СОВРЕМЕННАЯ БОТАНИКА - П. РЕЙВН - 1990
РАЗДЕЛ IV. РАЗНООБРАЗИЕ
ГЛАВА 15. МНОГОКЛЕТОЧНЫЕ ПРОТИСТЫ: КРАСНЫЕ, БУРЫЕ И ЗЕЛЕНЫЕ ВОДОРОСЛИ
Открытое море, прибрежная зона и суша — три жизненные зоны, образующие нашу биосферу. В первых двух (древнейших) областях водоросли играют роль, сравнимую с ролью растений в гораздо более молодом наземном мире (рис. 15-1). Водоросли часто преобладают и в пресноводных водоемах — лужах, реках и озерах, внося иногда основной вклад в продуктивность этих экосистем. Где бы они ни росли, их экологическое значение не меньше, чем у растений в сухопутных местообитаниях.
Рис. 15-1. Водоросли на камнях при отливе (побережье Северной Каролины)

Вдоль каменистого морского берега можно обнаружить более крупные и более сложно организованные водоросли, как правило, образующие четко различимые горизонты в зависимости от положения в приливно-отливной зоне, или литорали (рис. 15-2). Сложность их структуры отражает приспособление к жизни в этой изменчивой среде, где дважды в сутки они испытывают сильные колебания влажности, температуры, солености, освещенности, а кроме того, удары прибоя и абразивное действие взвешенных частиц песка, поднимаемых волнами.
Рис. 15-2. Камни вдоль побережья Корнуолла в Англии. Можно видеть несколько вертикальных зон, образованных различными видами водорослей

Водоросли, прикрепленные к дну вне прибойной зоны, дают убежище разнообразнейшим микроорганизмам, а также рыбам и беспозвоночным, которые питаются этими микроорганизмами или друг другом. Такие водорослевые заросли по своей протяженности и густоте вполне можно сравнить с лесами. Например, вдоль побережья Калифорнии тянутся огромные массивы гигантских бурых водорослей с парящими в воде широкими пластинчатыми телами длиной до 15 м на извилистых заякоренных в дне «стеблях» длиной до 30 м и более. Многие крупные хищники, включая каланов и тунцов, находят в этих зарослях добычу и укрытие. Такие водоросли собираются и человеком, который использует их в пищу и как удобрение.
В данной главе обсуждаются три отдела водорослей, в которых все или часть представителей многоклеточные. Именно эти протисты образуют прибрежные массивы или свободно плавают в открытом океане, например, в так называемом Саргассовом море. Отдел зеленые водоросли (Chlorophyta) включает и одноклеточные формы, многие из которых вносят существенный вклад в продуктивность пресноводного и морского планктона. Этот исключительно разнообразный отдел представляет особый интерес еще и потому, что почти наверняка именно его виды около 430 млн. лет назад дали начало растениям. Хотя сейчас, возможно, и не существует зеленых водорослей, сходных с предками растений, многие формы сохраняют их отдельные предполагаемые черты.
Кроме зеленых водорослей, которые лучше представлены в пресной воде, чем в море, и обычны также в наземных местообитаниях, имеются еще два отдела, в основном многоклеточных протистов: красные и бурые водоросли (отделы Rhodophyta и Рhaeophytа). Оба они включают почти исключительно морские формы. Известно всего несколько родов красных и бурых водорослей, обитающих в пресной воде. Все бурые водоросли — многоклеточные, в то время как некоторые красные водоросли — одноклеточные.
Характеристика красных, бурых и зеленых водорослей
Как уже обсуждалось в предыдущей главе, хлоропласты у красных, бурых и зеленых водорослей произошли, вероятно, от разных бактерий. У красных водорослей они содержат хлорофилл а и фикобилины, напоминая древнюю группу цианобактерий. Многие ученые считают данные в пользу происхождения хлоропластов красных водорослей от симбиотических цианобактерий достаточно убедительными. В то же время зеленые водоросли с хлоропластами, содержащими хлорофиллы а и b, близки в этом отношении эвгленовым и своим потомкам растениям. По биохимическим характеристикам их хлоропласты сходны с бактерией Prochloron (см. рис. 11-2, 11-17) и, возможно, возникли из организма именно такого типа. Наконец, не исключено, что золотистобурые хлоропласты близкородственных бурых водорослей,
хризофитов и динофлагеллят с хлорофиллами а и с берут начало от бактерии, сходной с недавно открытым видом Heliobacterium chlorum. Сами хлоропласты здесь, возможно, произошли от поглощенных водорослями эукариотических клеток, но в конечном итоге их фотосинтезирующие органеллы имеют бактериальное происхождение.
Представители рассматриваемых в этой главе отделов сильно различаются многими другими признаками, некоторые из которых представлены в табл. 14-1. Хотя целлюлоза обнаружена в клеточных стенках некоторых видов каждого отдела, химический состав этих оболочек крайне неодинаков как у разных групп, так и внутри них. Красные водоросли на всех стадиях жизненного цикла лишены жгутиков и центриолей. Они имеют структуры, называемые полярными кольцами, связанные с ядрами и движущиеся в профазе митоза подобно центриолям; гомологичны ли они последним — неизвестно. Черты сходства между красными водорослями и аскомицетами отмечались уже давно и требуют дальнейшего изучения с привлечением дополнительных признаков. Жгутиковые клетки бурых водорослей, как и у близкородственных им хризофитов и диатомовых, имеют разные жгутики — направленный вперед перистый и гладкий задний (см. рис. 2- 26). Жгутики зеленых водорослей (когда они имеются), напротив, почти всегда одинаковые и гладкие.
Зеленые водоросли, как и растения, запасают питательные вещества в виде крахмала в пластидах (рис. 15-3, А). Все прочие группы фотосинтезирующих эукариот, включая рассмотренные в гл. 14, красные и бурые водоросли, откладывают запасы вне хлоропластов в цитоплазме, причем красные водоросли в виде крахмала, а бурые — ламинарина, также полиглюкозида, но не с такими, как у крахмала, связями между остатками глюкозы. Кроме того, бурые водоросли запасают спирт маннитол.
Когда клетки водорослей делятся, их плазматические мембраны обычно перешнуровываются, как и у животных, грибов и протистов. Однако у одной бурой водоросли и нескольких родов нитчатых зеленых водорослей были обнаружены клеточные пластинки, как у растений. В большинстве случаев (исключение — красные водоросли) присутствуют центриоли, становящиеся базальными тельцами жгутиков у подвижных клеток (см. с. 36).
Рис. 15-3. У зеленых водорослей и высших растений запасные питательные вещества накапливающей в виде крахмала в пластидах, тогда как у других фотосинтезирующих эукариот аналогичные запасы возникают в ципюплазме. А. Chlorella, зеленая водоросль с крупными светлыми зернами крахмалю в пластидах. В. Окруженная пластидами масса крахмальных зерен в цитоплазме красной водоросли Batrachospermum moniliforme

В целом многоклеточные организмы рассматриваемых здесь отделов не имеют в отличие от сосудистых растений сложно организованных тканей. Однако у некоторых бурых водорослей обнаружен центральный проводящий тяж из клеток, напоминающих ситовидные элементы стебля и листовой пластинки. Репродуктивные структуры водорослей, как правило, одноклеточные, а не многоклеточные со стерильными защитными покровами, как у моховидных и сосудистых растений (см. гл. 16).
Три рассматриваемых отдела водорослей — древние группы, резко различающиеся своими признаками. Ископаемые формы, сходные с одноклеточными зелеными водорослями, найдены в отложениях формации Биттер-Спрингс в центральной Австралии возрастом около 900 млн. лет. В кембрийский период, т. е. по крайней мере 550 млн. лет назад, появились крупные сифоновые (ценоцитные) зеленые, а также красные водоросли. Также в палеозое возникли многоклеточные линии зеленых водорослей. Например, древнейшие харовые, представители наиболее своеобразной их группы, датируются поздним силуром (около 420 млн. лет назад). Эти зеленые и красные водоросли, лучше всего представленные ископаемыми отпечатками, имеют обызвествленные клеточные стенки. Такие стенки известны только у одного рода бурых водорослей, поэтому ископаемые остатки этой группы относительно редки; они не сохранились так хорошо, как у некоторых зеленых и красных водорослей.
Отдел Rhodophyta
Красные водоросли, или багрянки, имеют сложно организованные клетки без жгутиков и сложные жизненные циклы. Они лишены центриолей, отличаясь этим от зеленых и бурых водорослей, у которых центриоли обычно присутствуют даже в вегетативных клетках. У багрянок их функцию выполняют полярные кольца.
К этому отделу относятся около 4000 видов, особенно обильные в тропических и теплых водах, хотя многие виды обитают в холодных областях земного шара. Менее 100 видов обнаружены в пресной воде, но в море их число больше, чем у всех прочих групп многоклеточных морских водорослей, вместе взятых. Представители этого отдела обычно прикреплены к камням или другим водорослям; имеется несколько свободноплавающих, а также одноклеточных колониальных видов.
Характерную окраску красным водорослям придают водорастворимые фикобилины — вспомогательные пигменты, маскирующие цвет хлорофилла а. Эти организмы особенно хорошо приспособлены к поглощению зеленых, фиолетовых и синих лучей света, проникающих на большую глубину. Их хлоропласты, по-видимому, произошли от симбиотических цианобактерий, с которыми они сходны биохимически и структурно.
Клеточные оболочки большинства красных водорослей содержат жесткие микрофибриллы целлюлозы или другого полисахарида, погруженные в слизистый матрикс, обычно из сульфатированного полимера галактозы, например, агара или каррагинана (см. «Экономическое использование морских водорослей», с. 257). Именно последние компоненты делают красные водоросли гибкими и скользкими на ощупь.
Кроме того, многие багрянки откладывают в своих клеточных стенках карбонат кальция. Среди них особенно жесткие и «каменистые» образуют крупное семейство кораллиновых (Соrallinасеае). Они широко распространены в океанах на стабильных, достаточно освещенных субстратах (рис. 15- 5, Б, В). Недавно корковая кораллиновая водоросль была отмечена на глубине 268 м; это рекордная отметка для фотосинтезирующих организмов (рис. 15-6). Она почти на 100 м ниже глубины, куда обычно проникает солнечный свет.
Кораллиновые водоросли играют важную роль в образовании коралловых рифов. Действительно, продуктивность таких рифов и их способность произрастать в относительно бедных питательными веществами тропических водах непосредственно зависит от этих водорослей и симбиотических динофлагеллят (зооксантелл), обитающих внутри полипов (см. рис. 14-20). Рифообразующие кораллиновые обычно не членистые (рис. 15-5, В), в то время как многие другие представители этого семейства имеют хорошо заметное членистое строение (рис. 15-5, Б). Некоторые кораллиновые, подобно накипным лишайникам, образуют корки на камнях (рис. 15-6). Эта древняя группа известна уже из позднего кембрия, т. е. ее возраст превышает 500 млн. лет.
Большинство багрянок состоит из нитей, которые зачастую плотно переплетены и удерживаются вместе слизистым межклеточным матриксом. Более простые формы, например, пресноводная водоросль Batrachospermum, представлены нс собранными в такие пучки одиночными нитями (рис. 15-4). Рост у нитчатых багрянок начинается от куполообразной апикальной клетки, которая последовательно делится, образуя ось водоросли. Эта ось затем образует мутовки боковых ветвей (рис. 15-4, Б). Большинство красных водорослей многоосевые с внешне паренхиматозным трехмерным телом. У таких форм нити, вторично соединяясь между собой, образуют сеть, скрепленную слизистым матриксом. Для многих красных водорослей характерны особые поровые соединения клеточных стенок, представляющие собой линзовидные пробки, удерживающиеся в стенках за счет своих экваториальных перетяжек (рис. 15-8). У нескольких родов (например, у Porphyra, см. с. 258) обнаружена настоящая паренхима из плотно упакованных клеток, как у высших растений.
Рис. 15-4. А. Простая нитчатая багрянка Batrachospermum. Мягкие слизистые разветвленные нити этого пресноводного протиста встречаются в основном в холодных ручьях, прудах и озерах по всему земному шару. Б. Апекс нити Batrachospermum с мутовками боковых ветвей. В. Два цистокарпа Batrachospermum

Рис. 15-5. А. У Bonnemaisonia hamifera четко видна типичная нитчатая структура красных водорослей. По- видимому, многоклеточностъ появилась у багрянок независимо от зеленых и бурых водорослей, у которых подобная организация не обнаружена. Разветвленные нити этой красной водоросли несут крючки, позволяющие ей цепляться за другие водоросли. Б. Членистые кораллиновые водоросли в водоеме, оставшемся во время отлива на калифорнийской литорали. В. Рифообразующая нечленистая кораллиновая водоросль Porolithon craspedium. Г. «Ирландский мох» (Chondrus crispus), важный источник каррагинана и других коллоидов

Рис. 15-6. А. Фотография в сканирующем электронном микроскопе неидентифицированного вида красной водоросли, собранной с глубины 268 м Марком Литтлером и его коллегами из Смитсоновского института в октябре 1984 г. Эта водоросль образует пятна около 1 м диаметром на подводной горе у Багамских островов при освещенности примерно 0,0005% ее значения на поверхности океана, покрывая около 10% поверхности скалистого субстрата. Лабораторные исследования показали, что она примерно в 100 раз более эффективно поглощает и использует свет, чем ее сородичи, обитающие на мелководье. Эта водоросль, растущая без перерывов до вершины подводной горы (глубина около 70 м), обитает приблизительно на 100 м глубже нижнего предела жизни, установленного ранее для любых прикрепленных фотосинтезирующих организмов. Б. Пурпурные корковые кораллиновые и другие глубоководные водоросли с этой же подводной горы. В. Подводный исследовательский аппарат «Джонсон Си Линк I», с помощью которого были получены самые глубоководные снимки водорослей

Рис. 15-7. Жизненный цикл Polysiphonia, широко распространенной морской багрянки. Органы полового размножения возникают у верхушек ветвей гаплоидных гаметофитов, которые образуются из гаплоидных тетраспор. Из сперматангиев, расположенных плотными пучками, выходят клетки, функционирующие как спермации. Расширенная базальная часть карпогония содержит ядро и играет роль яйцеклетки. После оплодотворения в результате митоза внутри карпоспорангия образуются диплоидные карпоспоры, выходящие наружу через отверстие в перикарпе, т. е. во внешней развивающейся вокруг них структуре. Перикарп образуется из карпоспорофита. Карпоспоры прорастают в тетраспорофиты, сходные по размерам и строению с гаметофитами. На тетраспорофитах формируются тетраспорангии, в каждом из которых происходит мейоз с образованием четырех гаплоидных тетраспор. После этого цикл начинается заново

Жизненный цикл большинства красных водорослей необычен тем, что включает три фазы: (1) гаплоидный гаметофит, (2) диплоидную фазу — карпоспорофит и (3) другую диплоидную фазу, называемую тетраспорофитом (рис. 15- 7). Мужская особь гаметофитного поколения образует сперматангии (антеридии), из которых выходят неподвижные спермации — мужские гаметы, пассивно плавающие в воде. На женском половом органе, карпогонии, развивается длинный волосовидный вырост — трихогина, аналогичная по структуре и функции трихогинам аскомицетов (гл. 13). Спермаций попадает на трихогину случайно с током воды и прикрепляется к ней. После этого его ядро проникает в трихогину через пору, мигрирует к ядру яйцеклетки и сливается с ним.
У просто организованных багрянок типа Batrachospermum диплоидное поколение — карпоспорофит (рис. 15-4, В) — развивается прямо из оплодотворенного карпогония, на котором возникают концевые спорообразующие структуры — карпоспорангии. У более продвинутых групп диплоидное ядро, возникшее внутри карпогония в результате синга- мии, переносится во вторую, так называемую ауксиллярную клетку, из которой и развивается карпоспорофит. Такой жизненный цикл наблюдается у рода Polysiphonia (рис. 15-7).
Диплоидные карпоспоры, формирующиеся внутри карпоспорангия, выходят из него и превращаются в свободноживущий тетраспорофит. На нем возникает еще один тип спорообразующей структуры — тетраспорангий, в котором происходит мейоз и образуются четыре гаплоидные тетраспоры. Прорастая, каждая их них дает гаметофит.
У большинства красных водорослей гаметофит, кариоспорофит и тетраспорофит очень сходны, т. е. являются изоморфными, например, у Polysiphonia и кораллиновых. Сейчас описывается все больше гетероморфных циклов, когда тетраспорофиты имеют микроскопическую нитчатую структуру или образуют тонкую корку, плотно прикрепленную к каменистому субстрату.
Рис. 15.8. Поровые соединения между клетками красной водоросли Palmaria. Они имеют вид хорошо заметных линзовидных пробок с белковой сердцевиной и по крайней мере частично полисахаридными покровными слоями. Такие связи образуются между сестринскими клетками и соседними нитями, объединяя их в единое тело красной водоросли. Они возникают также между многими паразитическими багрянками и их хозяевами, возможно, способствуя переносу между ними ассимилятов
Отдел Рhaeophyta
Бурые водоросли — это почти целиком морская группа, включающая большинство крупных по размерам протистов умеренных областей. Объединяя всего около 1500 видов, она представляет значительный интерес, так как преобладает на каменистых прибрежных субстратах в холодных районах земного шара (рис. 15-9). Крупные виды из порядка ламинариевых (Laminariales) образуют вдоль берегов обширные заросли. В прозрачной воде бурые водоросли процветают от линии отлива до глубины 20 — 30 м и при слабом уклоне дна могут распространяться на 5 — 10 км от берега. Даже в тропиках, где бурые водоросли встречаются реже, род образует плотные плавающие массы (рис. 15-10), например, в Саргассовом море, т. е. в Атлантическом океане северо-восточнее Антильских островов.
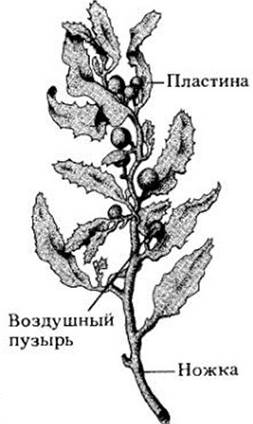
Рис. 15-9. Бурые водоросли. A. Durvillea antárctico во время отлива на каменистой литорали Новой Зеландии. Б. Фрагмент ламинарии; видны прикрепительные структуры, ножки и основания пластин. В. Fucus vesiculosas, густо покрывающий многие каменистые участки, обнажающиеся во время отлива. Когда водоросли заливает вода, наполненные воздухом пузыри на пластинках выносят их к свету. Скорость фотосинтеза у часто оказывающихся на воздухе морских водорослей может быть в семь раз выше в воздушной среде, чем в воде; в то же время у видов, почти постоянно пребывающих под водой, эта скорость выше в погруженном состоянии. Частично это объясняет вертикальное распределение морских водорослей в приливно-отливной зоне
Рис. 15-10. Бурая водоросль Sargassum имеет сложную организацию. Этот род, как и фукус, относится к порядку Fucales и имеет жизненный цикл, показанный на рис. 15-14. Два вида этого рода, не размножающиеся половым путем, образуют громадные, свободно плавающие массы в Саргассовом море; другие прикрепляются к камням вдоль берегов
Размеры бурых водорослей колеблются от микроскопических до самых крупных среди всех морских водорослей. Самые простые по строению представители отдела имеют форму разветвленных нитей (например, Ectocarpus, рис. 15- 11), растущих с помощью внутренних интеркалярных меристем. У более высокоорганизованных нитчатых бурых водорослей нити объединены в плотное тело, а у самых сложных форм имеется трехмерная клеточная структура, сходная с наблюдаемой у растений.
Рис. 15-11. Ectocarpus — бурая водоросль с простыми разветвленными нитями. Эта микрофотография Е. siliculosus показывает одногнездные спорангии (короткие округлые светлоокрашенные структуры) и многогнездные спорангии (удлиненные темные структуры), которые образуются на спорофитах. Мейоз происходит внутри одногнездных спорангиев (на фотографии из них уже вышли зооспоры). Эти водоросли обитают на мелководье и в эстуариях по всему миру: от холодных арктических и антарктических областей до тропиков

Крупные бурые водоросли, в частности Laminaria, состоят из листоподобных пластинок, ножки и структуры для прикрепления к субстрату, причем зона меристемы находится между пластинкой и ножкой (рис. 15-9, Б). Этот тип роста особенно важен для практического использования представителей Macrocystis, собираемых вдоль побережья Калифорнии; когда промысловики срезают отросшие пластины этих водорослей, их остающиеся глубже части, регенерируют. Гигантские представители родов Macrocystis и Nereocystis могут достигать в длину свыше 60 м и растут очень быстро, давая большое количество добываемой биомассы. Один из наиболее важных получаемых из них продуктов — альгинат, слизистое межклеточное вещество, используемое как стабилизатор и эмульгатор в пищевой промышленности и красильном деле, а также для покрытия бумаги.
Фукус (Fucus, рис. 15-9, В) — это дихотомически ветвящаяся бурая водоросль с воздушными пузырями на концах пластин. Дифференцировка его тела сходна с наблюдаемой у ламинариевых. Фукусу родственны виды Sargassum (рис. 15- 10), особи которых часто отрываются от субстрата, образуя плавающие скопления без каких-либо прикрепительных структур. Fucus и Sargassum в отличие от ламинариевых растут за счет деления единственной апикальной клетки, а не интеркалярных меристем.
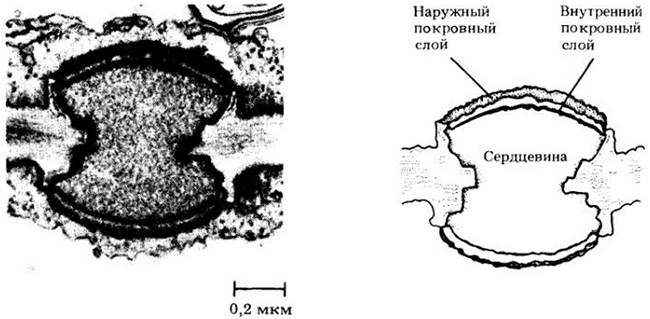
Внутреннее строение ламинариевых довольно сложное. У некоторых из них в центре ножки находятся удлиненные клетки, модифицированные для проведения питательных веществ и напоминающие флоэмные элементы сосудистых растений (рис. 15-12). Они имеют ситовидные пластинки и могут транспортировать ассимиляты из пластинки, лежащей вблизи поверхности воды, в слабо освещенные ножку и прикрепительную часть, расположенные гораздо глубже, со скоростью около 60 см/ч. У многих относительно массивных ламинариевых происходит также латеральный транспорт из внешних фотосинтезирующих слоев к внутренним клеткам. В основном переносятся спирт маннитол и аминокислоты.
Рис. 15-12. Некоторые бурые водоросли, например, гигантский представитель ламинариевых Macrocystis pyriferа, в процессе эволюции приобрели ситовидные трубки, сходные с имеющимися у сосудистых растений. А. Поперечный разрез, показывающий строение ситовидной пластинки. Б. Продольный разрез через ножку с ситовидными трубками. На данном препарате ситовидные пластинки кажутся утолщенными, так как покрыты натеком вещества клеточной стенки
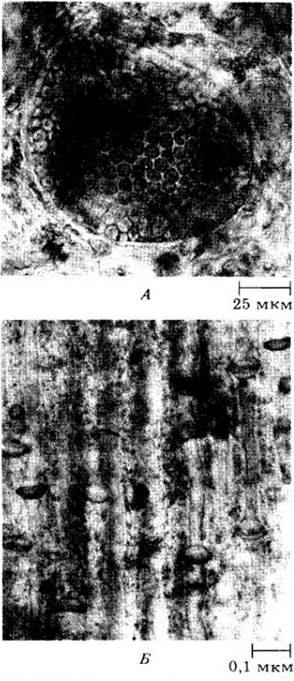
Как уже говорилось в гл. 14, бурые водоросли близкородственны хризофитам (отдел Chrysophyta), поэтому многие ботаники сейчас склоняются к объединению этих групп в один отдел. Общие их черты приведены в табл. 14-1. Некоторые хризофиты явно напоминают гипотетических одноклеточных предшественников бурых водорослей.
Жизненные циклы большинства бурых водорослей включают чередование поколений с мейозом на стадии спорообразования (см. рис. 10-11, В). Гаметофиты формируют многоклеточные репродуктивные структуры, многогнездные гаметангии, образующие мужские или женские гаметы, или жгутиковые гаплоидные споры, которые дадут начало новому гаметофиту. Спорофиты диплоидны и формируют как многогнездные, так и одногнездные спорангии (рис. 15-11). В многогнездных спорангиях возникают диплоидные зооспоры, дающие новые спорофиты. Внутри одногнездных спорангиев происходит мейоз с образованием гаплоидных зооспор, которые прорастают в гаметофиты.
В роде Ectocarpus гаметофит и спорофит одинаковы по размеру и форме (изоморфны). У многих крупных бурых водорослей, включая ламинариевых, чередуются гетероморфные поколения — крупный спорофит и микроскопический гаметофит (рис. 15-13). У Laminaria одно гнездные спорангии возникают на поверхности зрелых пластин. Половина образуемых зооспор прорастает в мужской гаметофит, другая — в женский. Многогнездные гаметангии, формирующиеся на этих гаметофитах, в процессе эволюции видоизменились в одноклеточные антеридии, дающие по одному спермию, и также одноклеточные оогонии с одной яйцеклеткой. Оплодотворенная яйцеклетка здесь остается прикрепленной к женскому гаметофиту, где развивается в новый спорофит. Молекулы веществ, выделяемые женскими гаметами для привлечения мужских, у некоторых родов бурых водорослей являются олефиновыми углеводородами с открытой цепочкой и одной или несколькими двойными связями.
У фукуса уникальный жизненный цикл (рис. 15-14) с мейозом на стадии образования гамет (см. рис. 10-11, Б). Мейоз в зиготе, типичный для двух крупных классов зеленых водорослей, у бурых водорослей неизвестен.
Рис. 15-13. Жизненный цикл бурой водоросли Laminaria. Подобно большинству бурых водорослей, ламинария имеет чередование гетероморфных поколений с крупным спорофитом. Подвижные гаплоидные зооспоры образуются в спорангиях в результате мейоза. Из них вырастают микроскопические нитчатые гаметофиты, дающие в свою очередь подвижные спермии и неподвижные яйцеклетки. У более простых бурых водорослей спорофит и гаметофит часто сходны, т. е. происходит чередование изоморфных поколений

Рис. 15-14. У фукуса гаметангии образуются в особых камерах (концептакулах), которые находятся в фертильных зонах (рецептакулах) на концах ветвей диплоидных особей. Имеются два типа гаметангиев — оогонии и антеридии. За мейозом сразу следует митоз, причем в оогонии образуются 8 яйцеклеток, а в антеридии 64 сперматозоида. Затем яйцеклетки и спермии выходят в воду, где и происходит оплодотворение. Мейоз гаметический, поэтому зигота непосредственно дает начало новой диплоидной особи
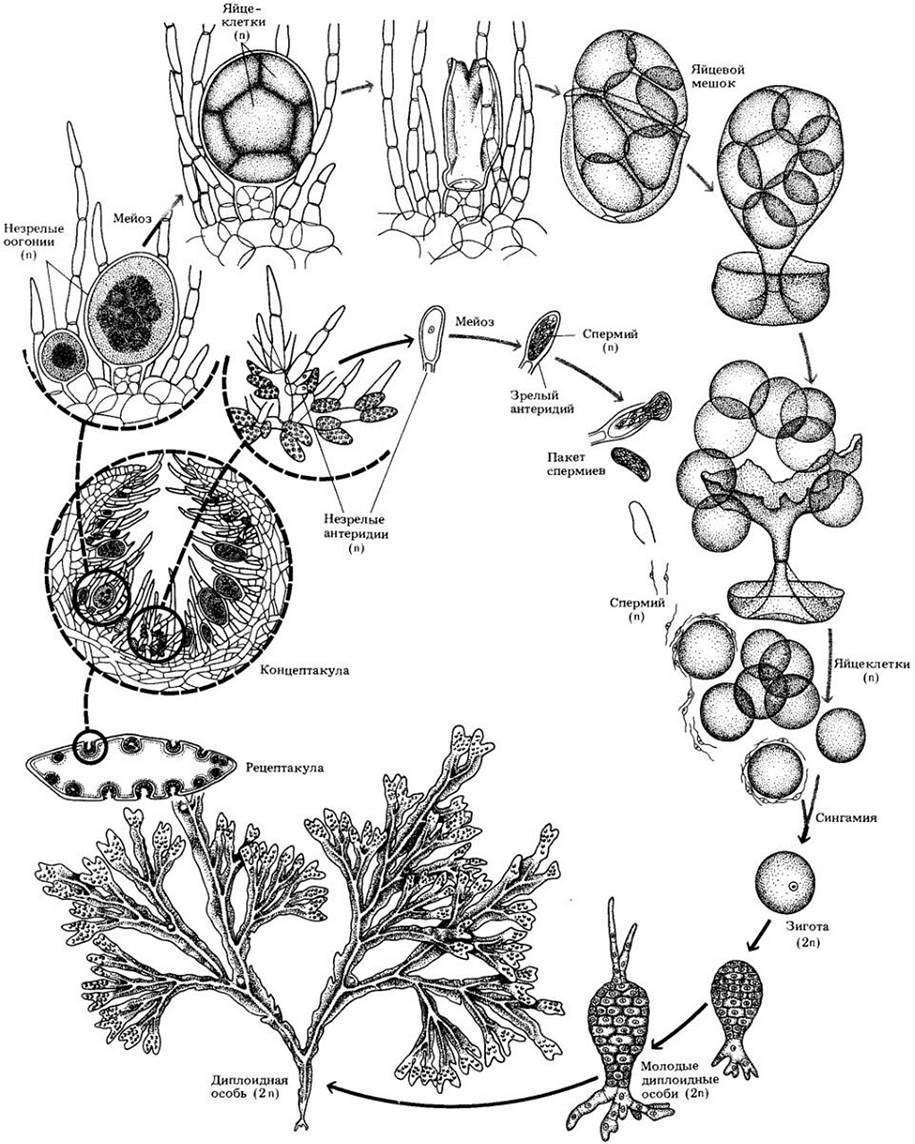
Отдел Chlorophyta
Зеленые водоросли — наиболее разнообразная группа из всех водорослей как по строению, так и по жизненному циклу. Она объединяет около 7000 видов; большинство их обитает в воде, но некоторые и в других местообитаниях, в том числе на поверхности снега, стволов деревьев, в почве, в симбиотических ассоциациях с лишайниками, простейшими и гидрами. Известны зеленые водоросли, например, виды одноклеточного рода Chlamydomonas, растущие на снегу, или нитчатая Trentepohlia, поселяющаяся на ветвях деревьев, которые образуют для защиты от яркого света большое количество каротиноидов. Из-за наличия этих вспомогательных пигментов они часто выглядят красными или оранжевыми. Большинство зеленых водорослей обитает в пресной воде, но несколько групп—сугубо морские. Размер многих представителей отдела микроскопический, но некоторые морские виды, например, мексиканский Codium magnum, достигают в ширину 25 см при длине более 8 м.
Зеленые водоросли напоминают растения некоторыми важными признаками. Они содержат хлорофиллы а и b, накапливают запасной крахмал внутри пластид (это характерно только для них и растений), имеют жесткие клеточные стенки, образованные у некоторых родов целлюлозой, пропитанной матриксом из гемицеллюлоз и пектиновых веществ. Более того, детали связи жгутика с базальным тельцем у спермиев растений и клеток ряда зеленых водорослей совпадают, что подтверждает гипотезу о прямом родстве этих двух групп. На основании перечисленных и других аргументов полагают, что зеленые водоросли дали начало растениям.
Исследования с помощью электронной микроскопии значительно расширили представление о разнообразии зеленых водорослей. Представители этого отдела разделяются на ряд эволюционных линий, независимо возникших от одноклеточных жгутиковых предков. Хотя хлоропласты сохраняют одинаковые черты, по другим признакам наблюдаются чрезвычайно сильные различия. Общий предок (если он был), возможно, имел жгутик и покровные чешуйки, которые в процессе эволюции слились в единую клеточную стенку, поэтому одноклеточные жгутиковые формы считаются наиболее примитивными в отделе. Они входят в класс Мicromonadophyсеае.
Клеточное деление у зеленых водорослей
У представителей самого крупного класса зеленых водорослей, Сhloropyсеае, уникальный тип клеточного деления с участием фикопласта (рис. 15-15). Дочерние ядра здесь движутся навстречу друг другу, в то время как неустойчивое веретено разрушается и параллельно плоскости клеточного деления образуется новая система микротрубочек — фикопласт. Вероятно, он должен гарантировать формирование борозды деления между дочерними ядрами. Ядерная оболочка в процессе митоза сохраняется. У подвижных клеток жгутики прикреплены к переднему концу. Внутри таких клеток находится система корешковых волокон жгутиков, образованная крестообразно расходящимися четырьмя узкими пучками микротрубочек (рис. 15-16). Эти пучки начинаются от передних базальных телец или вблизи них.
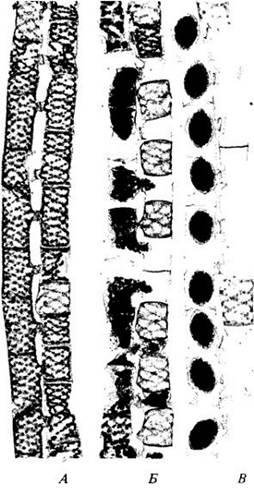
Рис. 15-15. Деление клетки у двух классов зеленых водорослей. В классе Chlorophyceae (А, Б) веретено деления отсутствует (не сохраняется) и дочерние ядра, расположенные относительно близко друг от друга, разделяются фикопластами. А. Деление клетки путем перешнуровывания. Б. Деление клетки путем формирования клеточной пластинки. В классе Charophyceae (В. Г) веретено деления устойчиво и дочерние ядра относительно отдалены друг от друга. В. Деление клетки путем перешнуровывания. Г. Присутствие фрагмопласта и деление с образованием клеточной пластинки. У Ulvophyceae веретено устойчиво, как у Charophyceae, но не формирует фрагмопласта
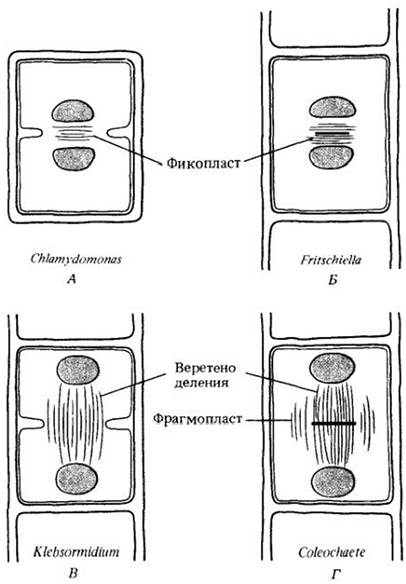
Рис. 15-16. Схема крестообразною расположения микротрубочек, связанных с центриолями (базальными тельцами) жгутиков; этот признак характерен для зеленых водорослей класса Сhlorophyсеае
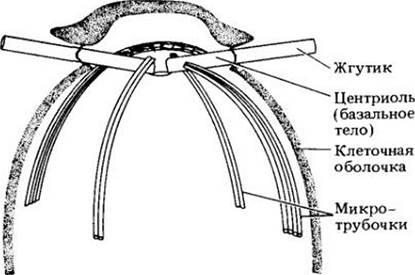
У Других зеленых водорослей веретено при делении клетки сохраняется, а в некоторых случаях образуется фрагмопласт. Микротрубочки фрагмопласта ориентированы перпендикулярно плоскости деления (рис. 15-15). Веретено остается до тех пор, пока не «разорвется» либо клеточной пластинкой, которая появляется в центре клетки и растет к ее краям, либо бороздой деления. Этот способ менее специализированный, чем с участием фикопласта. Типу клеточного деления придается большое значение в систематике зеленых водорослей.
Классификация зеленых водорослей
Небольшой класс одноклеточных Мicromonadaphyceae, чешуйчатых или голых жгутиковых форм, характеризуется многими, предположительно, предковыми чертами этого отдела. Кроме него современные альгологи выделяют еще четыре класса зеленых водорослей, каждый из которых, как считается, представляет собой самостоятельную ветвь эволюции. Один из них, Pleurastophyceae (см. с. 258) — небольшая группа, не рассматривается здесь в деталях. Остальные три класса — обширные группы, включающие большое число хорошо изученных организмов.
Три основных класса зеленых водорослей — Сharophусеае, Ulvophусеае и Сhlorophусеае — отличаются друг от друга рядом существенных особенностей. У Сhlorophусеае, образующих фикопласт, нити веретена в анафазе всегда укорачиваются, а у Сharophусеае и Ulvophyусеае (сохраняющих при митозе ядерную оболочку) этого обычно не происходит. У последних двух классов ядерная оболочка разрушается в начале митоза, как у растений, а у Сhlorophусеае сохраняется в течение всего этого процесса. Каждый класс включает одноклеточные жгутиковые формы, являющиеся либо целыми организмами (Сhlorophусеае), либо репродуктивными клетками — у двух других классов. Предковые формы каждого класса наверняка были одноклеточными жгутиковыми, а многоклеточные организмы появились в каждом из них независимо.
Сharophусеае и Сhlorophусеае обитают в основном в пресной воде, а Ulvophусеае — преимущественно в морс. Подвижные клетки в этих трех классах сильно различаются: у Сharophусеае они асимметричны, а у двух других целиком или почти радиально-симметричны. Жгутики у Сharophусеае боковые или субапикальные, т. е. отходят вблизи верхушки клетки (апекса), и направлены вбок под прямыми углами к ее оси. У Сhlorophусеае и Ulvophусеае они апикальные и направлены вперед (рис. 15-17).
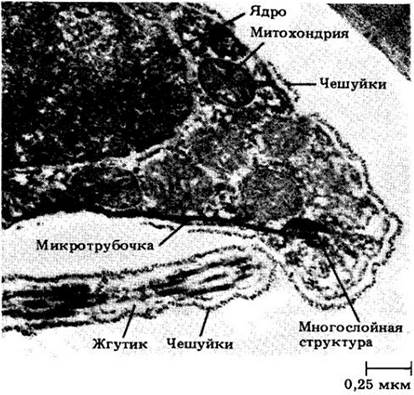
Рис. 15-17. Электронная микрофотография передней части спермин зеленой водоросли Сoleochaete; видна многослойная структура рядом с митохондриями. Слой микротрубочек идет от этой структуры к заднему концу клетки, лишенной стенки, и служит для нее цитоскелетом. Жгутиковая и клеточная мембраны покрыты слоем мелких ромбовидных чешуек
Внутри клеток Сharophусеае находится система микротрубочек в форме плоской широкой полосы, начинающейся от многослойной структуры вблизи латерально расположенных базальных телец и расходящейся к нижнему концу клетки. Аналогичное образование обнаружено и в подвижных спермиях растений. Более того, в пероксисомах у Сharophусеае образуется фотодыхательный фермент гликолатоксидаза, который, насколько известно, локализован там только у растений, хотя вне пероксисом обнаружен и у некоторых зеленых водорослей. Эти и другие признаки теснее связывают Сhlarophусеае с растениями, чем прочие классы зеленых водорослей, поэтому форму, наиболее близкую к предку растений, по-видимому, нужно искать именно среди представителей данного класса.
Половой процесс у Сharophусeае и Сhlorophусеаe всегда включает образование покоящейся зиготы (зигоспоры), в которой происходит мейоз (см. рис. 10-11, А). В то же время половое размножение у Ulvophусеае часто связано с чередованием поколений и мейозом на стадии споры, а покоящиеся зиготы редки. Жизненный цикл Ulvophусеае, по-видимому, наиболее продвинутый среди этих классов, а у Сharophусеае — самый примитивный.
Класс Сharophусеае
К этому классу принадлежат одноклеточные, малоклеточные, нитчатые или паренхиматозные представители, родство которых подтверждается многими важными структурными и биохимическими признаками. Среди них — наличие асимметричных жгутиковых клеток с латеральными или субапикальными жгутиками, расположенными под прямым углом к оси клеток; мейоз на стадии зиготы и образование покоящихся зигоспор; наличие многослойной структуры в подвижных клетках, а также особенности клеточного деления. Все Сharophусeае имеют устойчивое митотическое веретено, а у некоторых возникает фрагмопласт, способствующий формированию новой клеточной пластинки.
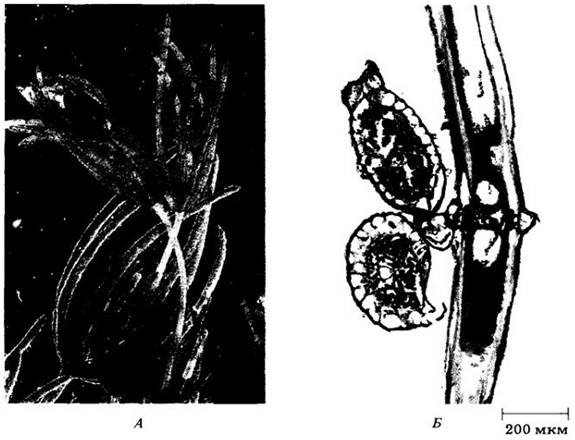
Спирогира (Spirogyra, рис. 15-18) — хорошо изученный род неразветвленных нитчатых водорослей, часто образующих пенистые или слизистые плавающие скопления в пресных водоемах. Каждая нить покрыта водянистым, слизистым на ощупь чехлом. Название рода указывает на наличие одной или нескольких спирально закрученных лент хлоропластов внутри каждой одноядерной клетки. Хлоропласты содержат многочисленные пиреноиды, т. е. дифференцированные участки, являющиеся центрами образования крахмала у зеленых водорослей, но почти всегда отсутствующие в хлоропластах растений. Недавние исследования показали, что у двух других зеленых водорослей здесь концентрируется фермент рибулозобисфосфаткарбоксилаза (см. с. 102). Пиреноиды, возможно, связаны также с превращением сахаров в крахмал, который обычно окружает эти структуры.
Рис. 15-18. А. Половое размножение у Spirogyra требует образования конъюгационных мостиков между клетками сближенных нитей. Б. Содержимое клеток мину с штамма переходит через эти мостики в клетки плюс-штамма. В. Внутри последних происходит сингамия, зигота образует толстую устойчивую стенку и превращается в зигоспору. Вегетативные нити спирогиры гаплоидны и мейоз происходит во время прорастания зигоспор, как и у всех Сharophусеае
Бесполое размножение у спирогиры происходит путем деления клеток и фрагментации нитей, причем ни на одной стадии жизненного цикла нет жгутиковых клеток. Половое размножение по морфологическим признакам нужно отнести к изогамии, однако при этом одна изогамета ведет себя как мужская репродуктивная клетка, двигаясь по конъюгационной трубке, чтобы слиться с другой изогаметой. Мейоз зиготический, с образованием покоящихся зигоспор, как у всех Сharophусeае.
Десмидиевые — это крупная группа пресноводных зеленых водорослей, родственных спирогире; у них также нет жгутиковых клеток. Некоторые десмидиевые — нитчатые, но большинство одноклеточные. Строение их клетки необычно тем, что ее стенка состоит из двух половин с узкой
перетяжкой между ними (рис. 15-19). Деление клетки, как и у спирогиры, идет с образованием устойчивого веретена и борозды деления. Некоторые систематики считают, что число видов десмидиевых превышает 10 000.
Рис. 15-19. У этой десмидиевой водоросли из рода Micrasterias каждая клетка имеет глубокую перетяжку, как и у большинства представителей этой обширной группы одноклеточных пресноводных Сharophусeае

Сharophyceae, сходные по строению с высшими растениями
Две группы зеленых водорослей, рассматриваемые ниже, больше всего напоминают растения деталями клеточного деления, а именно фрагмопластом и разрушением ядерной оболочки в начале митоза. Кроме того, они, как и растения, оогамны. Однако ни одна их этих групп не является их прямым предком, так как каждая в некоторых отношениях слишком специализирована. Растения, вероятно, произошли от вымершего представителя Сharophyсеае, о чем говорит их близкое сходство с ними.
Первая группа, которую мы будем рассматривать в связи с происхождением растений, — порядок Соleochaetales, включающий нитчатые разветвленные, дисковидные и одноклеточные формы. Род Соleoc сhaete видами, произрастающими на поверхности погруженных пресноводных растений, имеет особенно сложное строение (рис. 15-17, 15-20). Эти водоросли при бесполом размножении образуют зооспоры; их зиготы покрыты слоем стерильных клеток, образующимся после оплодотворения. Отдельные особи Соleochaete могут состоять из массы дихотомически ветвящихся нитей или иметь дисковидную форму. Вегетативные клетки — одноядерные, с одним крупным хлоропластом и одним или более пиреноидами. Клеточное деление может идти на апексах нитей или по краям диска в зависимости от жизненной формы.
Рис. 15-20. A. Соeochaе со стебля водного цветкового растения, обитающего на озерном мелководье. Б. Особи этого вида Соleochaе представляют собой паренхиматозный диск толщиной обычно в один слой клеток. Крупные клетки — зиготы — защищены другими клетками. От диска отходят щетинковидные клетки с чехлом у основания. Соleochaе означает «щетинка в ножнах». Полагают, что щетинки защищают водоросль от поедания водными животными

Этот род считается близким к возможному предку высших растений. Некоторые его виды паренхиматозные и растут с участием краевой меристемы диска; размножение оогамное, зиготы защищены слоем клеток. Как и некоторые другие Сharophусеае, род Соleochaeе сходен с растениями также присутствием фрагмопласта, фото дыхательного фермента гликолатоксидазы в пероксисомах, а также многослойной структурой, связанной с жгутиками у репродуктивных клеток (см. рис. 15-17). Кроме того, обнаружены ископаемые формы, существовавшие примерно в эпоху возникновения первых растений и, по-видимому, напоминающие Coleochaete (рис. 15-21). Хотя сам этот род, вероятно, не является предком растений, его признаки, включая ряд структурных и биохимических адаптаций, полезных для жизни на суше, позволяют проследить путь, по которому могла идти эволюция земной флоры.
Рис. 15-21. Parka decipiens — ископаемая девонская водоросль возрастом около 380 млн. лет — сильно напоминает Coleochaete по форме, структуре и химии. У этого организма обнаружены многие признаки, которые позволяют предполагать его родственные связи с группой водных протистов, от которых возникли бриофиты и сосудистые растения

Порядок харовых (Charales) — вторая группа похожих на растения Сharophусеае (рис. 15-22). Эти своеобразные зеленые водоросли обитают в пресной или солоноватой воде. Некоторые из них имеют сильно обызвествленные клеточные стенки, поэтому хорошо сохранились в ископаемом виде. Сейчас существует около 250 видов харовых. Строение их довольно сложно: рост апикальный, как у растений; тело дифференцировано на узлы и междоузлия. В узлах возникают мутовки коротких ветвей, а междоузлия ценоцитные. Спермии харовых образуются в многоклеточных антеридиях, более сложных по строению, чем у прочих групп протистов, а яйцеклетки — в оогониях, устроенных проще антеридиев. Спермии — единственные жгутиковые клетки в жизненном цикле харовых.
Рис. 15-22. A. Chara, харовая водоросль, произрастающая на мелководье в озерах умеренной зоны. Четко виден ее своеобразный тип роста. Б. Ее сегмент с гаметангиями: сверху — оогоний, снизу — антеридий
Класс Ulvophусеае
Этот класс — единственный среди зеленых водорослей, который содержит в основном морские формы. Их жгутиковые клетки могут быть чешуйчатыми или «голыми», как у некоторых Сharophусеае, но они почти радиально симметричны и несут верхушечные, направленные вперед жгутики, как у Сhlorophусеае. Как и у последних, у Ulvophусеае, их может быть два, четыре или много, а у Сharophусеае только два. Что касается деталей клеточного деления, то у Ulvophусеае, — закрытый митоз с сохраняющейся ядерной оболочкой; веретено деления также не исчезает при цитокинезе. Это единственная группа зеленых водорослей с чередованием поколений и мейозом на стадии спор; в отличие от двух других подробно разбираемых здесь классов Ulvophусеае редко образуют покоящиеся зигоспоры.
Представители Ulvophусеае могут состоять из нескольких клеток, иметь форму нитей или плоских клеточных слоев, быть паренхиматозными или ценоцитными. Все нитчатые или более сложные формы морских зеленых водорослей имеют признаки этого класса, возникшего, вероятно, в отличие от других классов рассматриваемого отдела в море.
Первая эволюционная ветвь Ulvophусеае включает нитчатые формы с крупными многоядерными клетками, разделенными между собой септами. К ним относится род Cladophora(рис. 15-23), широко распространенный в пресной и морской воде. У его представителей нити растут в виде плотных скоплений, которые либо свободно плавают, либо прикреплены к камням или растениям; нити удлиненные и ветвятся ближе к концам. Каждая клетка содержит единственный периферический сетчатый хлоропласт с большим числом пиреноидов. У морских видов рода чередуются изоморфные поколения, а у пресноводных чередование отсутствует — по-видимому, утеряно в ходе эволюции.
Рис. 15-23. Cladophora из класса Ulvophyсeае — широко распространенная водоросль, обитающая в морской и пресной'воде. У морских видов происходит чередование поколений, как у большинства Ulvophусеае, а у пресноводных его нет. А. Ветвящиеся нити Cladophora. Б. Участок ветвящейся нити. В. Часть клетки с сетчатым хлоропластом. Г. Начало ветвления в апикальной части клетки. Д. Отдельная особь Сladophога, растущая в медленно текущем ручье в Калифорнии

Другой представитель этой ветви — Ulothrix, водоросль, обычная в холодных ручьях, прудах и озерах. Ее нити прикреплены к камням или другим субстратам с помощью особой структуры (рис. 15-24). Все клетки нитей в основном одинакового строения и содержат один кольцеобразный хлоропласт, пиреноид и ядро. Бесполое размножение происходит путем образования четырехжгутиковых зооспор, а половое — с участием изогамет с двумя жгутиками. Мейоз завершается перед прорастанием зиготы, поэтому нити Ulothrix гаплоидны. Хотя несколько групп зеленых водорослей, ранее относимых к видам Ulothrix, на самом деле являются представителями СЫогорЬусеае (см. рис. 15-37), сам этот род входит в класс Ulvophyceae.
Рис. 15-24. Ulothrix — представитель класса Ulvophyceae с не разветвленной нитчатой структурой. Нить слева с плотным содержимым клеток состоит из спорангиев, в которых формируются зооспоры. Другие нити — вегетативные. На правой нити виден прикрепительный аппарат

Род ульва (Ulva), известный под названием «морской салат», характеризуется иным типом роста. Эти обычные водоросли встречаются вдоль морских побережий всех умеренных областей земного шара (рис. 15-25). Отдельные особи ульвы имеют блестящий плоский таллом, или слоевище (просто организованное, относительно недифференцированное вегетативное тело), толщиной в два слоя клеток, а длиной в исключительных случаях до метра и более. Таллом прикрепляется к субстрату выростами базальных клеток. Каждая его клетка содержит одно ядро и хлоропласт. Ульва анизогамна; как большинство Ulvophyceae, имеет чередование изоморфных поколений (рис. 15-26).
Рис. 15-25. «Морской салат» Ulva — широко распространенный представитель класса Ulvophусеае, растущий по камням, сваям и сходным субстратам на морском мелководье всего земного шара


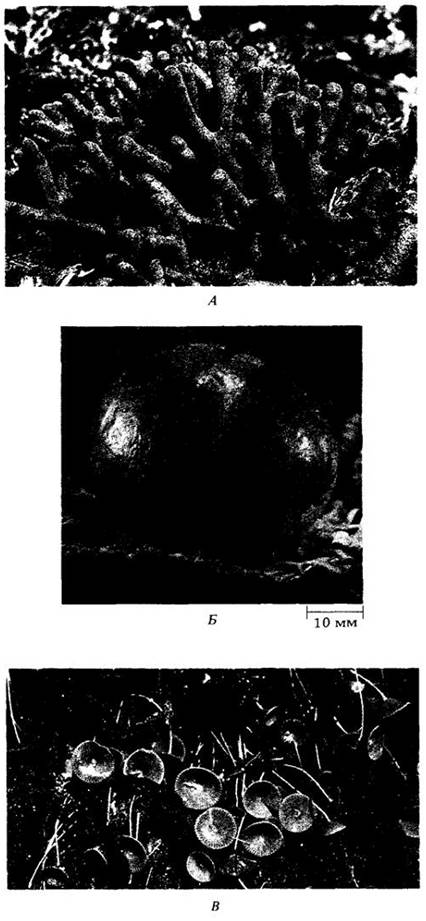
Водоросли морского порядка сифоновых (Siphonales) характеризуются очень крупными ветвистыми ценоцитными клетками, которые редко септированы и появляются в результате многократного ядерного деления без образования клеточных стенок (рис. 15-27). К этому порядку относится уже упоминавшийся вид Codium magnum. Клеточные стенки возникают у сифоновых только на стадии размножения. Одна из этих водорослей — Valonia, обычная в тропических водах, — широко использовалась в исследованиях этих стенок и в физиологических экспериментах, требующих большого объема клеточного сока. Valonia кажется одноклеточной, а на самом деле это крупный многоядерный пузырек с ризоидами и молодыми веточками (рис. 15-27, Б), дорастающий до размеров куриного яйца. Другой хорошо известный представитель сифоновых — ацетабулярия (Acetabularia), широко использовавшаяся в экспериментах по генетической регуляции дифференцировки. Ядра сифоновых, как правило, диплоидные; гаметы — единственные гаплоидные клетки в жизненном цикле.
Рис. 15-27. Три рода сифоновых водорослей (класс Ulvophyceae. А. Один из видов Codium, обильный вдоль атлантического побережья. Б. Vallonia обычно встречается в тропических водах, особи часто близки по размеру куриному яйцу. В. Acetabularia — «винный бокал для русалки» — водоросль в форме шляпочного гриба. Сифоновые водоросли на заднем плане — Dasycladus; фотография сделана на Багамских островах
Хлоропласты некоторых представителей этих водорослей (включая Codium) становятся симбионтами в теле морских голожаберных моллюсков, лишенных раковин. Эти животные поедают водоросли, хлоропласты которых сохраняются и делятся в клетках дыхательной полости. На свету эти хлоропласты фотосинтезируют столь эффективно, что по некоторым данным у моллюска Placobranchus ocettatus кислород может образовываться быстрее, чем потребляться в процессе дыхания животного.
Недавно было обнаружено, что один род сифоновых, Наlimeda (рис. 15-28), содержит вторичный метаболит, значительно снижающий выедание водоросли растительноядными рыбами и, по-видимому, служащий своеобразной химической защитой этих широко распространенных тропических протестов, часто являющихся самыми многочисленными фототрофными организмами рифовых сообществ, в которых они обитают. Такие токсичные вещества, вероятно, широко распространены среди водорослей и будут открываться с каждым годом все чаще.
Рис. 15-28. Halimeda — сифоновая водоросль, часто господствующая на рифах в теплых водах всего земного шара. Этот протист образует неприятные на вкус вещества, отпугивающие рыб и других морских растительноядных

Класс Сhlorophусеае
Большинство зеленых водорослей принадлежит к этой разнообразной группе, для которой характерно деление клетки с участием фикопласта. Этот уникальный признак указывает на то, что от представителей данного класса не возникла ни одна известная группа организмов. К Сhlorophусеае относятся жгутиковые и лишенные жгутиков одноклеточные водоросли, формы из нескольких клеток, подвижные или неподвижные колонии, нитчатые и паренхиматозные виды, обитающие в основном в пресной воде, хотя несколько одноклеточных планктонных видов встречается в прибрежных зонах морей. Некоторые представители класса ведут в основном сухопутный образ жизни и растут в почве или на древесине.
Подвижные одноклеточные Сhlorophусеае
К наиболее простым формам Сhlorophусеае относятся одноклеточные двужгутиковые организмы из порядка Сhlamydomonadales. Лучше всего изучен из них род хламидомонада (Chlamydomonas), включающий обычнейшие в пресных водах зеленые водоросли. Отдельные особи мелкие (как правило, меньше 25 мкм в длину), имеют зеленую окраску и округлую или грушевидную форму (рис. 15-29). Они быстро перемещаются специфическими резкими толчками, обусловленными биением двух гладких жгутиков, отходящих от более узкого переднего конца клетки. Двигаясь в противоположных направлениях, они продвигают водоросль сквозь толщу воды.
Каждая клетка хламидомонады имеет крупный массивный хлоропласт, содержащий красное пигментное тельце, стигму («глазок»), возможно, играющее роль экранирующего устройства, связанного со светочувствительной зоной. Недавние исследования показали, что фоторецептор хламидомонады гомологичен родопсину, зрительному пигменту многоклеточных животных. Предполагают, хотя это пока не доказано, что такой же зрительный пигмент содержится в «глазке» динофлагеллят, что свидетельствует об очень древнем его происхождении. Особи хламидомонады могут двигаться на свет определенной интенсивности. Недавно высказано мнение, что они также способны ориентироваться в магнитном поле, но это еще нуждается в подтверждении, и механизм такой ориентации неясен.
Рис. 15-29. Chlamydomonas — одноклеточная зеленая водоросль. На этой электронной микрофотографии можно видеть только основания жгутиков
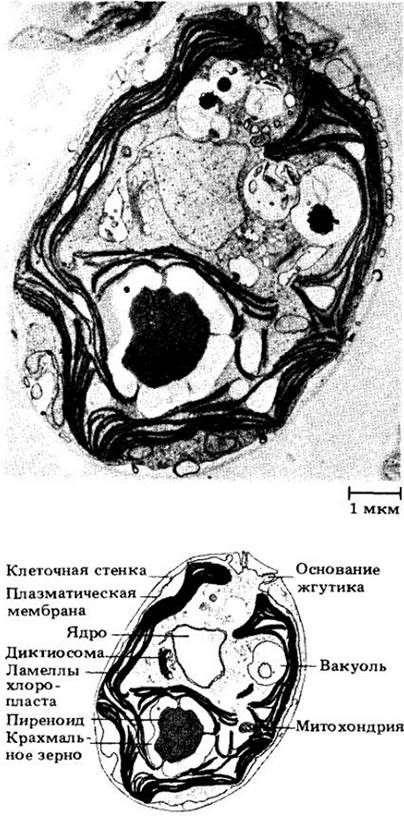
Хлоропласт хламидомонады содержит пиреноид близкой к округлой формы. Одноядерный протопласт окружен плазматической мембраной, снаружи от которой находится тонкая гликопротеиновая оболочка, богатая гидроксипролином. Целлюлоза в клеточной стенке отсутствует. На переднем конце клетки расположены две сократительные вакуоли, собирающие избыток воды и выделяющие ее наружу.
В определенных внешних условиях клетки хламидомонады становятся неподвижными. При этом они обычно теряют жгутики, а их стенка желатинизируется. Когда условия меняются, жгутики могут снова появиться, и клетка опять становится свободноплавающей.
Хламидомонада размножается бесполым и половым путями. В первом случае гаплоидное ядро обычно делится митотически с образованием под родительской клеточной стенкой четырех дочерних клеток. Каждая из них затем выделяет вокруг себя оболочку и образует жгутики. Эти клетки секретируют фермент, разрушающий материнскую оболочку, после чего они могут выйти наружу, хотя, даже полностью сформировавшись, часто остаются некоторое время внутри нее. У древних жгутиковых такие скопления дочерних клеток могли быть предшественниками колониальных организмов.
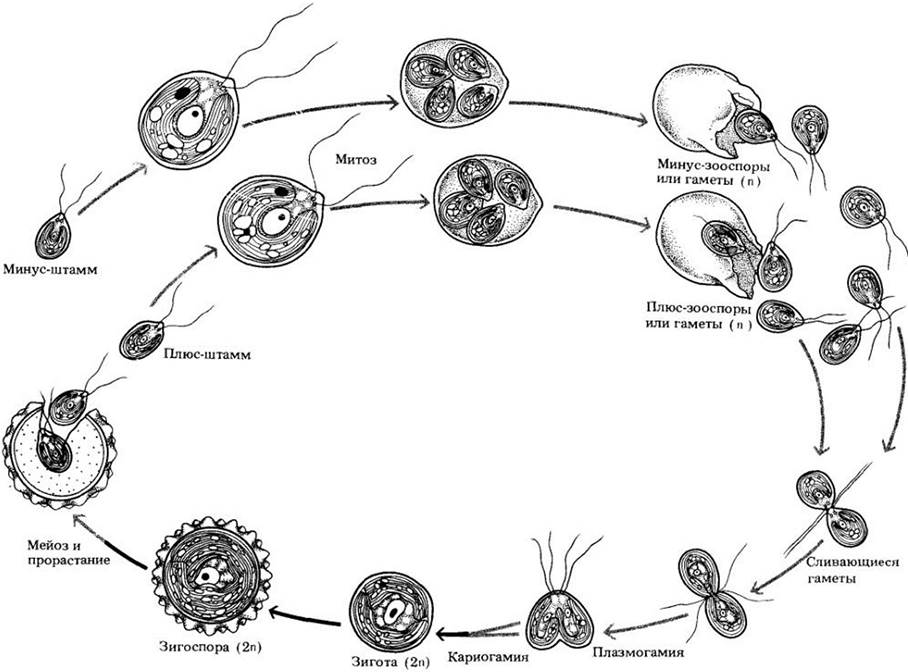
Половое размножение, известное у некоторых видов хламидомонады, осуществляется путем слияния особей, принадлежащих к разным типам спаривания (рис. 15-30). У вегетативных клеток азотное голодание стимулирует образование гамет, напоминающих эти клетки и скапливающихся в группы. Внутри этих скоплений формируются пары, в которых гаметы соединены сначала жгутиковыми мембранами, а затем тонким протоплазматическим тяжем у основания жгутиков. После образования этого протоплазматического мостика жгутики разъединяются, и одна или обе их пары начинают бить, перемещая частично слившиеся гаметы в воде. Затем происходит полное слияние. Вскоре после этого жгутики укорачиваются и в конечном итоге вообще исчезают, а вокруг диплоидной зиготы возникает толстая стенка. Толстостенная устойчивая зигота (зигоспора) проходит стадию покоя, в конце которой происходит мейоз с образованием четырех гаплоидных клеток с двумя жгутиками и собственной клеточной стенкой каждая. Эти клетки могут либо бесполо размножаться простым делением, либо сливаться с клетками другого штамма спаривания, давая новую зиготу. Таким образом, у хламидомонады мейоз зиготический (см. рис. 10-11, A) и в жизненном цикле преобладает гаплоидная фаза.
Рис. 15-30. Жизненный цикл хламидомонады. Половое размножение происходит, когда гаметы разных типов спаривания сближаются, слипаясь сначала своими жгутиковыми мембранами, а затем образуя тонкий протоплаз матический тяж — конъюгационный мостик. Протопласты обеих клеток полностью сливаются (плазмогамия), после чего объединяются их ядра (кариогамия). Затем вокруг диплоидной зиготы образуется толстая стенка. После периода покоя происходит мейоз и формируются четыре гаплоидные особи. Их бесполое размножение чаще всего идет путем простого деления
У многих видов хламидомонады клетки двух различных типов спаривания (условно обозначаемые значками «+» и «-») изогамны, т. е. одинаковы по размеру и строению. Помимо изогамных видов в этом роде имеются анизогамные (женская гамета еще подвижная, но крупнее мужской) и оогамные (с неподвижной женской гаметой) (рис. 15-31). Таким образом, в пределах одного рода встречаются все типы различий между гаметами, известные у водорослей.
Рис. 15-31. Типы полового размножения в зависимости от формы гамет; каждый из них обнаружен по крайней мере у одного вида хламидомонады. При изогамии гаметы одинаковы по размеру и форме. При анизогамии одна гамета, условно называемая мужской, мельче второй. При оогамии женская гамета неподвижна

В прошлом хламидомонаду считали примитивной формой рассматриваемого отдела, однако этот род имеет много продвинутых признаков, характерных для других Chlorophyсеае, и не похож в деталях на предка всех зеленых водорослей. Хламидомонадоподобные клетки напоминают некоторых представителей специализированных эволюционных ветвей этого класса, обсуждаемых ниже.
Неподвижные одноклеточные Chlorophyceae
Хлорелла (Chlorella) — одноклеточная зеленая водоросль без жгутиков, глазков и сократительных вакуолей; имеет округлую форму и по размерам меньше хламидомонады (рис. 15-32). В природе хлорелла широко распространена в пресной и соленой воде, а также почве. Каждая клетка содержит чашевидный хлоропласт с пиреноидом или без него и мелкое ядро. Единственный известный способ размножения — бесполый, причем каждая гаплоидная клетка митотически делится дважды или трижды с образованием соответственно четырех или восьми потомков.
Рис. 15-32. Chlorococcum echinozygotum — близкий родственник хлореллы, обычнейшая почвенная водоросль. А. Две особи. Б. Образование спор бесполого размножения в результате митоза внутри клетки
Хлорелла, первая выращиваемая в культуре водоросль, широко использовалась при изучении некоторых основных этапов фотосинтеза. Легкость культивирования делает ее идеальным экспериментальным организмом. Сейчас исследуется возможность использования хлореллы как источника питания для человечества (рис. 15-33). Опытные установки по ее выращиванию были созданы в США, ФРГ, Японии и Израиле. Японцы научились перерабатывать эту водоросль в безвкусный белый порошок, богатый витаминами и белком. Его можно смешивать с мукой при изготовлении хлебобулочных изделий. Недавно изучена возможность применения хлореллы для получения энергии; в этих экспериментах она выращивается вместе с бактерией, превращающей синтезируемый водорослью крахмал в липиды. Такие системы можно использовать на баржах или платформах в открытом океане или даже в космосе, что объясняет современную заинтересованность в их разработке. Другие водоросли, образующие большие количества углеводородов, также изучаются с точки зрения возможного промышленного применения.
Рис. 15-33. Эти бассейны под открытым небом в Седе-Бокер (Израиль) используются для проведения опытов, связанных с возможным промышленным получением белка из хлореллы. Хотя стоимость белка при этом выше, чем при его производстве из сои, что частично связано с неравномерным освещением воды в бассейнах, тем не менее эксперименты продолжаются и, как считают специалисты, вполне могут увенчаться успехом

Неподвижные колониальные Сhlorophyceае
Hydrodictyon, «водяная сеточка», — неподвижный колониальный представитель этого класса (рис. 15-34). При благоприятных условиях эта водоросль образует крупные скопления в прудах, озерах и медленно текущих ручьях. Каждая колония состоит из многих удлиненных клеток, расположенных в виде крупного полого цилиндра. Клетки сначала одноядерные, а затем становятся многоядерными. В зрелом состоянии каждая из них имеет обширную центральную вакуоль и пристенную цитоплазму, где располагаются ядра и крупный сетчатый хлоропласт с многочисленными пиреноидами. Бесполым путем Hydrodictyon размножается, образуя одноядерные двужгутиковые зооспоры. В конечном итоге зооспоры собираются в группы по четыре—девять (чаще всего по шесть) внутри цилиндрической материнской клетки, теряют жгутики и формируют дочерние колонии. Половой процесс у этого рода изогамный, с зиготическим мейозом, как у всех представителей Сhlorophусеае с половым размножением.
Рис. 15-34. А. «Водяная сеточка», Hydrodictyon — представитель колониальных Сhlorophyceae. Б. Фотография в сканирующем электронном микроскопе сплющенной молодой колонии Hydrodictyon reticulatum

Подвижные колониальные Сhlorophусеае
У подвижных колониальных родов этого класса хламидомонадоподобные формы объединены в колонии, передвигающиеся за счет биения жгутиков отдельных клеток (рис. 15- 35). Представителей этой группы часто называют вольвоксовыми — по роду вольвокс (Volvox), самому крупному и сложному из них колониальному организму (рис. 15-36). В колониях некоторых водорослей такого типа клетки связаны друг с другом цитоплазматическими мостиками, обеспечивающими интеграцию организма как единого целого. Наиболее просто устроенный представитель этой группы — гониум (Gonium, рис. 15-35, А, Б). Его колония состоит из отдельных клеток, которые удерживаются вместе студенистым матриксом. Каждая колония построена из 4, 8,16 и 32 клеток (в зависимости от вида), образующих слегка изогнутый щитовидный диск. Жгутики клеток бьют по-отдельности, двигая всю колонию вперед. Каждая клетка может делиться, образуя новую колонию.
Рис. 15-35. Несколько колониальных Сhlorophусеае. А, Б — С отит в двух ракурсах. В — Pandorina, Г — Eudorina. Клетки этих водорослей похожи на хламидомонаду; они соединены слизистым матриксом в многоклеточные колонии, которые передвигаются за счет биения жгутиков отдельных клеток. У различных родов уровень специализации клеток неодинаков

Близкородственный организм пандорина (Pandorina) представляет собой плотно упакованную яйцевидную или эллипсоидальную колонию из 16 или 32 клеток, собранных внутри матрикса (рис. 15-35, В). Колония полярна, причем на одном ее конце глазки клеток крупнее. Каждая клетка несет два жгутика, а поскольку все они обращены наружу, пандорина вращается в воде, как мяч. Когда клетки достигают максимального размера, колония опускается на дно, где каждая клетка делится, образуя дочернюю колонию. Последние остаются вместе, пока у всех не разовьются жгутики. Затем материнский матрикс вскрывается, подобно ящику Пандоры (отсюда и название водоросли), выпуская в воду новые организмы.
Eudorina — сферическая колониальная форма из 32, 64 или 128 (у различных видов) зеленых жгутиковых клеток (рис. 15-35, Г). Она отличается от гониума и пандорины тем, что некоторые меньшие других клетки на переднем, относительно направления движения ее конце не могут размножаться с образованием новых колоний. Таким образом, здесь началась функциональная специализация клеток.
Однако самый интересный представитель этих колониальных зеленых водорослей — вольвокс (рис. 15-36). Он представляет собой полую сферу, образованную одним слоем из 500— 60 000 (у разных видов) двужгутиковых вегетативных и небольшого числа репродуктивных клеток. Когда вольвокс, кружась, перемещается в воде, он кажется вращающейся Вселенной из множества звезд, закрепленных на невидимой небесной тверди.
Рис. 15-36. Бесполое рамножение Volvox carteri. На периферии прозрачной сферы находится около 2000 мелких хламидомонадоподобных соматических клеток; в зрелой колонии этого вида они не связаны между собой, как у некоторых других представителей рода. У приводимого здесь организма 16 репродуктивных клеток внутри сферы уже поделились, образовав молодые сферы с примерно 2000 крошечных соматических клеток и 16 репродуктивными клетками. Затем каждая дочерняя колония «проест» выход из материнской, выйдет наружу и цикл повторится

Колонии этого вида полярны, т. е. у них различаются передний и задний полюса. Жгутики каждой клетки бьют таким образом, что весь организм, продвигаясь вперед
(обычно к свету на большинстве стадий жизненного цикла), вращается вокруг своей оси по часовой стрелке.
Большинство клеток этой сферической колонии чисто вегетативные, а немногие связанные с бесполым размножением, как правило, особым образом расположены в заднем полушарии. У некоторых видов вольвокса клетки бесполого размножения не вполне различимы в молодых сферах, но позднее начинают выделяться, проходя последовательные циклы роста и деления. В конце концов эти делящиеся клетки образуют дочерние сферы, которые «вылупляются» из родительской, выделяя особый фермент, растворяющий ее прозрачный матрикс. У более продвинутых представителей рода такие репродуктивные клетки обособляются в каждом цикле размножения очень рано и все время структурно и функционально отличаются от вегетативных, или соматических (рис. 15-36). У этих видов, например, у Volvox carteri (рис. 15-36), наблюдается настоящее «разделение труда» между двумя взаимозависимыми типами клеток: подвижные соматические выносят неподвижные репродуктивные к поверхности воды, где достаточно света и СО2 для фотосинтеза. Таким образом, наиболее продвинутые представители Volvox уже не просто колониальные, а многоклеточные дифференцированные организмы, как растения и животные.
Половое размножение у вольвокса всегда оогамное, хотя существенно варьирует в деталях у разных видов. В одних случаях яйцеклетки и спермин могут возникать в пределах одной сферы, в других представители генетически однородного клона (происшедшие бесполым путем от одной особи) становятся мужскими и женскими колониями, а обоеполые сферы неизвестны. Однако у более продвинутых видов пол генетически детерминирован, и каждый возникший бесполым путем клон является либо мужским, либо женским. У всех изученных видов половое размножение в популяции колоний синхронизировано с помощью химических половых индукторов. Эти молекулы синтезируются сферой, достигшей «половой зрелости» за счет какого-то еще не вполне ясного механизма. Одна мужская колония V. carteri может выделить количество индуктора, достаточное для перехода к половому размножению более 0,5 млрд, других колоний этого вида.
Вольвокс — один из наиболее просто организованных многоклеточных организмов с четким «разделением труда» между клетками. Половой индуктор этой водоросли относится к наиболее сильным из известных в настоящее время биологически активным веществам. Поэтому с данным родом (особенно с У. carteri) в последние годы связано большое количество исследований. Подобно другим родственным водорослям, вольвокс гаплоиден, т. е. мутантные гены у него не маскируются доминантными аллелями и влияющие на развитие мутации могут быть легко обнаружены. Уже выделены сотни штаммов со специфическими наследственными дефектами, на которых исследуется регуляция клеточной дифференцировки специфическими генами.
Подводя итоги, необходимо отметить, что увеличение специализации у колониальных Chlorophyceae прослеживается по нескольким направлениям. Во-первых, происходит увеличение числа клеток и размеров колоний. Во-вторых, идет морфофункциональная специализация клеток. Наконец, возрастает половая специализация, параллельная отмеченной в роде Chlamydomonas. Так, G onium и Pandorina изогамны, a Eudorina и Volvox оогамны. Тем не менее эта ветвь эволюции явно тупиковая, поскольку не дала начала более сложно организованной группе организмов.
Нитчатые и паренхиматозные Chlorophyceae
Представители порядков Chaetophoraies и Oedogoniales имеют наиболее сложное строение среди водорослей данного класса. Их клетки часто специализированы с точки зрения выполняемых функций или положения в теле протиста и соединены между собой плазмодесмами, как у растений. Хотя многие роды этих групп при делении клеток образуют, как и растения, клеточную пластинку (рис. 15-37), они имеют также фикопласт, как все Chlorophyceae, поэтому не могут считаться предками растений. Кроме того, как и у всех Chlorophyceae, в течение митотического цикла сохраняется ядерная оболочка.
Рис. 15-37. Stigeocloniшn — представитель класса Сhlorophусеае, относимый раньше к роду Ulothrix (Ulvophусеае). Деление клетки у Stigeoclonium происходит с образованием клеточной пластинки, которая почти полностью сформировалась на стадии цитокинеза, показанной на этой электронной микрофотографии. Ядра находятся рядом, так как веретено деления исчезло на стадии телофазы прошедшего митоза. Фикопласт не заметен из-за относительно малого увеличения

Среди нитчатых представителей этого класса Stigeoclonium (рис. 15-37) внешне напоминает Ulothrix (рис. 15-24), но имеет все признаки Chlorophyceae. Эти два рода вместе с другими Chlorophyceae объединяли в один, но теперь детальные исследования с помощью электронной микроскопии показали их принадлежность к разным классам. У Stigeoclonium в отличие от Ulothrix разветвленные нити.
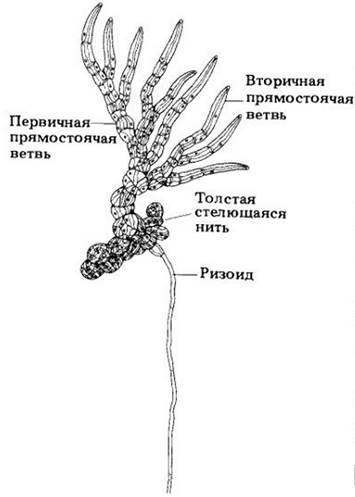
Fritschiella — это наземная водоросль, по большинству микроскопических признаков сходная с Stigeoclonium, но более сложная по строению. Ее тело состоит из подземных ризоидов, паренхиматозных стелящихся тяжей вблизи поверхности почвы и двух типов прямостоячих выступающих из нее веточек (рис. 15-38). Fritschiella обитает также на влажных поверхностях — сырых стенах, стволах деревьев, листьях. Она, как и растения, приспособилась к наземному образу жизни и имеет некоторые признаки, вероятно, присущие их предковым формам.
Рис. 15-38. Fritschiella — сухопутная водоросль из класса Сhlorophусеае. В связи с адаптацией к наземному образу жизни у нее в процессе эволюции независимо появились признаки, характерные для растений
Oedogonium — неразветвленная нитчатая водоросль, прикрепленная к субстрату особым приспособлением. У нее весьма своеобразные признаки, особенно в отношении клеточного деления. Клетки этого рода одноядерные с периферическим сетчатым хлоропластом. Когда клетка делится, то у верхнего (апикального) ее конца из материала клеточной стенки образуется кольцо в форме бублика. После деления ядра одно из дочерних ядер мигрирует к верхнему концу клетки, стенка которой резко разрывается точно у этого кольца. Затем оно вытягивается в цилиндр, образуя стенку верхней дочерней клетки, расположенной ближе к апексу нити. Новая поперечная перегородка формируется в плоскости фикопласта сразу же за краем разорванной родительской стенки. В результате удлинения кольца края исходной материнской клетки отгибаются наружу, образуя характерные рубцы (рис. 15-39), указывающие на число происшедших ранее делений.
Рис. 15-39. Oedogonium — неразветвленная нитчатая водоросль из класса Сhlorophусеае. Участок вегетативной нити с рубцами

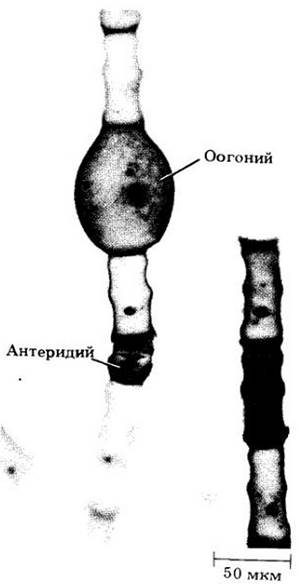
Бесполое размножение Oedogonium происходит зооспорами, образующимися по одной на клетку. Каждая зооспора имеет венчик из примерно 120 жгутиков. Половое размножение — оогамия (рис. 15-40). Каждый антеридий образует два многожгутиковых спермия, а каждый оогоний — одну яйцеклетку. Мейоз зиготический, как и у всех Chlorophyceae.
Рис. 15-40. Половое размножение у Oedogonium оогамное. Каждый оогоний образует одну яйцеклетку, а каждый антеридий — два многожгутиковых спермия