, защищены от экстремальных воздействий среды грибными симбионтами. В результате таких симбиотических взаимоотношений лишайники могут населять самые суровые для живого местообитания (рис. 13- 19). В их составе обнаружено около 20 000 видов грибов различной морфологии, заметно отличающихся от примерно 30 000 видов других аскомицетов, и около 26 родов фотосинтезирующих организмов. Наиболее часто встречаются зеленые водоросли Trebouxia, Pseudotrebouxia, Trentepohlia и цианобактерия Nostoc, являющиеся автотрофными компонентами у примерно 90% всех видов лишайников.
1 Около дюжины видов базидиомицетов также сосуществуют с водорослями, но они тесно связаны со свободноживущими базидиомицетами и сильно отличаются от других грибов, образующих лишайники.
Лишайники чрезвычайно широко распространены в природе; они обитают по всему миру — от аридных пустынь до Арктики: на голой почве, стволах деревьев, раскаляемых солнцем скалах, на заборах и обдуваемых ветрами альпийских пиках (рис. 13-20, 13-21, и 13-22). Некоторые из них настолько малы, что почти не видны невооруженным глазом, другие, например, олений «мох», покрывают огромные площади ковром глубиной по щиколотку. Вид Verrucaria serpuloides постоянно погружен в морскую воду. Лишайники часто первыми поселяются на обнажающихся каменистых субстратах. В Антарктике их обитает более 350 видов (рис. 13-19) — и только два вида сосудистых растений; семь видов лишайников сейчас встречается на 86° ю. ш., т. е. около самого Южного полюса! Их почвообразовательная деятельность на голых скалах — первая стадия биологической сукцессии (см. гл. 31) в этих районах. Виды лишайников, у которых автотрофный симбионт представлен цианобактерией, имеют особое значение, так как способствуют связыванию N2 в почве. Они являются основным источником доступного азота во многих районах.
Рис. 13-19. В этом сухом, на первый взгляд безжизненном районе Антарктиды (А) лишайники (Б) живут сразу же под поверхностью песчаника. На изломе породы видны различно окрашенные полосы, соответствующие особым биологическим зонам. Черная и белые зоны образованы лишайниками, в то время как нижняя зеленая зона —
одноклеточными зелеными водорослями. Температура в этой части Антарктиды поднимается примерно до 0°С летом, а зимой может падать до — 60°С

Окраска лишайников — от белой до черной через различные оттенки красного, оранжевого, коричневого, желтого и зеленого. Они содержат много необычных химических соединений. Многие лишайники используются во всем мире как источники красителей. Например, раньше характерный цвет харрисского твида получали, окрашивая шерсть веществом именно такого происхождения. Многие лишайники применяются в медицине и парфюмерии, а у некоторых народностей употребляются в пищу.
Рис. 13-20. Коркообразный (накипной) лишайник на голой поверхности скалы в центральной Калифорнии
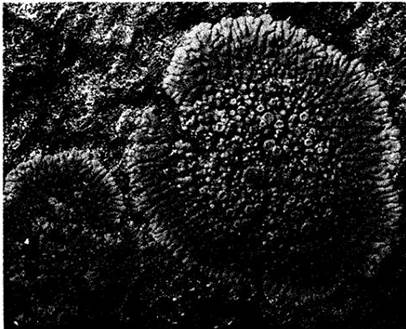
Рис. 13-21. A. Parmeliaperforata — листоватый лишайник, покрывающий гнездо колибри на мертвой ветке дерева в Миссисипи. Б. Usnea — лишайник, который часто в массе свешивается с ветвей деревьев. Удивительно похож на этот лишайник и занимает ту же экологическую нишу «бородатый мох» (Tillandsia usneoides), распространенный в южных штатах США, являющийся на самом деле настоящим цветковым растением из семейства броме лиевых

Рис. 13-22. Несколько кустистых лишайников. A. Teloschistes chysophthalmus. Б. Cladonia cristatella высотой 1 — 2 см. В. Cladonia subtenium так называемый «олений мох», являющийся на самом деле лишайником. Виды этой группы, обильно представленные в Арктике, концентрировали радиоактивные вещества, выпадавшие на землю после атмосферных ядерных испытаний. Северные олени, питаясь ими, накапливали радиоизотопы еще сильнее, передавая их человеку и животным, поедающим самих оленей, мясо или продукты, получаемые от них, особенно молоко и сыр
Лишайники уже давно исследуются биологами в связи с интересной природой взаимоотношений между их грибными и водорослевым симбионтами. По-видимому, гриб определяет форму всего организма, однако недавно выяснилось, что один и тот же гриб с разными водорослями может давать морфологически весьма различные структуры, которые традиционно относили к различным родам. Водоросли или цианобактерии в составе лишайников представлены видами, встречающимися и в свободноживущем состоянии, в то время как лишайниковые грибы, как правило, существуют только в симбиозе с ними.
Некоторые лишайники образуют особые структуры, соредии, состоящие из грибных гиф и водорослей или цианобактерий (рис. 13-23). Рассеивание этих соредий способствует заселению лишайниками новых местообитаний. Лишайниковые грибы часто образуют аскокарпы, аналогичные наблюдаемым у свободноживущих видов и отличающиеся только продолжительным существованием и образованием спор в течение нескольких лет.
Рис. 13-23. А. Поперечный срез через лишайник Lobaria verrucosa. В простейшем случае на таком срезе виден корковый слой из грибных гиф, оплетающих клетки водорослей. В более сложном случае гифы и водоросли образуют талломы с определенной формой роста и характерным внутренним строением. В лишайнике, представленном на рисунке, четко заметны четыре разных слоя: верхняя защитная кора из сильно желатинизированных гиф; водорослевый слой из клеток водорослей и рыхло переплетенных тонкостенных гиф; сердцевина — толстый слой из рыхло лежащих, бесцветных, слабо желатинизированных гиф (этот слой, составляющий около 2/3 толщины таллома, вероятно, является запасающим и образован гифами с крупными клетками); нижняя кора, которая тоньше верхней и несет особые выросты для прикрепления к субстрату. Соредии — фрагменты лишайника, содержащие клетки водорослей и грибные гифы, — служат для его расселения в новые местообитания. Б. Фотография в сканирующем электронном микроскопе среза Cladonia cristatella (см. pue. 13-22, Б и 13-25). Показана верхняя кора, водорослевый слой и сердцевина. У этого вида нижней коры нет

Биология лишайников
Как могут лишайники выживать в условиях среды, столь неблагоприятных для любой другой формы жизни? Одно время полагали, что секрет их успеха связан с защитой водоросли или цианобактерии от высыхания грибным симбионтом. Однако на самом деле одним из важнейших факторов их выживания является, по-видимому, именно способность очень быстро высыхать. Лишайники часто пребывают в почти обезвоженном состоянии, когда их влажность составляет от 2 до 10% сухой массы. При высыхании фотосинтез прекращается; погрузившись в такой «анабиоз», некоторые виды могут выдерживать сильное солнечное облучение, сильное нагревание и жестокие холода. Прекращение фотосинтеза в значительной степени обусловлено тем, что верхняя кора лишайника, высыхая, становится толще и непрозрачнее, преграждая путь солнечной энергии. Влажный лишайник разрушается ярким светом и экстремальными температурами, не влияющими на него в сухом виде.
Смачиваясь дождем, лишайники очень быстро поглощают воду (в 3 — 35 раз больше собственной массы). Если сухой хрупкий лишайник погрузить в воду, он станет через несколько минут мягким и гибким. Так действует простой физический процесс впитывания жидкости, аналогичный наблюдаемому в случае промокательной бумаги.
Максимально жизнеспособен лишайник, если судить по фотосинтезу, после того как, пропитавшись водой, начинает подсыхать. Фотосинтез идет наиболее интенсивно при влажности 65— 90% предельной влагоемкости; ниже этого уровня, если потеря воды продолжается, скорость фотосинтеза падает. Во многих местообитаниях влажность лишайников сильно колеблется в течение суток, и у многих из них фотосинтез возможен в течение лишь нескольких часов, обычно рано утром после смачивания туманом или росой. Следствие этого — очень низкая скорость роста лишайников; их радиус увеличивается на 0,1 — 10 мм в год. Если, исходя из размеров, рассчитать возраст некоторых зрелых лишайников, то получится 4500 и более лет. Наиболее активно растут и развиваются эти организмы на морских побережьях или в горах с обильными туманами.
По-видимому, лишайники поглощают некоторые минеральные вещества из своего субстрата (это подтверждается тем, что ряд их видов обитает только на особых горных породах, почвах или древесных стволах), но большая часть элементов улавливается ими из воздуха и дождевой воды. Поглощение элементов из дождевой воды идет очень быстро и сопровождается их концентрированием. Лишайники играют важную роль в функционировании экосистем, однако они особо чувствительны к токсичным веществам, так как не могут выделять в среду впитанные элементы. Яды вызывают разрушение хлорофилла в клетках водорослей или цианобактерий. Рост лишайников — очень чувствительный индикатор наличия в воздухе вредных примесей, и они все больше используются в мониторинге загрязнения атмосферы, особенно вокруг крупных городов. Наиболее резко лишайники реагируют на сернистый газ, который, возможно, быстро разрушает и без того небольшое количество их хлорофилла.
Как «здоровье» лишайника, так и их химический состав используются для индикации «качества» местообитания. На этой основе можно следить за присутствием тяжелых металлов или других загрязнителей вокруг промышленных центров. Многие лишайники способны связывать тяжелые металлы на наружной стороне своих клеток, тем самым предотвращая их поражение.
Когда ядерные испытания проводились в атмосфере, лишайники использовались для контроля за выпадением радиоактивных осадков. Сейчас полагают, что их применение целесообразно в мониторинге радиоактивного загрязнения, возможного при разрушении спутников, особенно когда это происходит в отдаленных районах, которые трудно обследовать другими средствами.
Природа взаимоотношений между грибами и фотосинтезирующими организмами
Каковы взаимоотношения между двумя компонентами лишайника? Ясно, что гриб получает органический углерод от водоросли или цианобактерии, поскольку лишайник ведет себя как типичный автотрофный организм, зависящий только от света, воздуха и минеральных веществ. Действительно, транспорт органического углерода из водоросли или цианобактерии в гриб был продемонстрирован с помощью меченного 14С углекислого газа. У лишайников, содержащих цианобактерии типа Nostoc, важны также связывание автотрофом азота и передача его гетеротрофному компоненту. В лишайнике грибные гифы образуют густую сеть вокруг лежащих внутри клеток. Широко распространенные здесь гаустории (специализированные гифы паразитических грибов) проникают в фотосинтезирующие клетки (рис. 13-24); другие особые органы, апрессории, плотно прилегают к поверхности этих клеток и внедряются в них своими выступами. Взаимодействие между грибом и автотрофным организмом сильно влияет на метаболизм последнего. Например, зеленые водоросли выделяют большие количества спиртов D-сорбитола и D-рибитола только под влиянием гриба в составе лишайника.
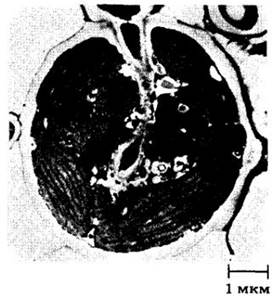
Рис. 13-24. Гаустория грибного компонента лишайника Strigula elegans проникает в своего партнера, зеленую водоросль Cephaleuros virescens. В ней хорошо заметны плотно упакованные хлоропласты
В некоторых случаях удается разделить фотосинтезирующий и грибной компоненты лишайника и вырастить их чистые культуры. При этом гриб образует компактные колонии, совершенно непохожие на симбиотический организм. Для его роста необходим широкий набор сложных углеводов, а спорообразующих структур, как правило, не образуется. И напротив, выделенные из лишайника водоросли или цианобактерии быстрее растут в свободном состоянии. Таким образом, можно предполагать, что партнерство в лишайнике скорее является контролируемым паразитизмом гриба на автотрофе, чем их симбиозом. Тем не менее лишайник не простая сумма компонентов, а вполне самостоятельный организм. При совместном выращивании в культуре гриб, по-видимому, первым берет под контроль фотосинтезирующего партнера, что приводит к появлению характерных морфологических черт зрелого лишайника (рис. 13- 25).
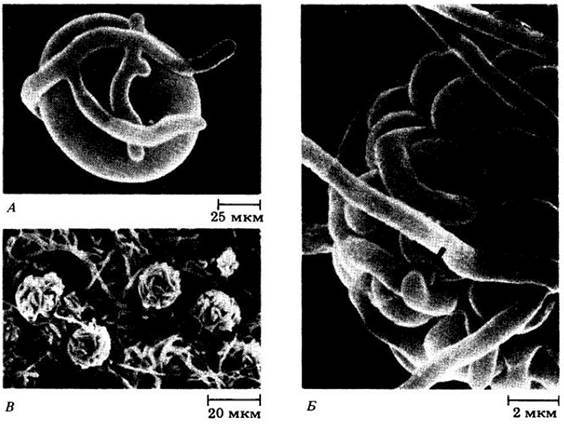
Рис. 13-25. Фотография в сканирующей электронном микроскопе ранних стадий взаимодействия между грибным и водорослевым компонентом лишайника Сladonia cristella в лабораторной культуре. Фотосинтезирующий партнер в этом лишайнике (см. рис. 13-23, Б) — зеленая водоросль Trebouxia. А. Клетка водоросли, окруженная грибными гифами. Б. Проникновение грибных гаусториев в клетки водорослей. В. Смешанная группа гиф и водорослевых клеток, развивающаяся в зрелый лишайник
Отдел Basidiomycota
Наиболее привычные нам грибы принадлежат к этому большому отделу, включающему около 25 000 описанных видов, в том числе съедобные и ядовитые шляпочные грибы, веселки, дождевики, трутовики и две важные группы патогенов растений — ржавчинные и головневые грибы (рис. 13-26). Отличительный признак базидиомицетов — образование базидиоспор на наружной стороне булавовидной спороносной структуры, базидии (рис. 13-27). В природе они размножаются в основном такими спорами. Хотя крупные базидиомицеты наиболее изученная группа грибов, неописанных видов и здесь гораздо больше, чем известных.
Рис. 13-26. Головня кукурузы — распространенная болезнь растений, при которой гриб Ustilago mayáis образует черные пылевидные массы спор на початках кукурузы. Головневые грибы относятся к классу телиомицетов из отдела Basidiomycota
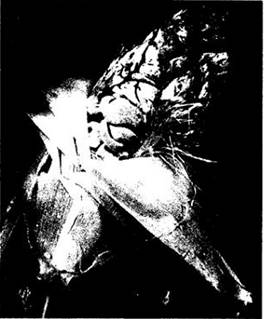
Рис. 13-27. Фотография в сканирующем электронном микроскопе верхушки базидии вида Aleurodiscus amorphus с удлиненными бородавчатыми базидиоспорами, прикрепленными к четырем стеригмам

Мицелий базидиомицетов всегда септированный, и в течение жизненного цикла большинство видов проходит две различные фазы — первичную и вторичную. Прорастая, базидиоспоры образуют первичный мицелий. Сначала он может быть многоядерным, но вскоре образуются септы, разделяющие его на монокариотические (одноядерные) клетки. Обычно вторичный мицелий возникает при слиянии первичных гиф, относящихся к разным типам спаривания (в этом случае он гетерокариотический), но иногда септы после ядерного деления не формируются, и он является гомокариотическим. В любом случае результатом будет дикариотический (с двуядерными клетками) мицелий, так как кариогамия следует за плазмогамией не сразу.
Септы вторичного мицелия базидиомицетов перфорированы, однако поры окаймлены толстыми боченковидными структурами (исключение — ржавчинные и головневые грибы) (рис. 13-28). Ядра через такие поры проходить не могут.
Рис. 13-28. Электронная микрофотография характерной септы вторичного мицелия базидиомицета Laetisaria arvalis, обычного гриба, портящего древесину. Как свойственно для базидиомицетов, септа перфорирована порой

В гетерокариотических вторичных мицелиях, где миграция ядер, по-видимому, все же происходит, поры в септах простые и широкие, аналогичные наблюдаемым у аскомицетов (см. рис. 13-7).
Апикальные (верхушечные) клетки вторичного мицелия обычно делятся с образованием пряжковидного мостика (рис. 13-29). Эти мостики (пряжки), обеспечивающие распределение ядер каждого типа между дочерними клетками, — характернейший признак базидиомицетов.
Рис. 13-29. А. У базидиомицетов дикариотические гифы отличаются наличием соединений-пряжек в обход септ; пряжки образуются при делении клетки и, по-видимому, обеспечивают правильное распределение двух генетически различных типов ядер в базидиокарпе. Б. Пряжки и септа характерного строения в гифе базидиомицета

Именно вторичный мицелий образует базидиокарпы — мясистые спорообразующие тела, как у шляпочных грибов или дождевиков. Их формирование иногда требует света; по мере его вторичный мицелий дифференцируется на специализированные гифы, выполняющие в базидиокарпах различные функции. Такой мицелий иногда называется третичным.
Базидиомицеты делятся на три класса: гименомицеты (Нymenomycetes), гастеромицеты (Gesteromycetes) и телиомицеты (Теliomycetes).
Класс Hymenomycetes
К этому классу относятся съедобные и ядовитые шляпочные грибы, рогатиковые грибы и трутовики (рис. 13-4, 13-30). Сюда включают все формы, образующие базидиоспоры на базидиокарпе (у гастеромицетов — внутри базидиокарпа). Базидии гименомицетов всегда формируются на гимении (откуда и название данной группы), а не из изолированных клеток, как у телиомицетов. Базидиокарпы гимено- и гастеромицетов аналогичны аскокарпам аскомицетов.
Рис. 13-30. Базидиомицеты из класса гименомицетов. А. Мухомор Amanita muscarina на разных стадиях развития; можно видеть спороносные пластинки. Типичный признак данного рода, в котором много ядовитых видов, — кольцеобразный «воротничок» на ножке и чашеобразная структура в ее основании. Б. Масленок Suillus bovinas. Спороносная часть базидиокарпа представлена губчатой структурой с порами. В. Трутовик Ganoderma tsugae. Такие грибы вызывают большинство случаев порчи древесины. Г. Съедобный рогатиковый гриб Hercium corralloides. Гимений, внешний спороносный слой с базидиями, образуется по всей поверхности базидиокарпа

Некоторые гименомицеты имеют булавообразные несептированные (т. е. не разделенные перегородками) базидии, каждая из которых обычно несет четыре базидиоспоры на крошечных выступах, стеригмах (рис. 13-27, 13-31). У других студенистых с виду, так называемых дрожалковых грибов (рис. 13-32) базидии септированные, как у телиомицетов.
Рис. 13-31. Навозник Сорrinus, обычный шляпочный гриб. А. Срез через спороносные пластинки. Относительно темные их края соответствуют г имению. Б. Срез через гимениальный слой с развивающимися базидиями и базидиоспорами. В. Почти зрелые базидиоспоры, прикрепленные к базидии стеригмами

Рис. 13-32. Дрожалковый гриб на мертвом пне в амазонском тропическом лесу (Бразилия). Дрожалковые — единственная группа гименомицетов с септированными балидиями
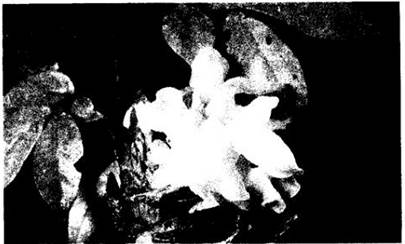
Шляпочный «гриб» в бытовом понимании — это базидиокарп, состоящий обычно из шляпки на ножке (пеньке). Масса гиф в базидиокарпе, как правило, образует различные ткани и слои. На ранних стадиях он может быть покрыт пленкой, которая разрывается по мере роста шляпочного гриба. У некоторых родов остатки ее видны на верхней поверхности шляпки или в основании ножки, где образуется чашевидное покрывальце (вольва). У многих гименомицетов нижняя поверхность шляпки состоит из радиально расходящихся пластинок,несущих гимении (рис. 13-31), у других он расположен иначе; например, у рогатиковых покрывает зубцы или пальцевидные выросты базидиокарпа (рис. 13- 30, Г). У трутовиков, белых грибов, подберезовиков и т. д. гимений выстилает многочисленные поры на нижней стороне шляпки (рис. 13-30, Б).
В относительно однородных местообитаниях, например, на полянах или лугах, мицелий распространяется под землей вниз и в стороны, образуя на поверхности кольцо базидиокарпов, достигающее иногда до 30 м в диаметре. Дело в том, что на открытом пространстве он равномерно растет во всех направлениях и отмирает в центре, а спороносные структуры формируются только по его наружному краю, где развитие гриба идет более активно, так как именно в этой зоне почва наиболее богата питательными веществами. В результате возникают кольца базидиокариов, становящиеся по мере роста мицелия все шире. В народе их называют «ведьмиными кольцами» (см. рис. 13-36).
Пластинчатые грибы — самые известные среди базидиомицетов. Особенно обилен Agaricus campestris — шампиньон обыкновенный. Родственный ему Agaricus bisporus (шампиньон двуспоровый) — один из немногих шляпочных грибов, которые можно разводить для продажи. Сейчас его выращивают более чем в 70 странах, и суммарная прибыль с урожая достигает 14 млрд, долларов в год. Вместе с грибом из Восточной Азии сиитаке (Lentinus edodes) он дает около 86% мирового производства грибов. Разводят и другие виды, но большинство съедобных гименомицетов в значительных количествах собирается в природе и регулярно поступает в продажу. Не все шляпочные грибы съедобны; например, много ядовитых форм среди группы пластинчатых. Так, род Amanita включает и самые опасные и некоторые съедобные виды. Даже крошечный съеденный кусочек бледной поганки (Amanita phalloides) может привести к смерти человека. Некоторые базидиомицеты содержат вещества, вызывающие у людей галлюцинации (рис. 13-34).
Типичный жизненный цикл шляпочного гриба представлен на рис. 13-33. Базидии образуются на гимении, выстилающем пластинки, как верхушечные клетки дикариотических гиф. Вскоре молодая базидия увеличивается в размерах и происходит кариогамия. Почти сразу же за этим следует мейоз каждого диплоидного ядра с образованием четырех гаплоидных ядер. Эти четыре ядра по одному мигрируют в стеригмы, которые расширяются на верхушке, формируя одноядерные базидиоспоры. Репродуктивные возможности одного шляпочного гриба громадны — один базидиокарп рассеивает миллиарды спор.
Рис. 13-33. Жизненный цикл шляпочного гриба (класс гименомицеты). Первичные мицелии образуются из базидиоспор. Вторичный дикариотический мицелий возникает из первичных. В некоторых случаях он образуется в результате слияния гиф различного типа спаривания и является гетерокариотическим. Вторичный мицелий делится и дифференцируется, давая третичный мицелий, из которого строится базидиокарп
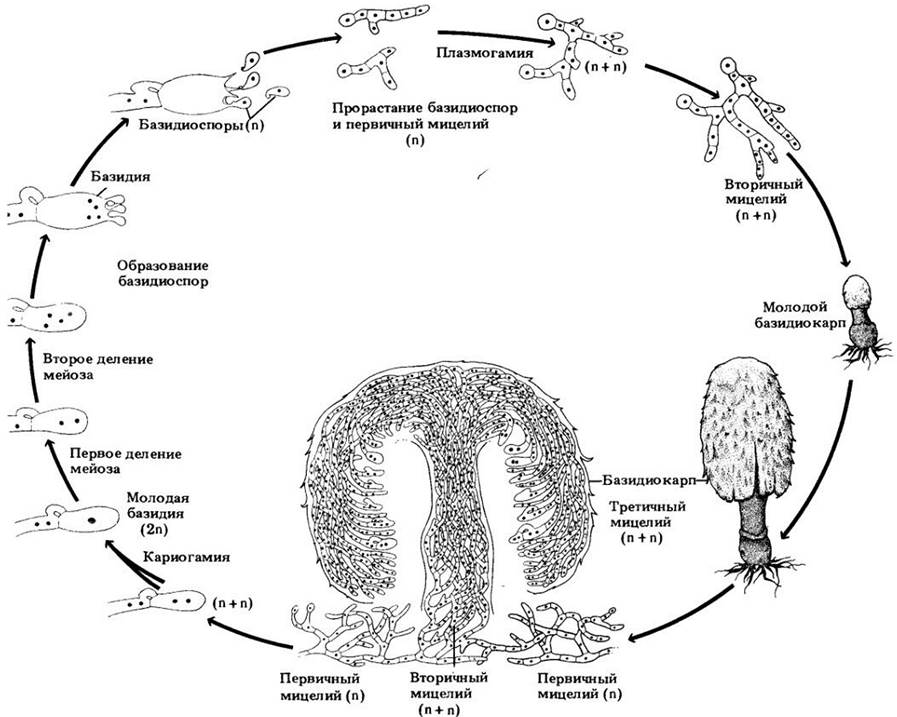
Рис. 13-34. Шляпочные грибы выполняют особые функции в религиозных церемониях некоторых индейских племен южной Мексики и Центральной Америки. При этом индейцы едят определенные базидиомицеты, способные вызывать галлюцинации. Один из наиболее известных таких грибов — Psilocybe mexicana — показан здесь растущим на пастбище близ Уаутла-де-Хименес (шт. Оахака. Мексика). Шаман Мария Сабина ест этот гриб в ходе ночной религиозной церемонии. Псилоцибин — химическое вещество, вызывающее цветные видения у съевшего такие «священные» грибы, — является структурным аналогом галлюциногенов ЛСД и мескалина (см. рис. 29-50)
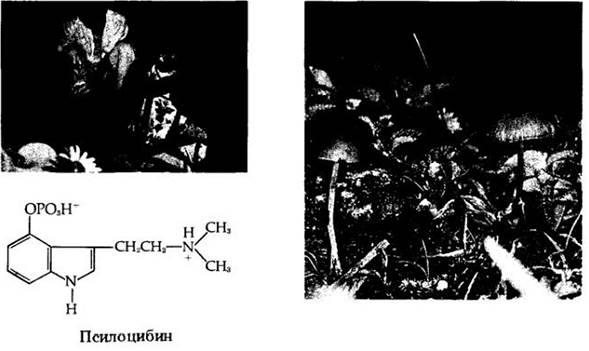
Рис. 13-35. Базидиомицеты из класса гастеромицетов. А. Дождевик Lycoper- don ericetorum. Споры высвобождаются из отверстия на верхушке каждого шаровидного базидиокарпа и разносятся ветром. Б. Веселка Phallus impudicus. Базидиоспоры погружены в дурно пахнущую липкую массу на верхушке базидиокарпа. Мухи (например, показанные здесь Mydaea urbana) посещают эти грибы в поисках пищи и распространяют споры, в большом количестве пристающие к их ногам и туловищу. В. Бокальчики Cyathus striatus. Их чашевидные базидиокарпы содержат базидиоспоры, которые выбрасываются наружу и разносятся дождевыми каплями. Г. Земляная звездочка Geastrum triplex. Внешний слой этого «дождевика» дольчато развернут

Класс Gasteromycetes
Гастеромицеты образуют базидиоспоры внутри базидиокарпов, полностью замкнутых по крайней мере на ранних стадиях развития. Этот класс, возможно, произошел от гименомицетов. Среди обычных его представителей — веселки, земляные звездочки, ложные трюфели, бокальчики и дождевики (рис. 13-35). Веселки (рис. 13-35, Б) имеют оригинальное строение. Они развиваются под землей в виде тугих мешочков («чертовых яиц»), а при созревании образуют из них надземные части — рецептакулы в виде ножки с глебой, представляющей собой наружный гимений с морщинистой поверхностью. Глеба покрыта дурно пахнущей липкой массой спор, привлекающих мух и жуков, которые и распространяют споры в своих фекалиях.
Дождевики — знакомые всем гастромицеты. При созревании их внутренняя часть высыхает и грибы, если до них дотронуться, выбрасывают тучу спор (рис. 13-35, А). Один дождевик может образовать несколько триллионов базидиоспор. Грибы бокальчики (рис. 13-35, В) начинают свое развитие как дождевики, но после разрушения большей части внутренней структуры становятся похожими на крошечные птичьи гнезда.
Класс Teliomycetes
Этот класс включает два крупных порядка: ржавчинные (Uredinaies) и головневые грибы (Ustilaginales). Ero представители отличаются от других базидиомицетов тем, что не образуют базидиокарпов. Однако у них возникают дикариотические гифы и базидии, которые септированы, как у дрожалковых грибов из класса гименомицетов. Ржавчинные и головневые (см. рис. 13-26) образуют свои споры в скоплениях, которые у первых мельче, называемых сорусы. Как патогены растений эти грибы имеют огромное экономическое значение, ежегодно нанося зерновым культурам во всем мире ущерб, оцениваемый миллиардами долларов.
Рис. 13-36. Неполное «ведьмино кольцо», образованное шампиньоном Agaricus arvensis. Установлено, что возраст некоторых «ведьминых колец» около 500 лет. Из-за обеднения почвы основными питательными веществами трава внутри кольца часто более низкорослая и светлая, чем за его пределами
Жизненный цикл многих ржавчинных очень сложен и постоянно изучается фитопатологами, в задачу которых входит поиск средств борьбы с ними. До недавнего времени считалось, что эти грибы облигатные паразиты сосудистых растений, но ряд их видов оказалось возможным выращивать и в искусственной культуре. Некоторые головневые также способны проходить полный цикл развития в лабораторных условиях. Хорошо известен жизненный цикл линейной (стеблевой) ржавчины злаков — Puccinia graminis, одного из 7000 видов порядка Uredinaies. Существует множество штаммов этого вида, которые паразитируют на пшенице и других злаках — ячмене, овсе, ржи и различных дикорастущих травах. Р. graminis— постоянная причина потерь урожаев пшеницы. За один год в Миннесоте, Северной и Южной Дакоте и в зоне канадских прерий они составили почти 8 млн. т. Еще в 100 г. Плиний писал об этой ржавчине как о «величайшем бедствии для зерновых культур». Сегодня фитопатологи в значительной степени снижают связанный с ней ущерб путем выведения устойчивых сортов пшеницы, но мутации и рекомбинации паразита делают любой достигнутый успех недолговечным. Парасексуальные процессы, ведущие к соматической рекомбинации, — один из факторов возникновения новых патогенных штаммов линейной ржавчины злаков.
Р. graminis — разнохозяйный паразит, т. е. для завершения жизненного цикла ему необходимы два различных хозяина (рис. 13-37). Однохозяйные паразиты, напротив, развиваются только на одном хозяине. Р. graminis может неограниченно долго обитать на злаке, размножаясь при этом только бесполым путем. Для полового размножения этого вида часть жизни он должен провести на барбарисе (Berberis), а другую — на злаке. В связи с этим предпринимались попытки избавиться от ржавчины, уничтожая барбарис. Например, в британской колонии Массачусетс вышел закон, согласно которому «каждый, на чьих землях растут кусты барбариса, должен уничтожить их или выкорчевать до 13 июня 1760 г».
Рис. 13-37. Жизненный цикл стеблевой ржавчины злаков Puccinia graminis. Этот разнохозяйный паразитический гриб часть жизни проводит на барбарисе, часть — на злаках (в данном случае — на пшенице). Ранней весной барбарис заражается одноядерными базидиоспорами. Плюс- и минус-гифы, возникающие из базидиоспор, образуют плюс- и минус-спермогонии. Когда спермации одного штамма вступают в контакт с воспринимающими гифами противоположного по знаку штамма, происходит плазмогамия и образуются эцидии. Дикариотические (n + n) эцидиоспоры, возникшие в эцидиях, инфицируют пшеницу, на которой вскоре после этого появляются красные полосы, уредини- умы, содержащие одноклеточные дикариотические урединиоспоры. Эти споры образуются в течение всего лета, вновь и вновь заражая пшеницу. С приближением осени красные полосы превращаются в темные телейтопустулы, содержащие двуклеточные дикариотические телейтоспоры, не являющиеся инфекционными. Ранней весной ядра в каждой половине телейтоспоры попарно сливаются (кариогамия). Когда две клетки телейтоспоры прорастают, происходит мейоз и каждая из них дает начало базидии с четырьмя гаплоидными базидиоспорами
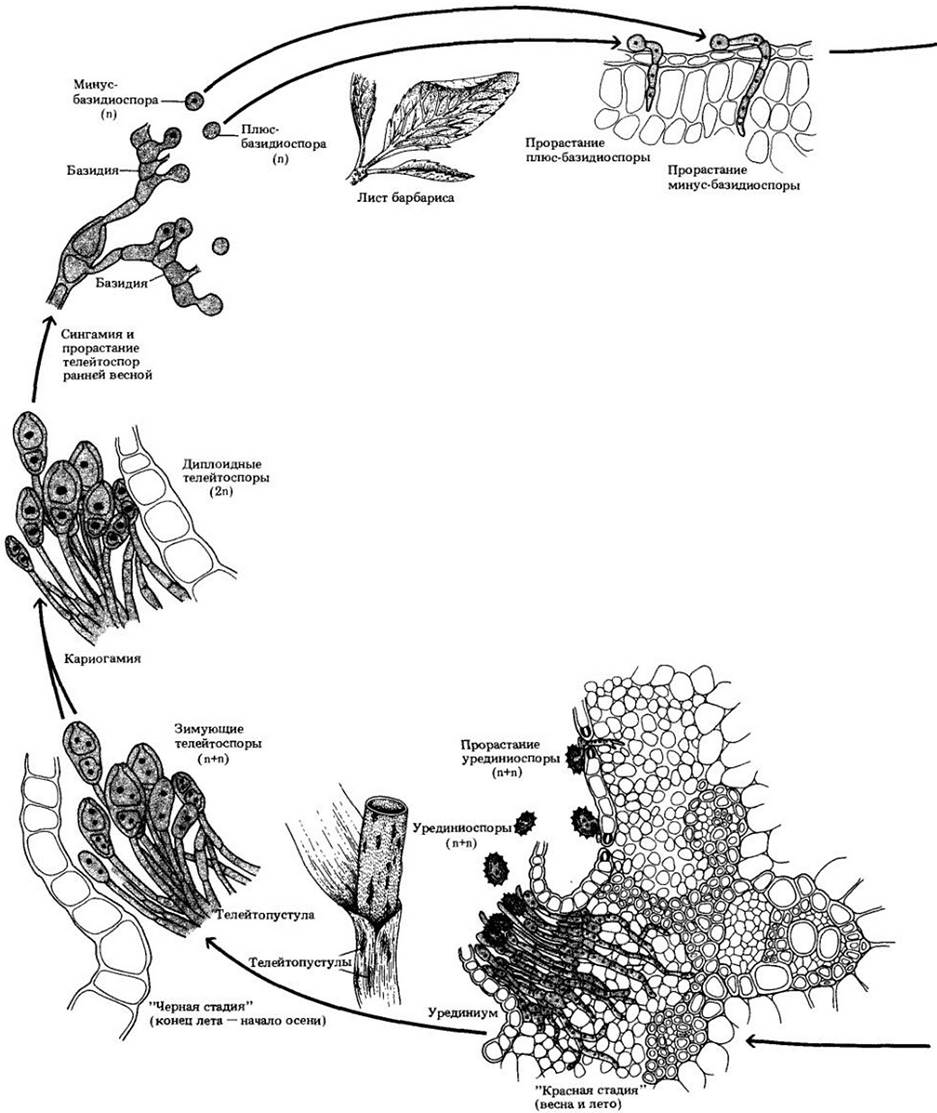

Заражение барбариса происходит весной, когда одноядерные базидиоспоры инфицируют растение, образуя в основном на верхней поверхности его листьев колбообразные спермогонии. Эта форма паразита включает плюс- и минус- штаммы, базидиоспоры и возникающие из них спермогонии также обозначаются «+» или «-». Каждый спермогоний содержит два типа гиф: один образует цепочки мелких клеток, называемых спермациями, а второй — так называемые воспринимающие гифы. Спермации выходят наружу через отверстие спермогония. Если плюс-спермаций одного спермогония вступит в контакт с минус-воспринимающей гифой другого или наоборот, произойдет плазмогамия и образуются дикариотические гифы, начинающие расти вниз из спермогония и формирующие в основном на нижней поверхности листа эцидии. В эцидиях возникают цепочки дикариотических эцидиоспор, заражающих впоследствии пшеницу.
Первое внешнее проявление инфекции на пшенице выражается в появлении полос ржавого цвета на листьях и стеблях («красная стадия»), так называемых урединиумов, содержащих одноклеточные дикариотические урединиоспоры. Такие споры образуются в течение всего лета, заражая другие растения пшеницы; они являются основным способом распространения ржавчины в тех районах мира, где возделывается эта культура. Поздним летом и ранней осенью красноватые сорусы постепенно темнеют, превращаясь в телейтопустулы с двуклеточными дикариотическими телейтоспорами («черная стадия»). Эти споры зимуют, не заражая ни пшеницу, ни барбарис. Ранней весной перед прорастанием два гаплоидных ядра каждой их клетки сливаются в одно диплоидное. С началом прорастания наблюдается мейоз, происходящий, по-видимому, в коротких цилиндрических базидиях, выходящих из двух клеток телейтоспоры. Образующиеся ядра отделяются друг от друга септами, а затем мигрируют в стеригмы и развиваются в базидиоспоры. Так завершается годовой цикл развития.
В некоторых районах жизненный цикл стеблевой ржавчины злаков укорачивается за счет устойчивости урединальной стадии, если в радиусе распространения урединиоспор постоянно присутствуют активно растущие ткани растений. На североамериканских равнинах эти споры с озимой пшеницы в юго-западных штатах и Мексике мигрируют на север, пока не достигают южной Манитобы. Более поздние их генерации рассеиваются на запад до Альберты, а в конце лета дрейфуют в южном направлении — «на зимовку», видимо, вдоль восточного склона Скалистых гор. При таких условиях сохранение ржавчины уже не зависит от второго хозяина. В то же время в Евразии, где перенос урединиоспор с юга невозможен из-за протяженных горных хребтов, идущих в широтном направлении, барбарис необходим для выживания этого паразита.