СОВРЕМЕННАЯ БОТАНИКА - П. РЕЙВН - 1990
РАЗДЕЛ IV. РАЗНООБРАЗИЕ
ГЛАВА 16. МОХОВИДНЫЕ
Данные, приведенные в предыдущей главе, свидетельствуют о том, что растения (моховидные и сосудистые) произошли от каких-то древних групп зеленых водорослей. Как и у последних, у растений главный фотосинтетический пигмент — хлорофилл а, а вспомогательные — хлорофилл b и каротиноиды. У всех трех групп основной запасной углевод — крахмал, который откладывается в хлоропластах, а не в цитоплазме, как у других фотосинтезирующих эукариот. Целлюлоза является важнейшим компонентом клеточной стенки и у растений, и у некоторых зеленых водорослей. Наконец, растения при клеточном делении образуют фрагмопласт и клеточную пластинку. Среди других живых организмов эту особенность имеют только один род бурых и несколько родов зеленых водорослей. Так как все указанные признаки наблюдаются и у моховидных (рис. 16-1), и у сосудистых растений, можно допустить, что обе группы произошли от отдаленного общего предка, успешно завоевавшего сушу.
Рис. 16-1. Густо разросшийся мох Fissidens на известняковых скалах водопада. Эта фотография была сделана в заповеднике к западу от Ялты на Крымском полуострове в СССР

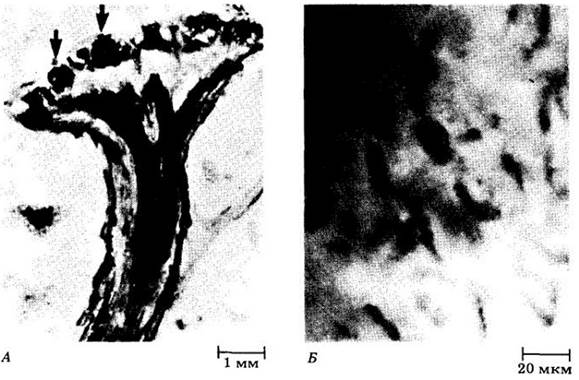
Если эта гипотеза верна, то эволюционные ветви моховидных (бриофитов) и сосудистых растений должны были разойтись очень давно. Древнейшие известные ископаемые, напоминающие моховидные, имеют девонский возраст, т. е. порядка 400 млн. лет (рис. 16-2), а остатки, безоговорочно относимые к бриофитам, — по крайней мере 370 млн. лет. Ископаемые, сходные с сосудистыми растениями, обнаружены в нижнесилурийских отложениях возрастом около 430 млн. лет (см. рис. 1-5, с. 14). В свете этих находок можно предположить, что гипотетический общий предок моховидных и сосудистых растений — относительно сложно устроенная зеленая водоросль (см. рис. 15-21) — заселил сушу более 430 млн. лет назад. Тетрады спор и кутикулоподобные фрагменты ордовикского возраста (около 450 млн. лет назад) наводят на мысль и о более раннем происхождении растений.
Рис. 16-2. А. Продольный срез гаметофита из кремнистых сланцев в окрестностях деревни Райни (Шотландия). Это растение жило в раннем девоне, около 400 млн. лет назад. Стрелки указывают положение антеридиев на верхней поверхности гаметофита. Б. На срезе антеридия видны зрелые мужские гаметы. По размеру и сложности строения такие гаметофиты похожи на гаметофиты моховидных. Неизвестно, родственны ли этим гаметофитам какие- либо раннедевонские спорофиты из этих же отложений
Так как у всех моховидных и у сосудистых растений наблюдается хорошо выраженное чередование поколений, это явление было свойственно, по-видимому, и их общему предку. Его гаметофиты почти наверняка формировали многоклеточные гаметангии, сходные с имеющимися у всех современных растений. У всех растений есть зародыш: зиготы начинают делиться внутри гаметофита и полностью зависят от него в смысле питания. У водорослей зародышей нет, и неизвестно, были ли они у первых растений. Мы не знаем также, имел ли предок растений проводящие ткани, а если да, то какие. Однако у некоторых нижнесилурийских ископаемых обнаружены удлиненные клетки, напоминающие трахеиды, с кольцевыми или спиральными утолщениями; некоторые данные говорят о наличии в их стенках лигниноподобных веществ. Не исключено, что первые наземные растения вступали в эндомикоризный симбиоз, возможно, с каким-то зигомицетом (см. с. 206).
Организмы, успешно переселившиеся из воды на сушу, развили структуры, предохраняющие их от высыхания. Одна из них — стерильный покровный слой вокруг формирующих соответственно спермии и яйцеклетки клеток мужского и женского гаметангиев, т. е. антеридия и архегония. Подобным же образом, стерильный слой сформировался вокруг спорообразующих клеток спорангиев.
Заселение суши предками растений сопровождалось задержкой зиготы внутри женского гаметангия и ее развитием здесь в зародыш. В результате на критических ранних стадиях развития молодой зародыш (спорофит) защищен женским гаметофитом. В противоположность этому у зеленых водорослей развитие зиготы, как правило, от женского гаметофита не зависит.
Надземные части большинства сосудистых растений покрыты восковым защитным слоем, кутикулой, отчасти предохраняющей от высыхания. Ее присутствие тесно коррелирует с устьицами, специализированными порами, главное назначение которых — регуляция газообмена. Кутикула, по- видимому, отсутствует у большинства бриофитов, но устьица имеются на спорофитах антоцеротовых и листостебельных мхов. Иногда у последних они состоят из единственной бубликовидной замыкающей клетки, т. е. совершенно не такие, как у сосудистых растений. У антоцеротовых устьица, вероятно, открыты до поздней стадии развития, а у мхов закрываются только после того, как спорофит полностью высохнет. Таким образом, устьица бриофитов не функционируют так, как у современных сосудистых растений.
Все растения оогамны и имеют чередование гетероморфных поколений.
Признаки моховидных
Моховидные — печеночники, антоцеротовые и мхи — относительно мелкие растения; длина многих из них менее 2 см, а большинства — менее 20 см (рис. 16-3). Часто они в изобилии произрастают в относительно влажных местах, где встречается большое число их видов с многочисленными особями. Мхи иногда господствуют, вытесняя прочие растения, на обширных пространствах приполярных областей и на каменистых склонах гор выше границы леса. Многие из них способны даже выдерживать длительные периоды сильных антарктических холодов (рис. 16-4). Подобно лишайникам, бриофиты весьма чувствительны к загрязнению воздуха (особенно двуокисью серы) и в сильно загрязненных областях часто отсутствуют или представлены лишь немногими видами. Ряд мхов встречается в пустынях, причем некоторые из них образуют обширные скопления на сухих открытых скалах, которые могут прогреваться до очень высоких температур. Многие мхи годами сохраняют жизнеспособность в сухом состоянии, быстро восстанавливая активность при увлажнении. Известны водные мхи и печеночники, погибающие в течение примерно суток при высыхании. Несколько бриофитов найдено даже на захлестываемых морскими волнами скалах, хотя ни один из них не является истинно морским обитателем. Насчитывается около 16 000 видов моховидных. Это больше, чем в любой другой группе растений, за исключением цветковых.
Рис. 16-3. Кукушкин лен (Polytrichum). Видны спорофиты, прикрепленные к женским гаметофитам. Каждый спорофит состоит из коробочки, ножки и стопы, соединяющей его с гаметофитом. Листья в нижней части фотографии — части гаметофитов, на которых образовались спорофиты после оплодотворения, происшедшего по крайней мере шестью месяцами раньше

Рис. 16-4. А. Гора Мельбурн в Антарктиде (примерно 75° ю. ш.). Здесь на высоте 3000 м над уровнем моря суточные температуры летом колеблются от — 10 до — 30 °С. В этих невероятно суровых условиях, на свободных от снега пространствах, заметных на фотографии, ботаники из Новой Зеландии открыли в 1984 г. картинки мха из рода Campylopus (Б), растущего там, где вулканическая активность прогревает субстрат до 30 °С. Произрастание Campylopus в этих местах демонстрирует скорее замечательную способность мхов к расселению, чем их способность выживать в суровых условиях, которая, впрочем, тоже высока

Два важных признака отличают бриофиты от сосудистых растений. Во-первых, отсутствие у них специализированных проводящих тканей — ксилемы и флоэмы. Это значит, что все моховидные, строго говоря, не имеют настоящих листьев, стеблей и корней, так как один из признаков перечисленных структур — присутствие в них именно таких тканей. Тем нс менее по отношению к листоподобным и стеблевидным структурам гаметофитов печеночников и мхов обычно используются термины «лист» и «стебель». Мы тоже будем следовать этой традиции.
Ножки, или спорофоры, спорофитов у большинства мхов содержат центральный тяж водопроводящих клеток, называемых гидроидами; сходные клетки имеются и в гаметофитах многих из этих видов. Гидроиды имеют удлиненную форму со скошенными на концах тонкими и легко проницаемыми для воды стенками, что делает их предпочтительными путями передвижения воды и растворов. Подобно трахеидам и сосудам сосудистых растений, зрелые гидроиды не имеют протопласта и кажутся пустыми. Однако они отличаются отсутствием специализированных утолщений стенки (рис. 20-13). У некоторых мхов тяж гидроидов окружен клетками,
проводящими питательные вещества, так называемыми лептоидами (рис. 16-5), что очень напоминает взаиморасположение флоэмы и ксилемы у некоторых сосудистых растений. Зрелые лептоиды имеют удлиненную форму, дегенерировавшие ядра и живой протопласт. Некоторые из них очень похожи на клетки флоэмы примитивных сосудистых растений. Не исключено, что проводящие воду и питательные вещества клетки мхов и сосудистых растений происходят от однотипных клеток, присутствовавших у общего предка этих двух групп.
Рис. 16-5. Проводящие тяжи в ножке спорофита мха Dawsonia superba. А. Поперечный срез, демонстрирующий общую структуру ножки (сканирующий электронный микроскоп). Б. Поперечный срез с центральным тяжем из гидроидов, округленным обкладкой из лептоидов и паренхимой коры. В. Продольный срез участка центрального тяжа. Слева направо: гидроиды, лептоиды, паренхима


У большинства моховидных гаметофит прикрепляется к субстрату вытянутыми одиночными клетками или нитями клеток, называемыми ризоидами. Как правило, они служат только для заякоривания растений, так как вода и неорганические ионы обычно непосредственно и быстро поглощаются всем гаметофитом. Корнеподобных органов нет, хотя структура подземных стеблей некоторых мхов довольно сложна.
Второй важный отличительный признак бриофитов — это характер чередования поколений: гаметофиты в отношении питания всегда независимы, тогда как спорофиты постоянно прикреплены к гаметофитам и в различной степени зависят от них. Другими словами, гаметофит моховидных — крупное доминирующее поколение, в то время как у сосудистых растений таким поколением является спорофит (см. рис. 10-11, В).
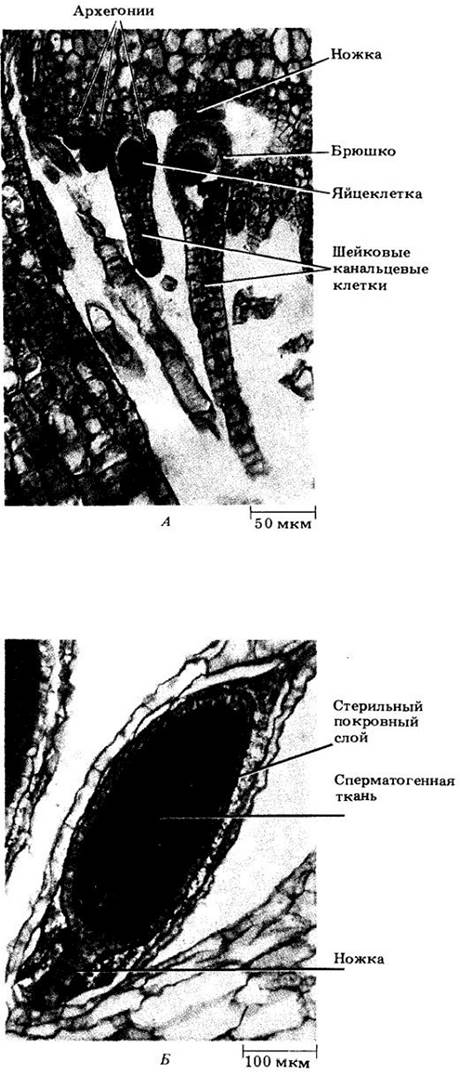
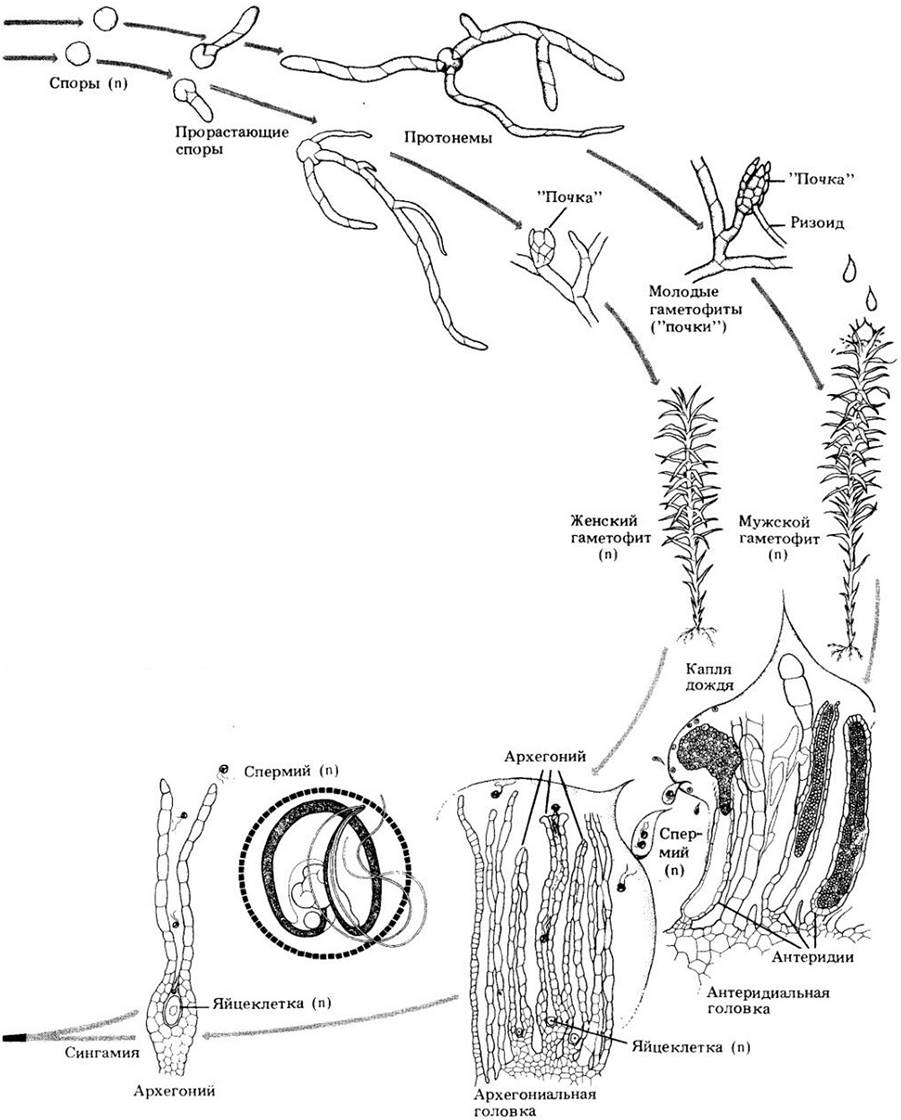
Моховидные населяют влажные (по крайней мере, в некоторые сезоны) местообитания. Чтобы произошло оплодотворение, двужгутиковый спермий должен подплыть в воде к яйцеклетке, находящейся внутри архегония. Последний имеет колбовидную форму, иногда ножку, длинную шейку и расширенную базальную часть — брюшко с яйцеклеткой (рис. 16-6, А). В центре шейки так называемые шейковые канальцевые клетки при созревании архегония разрушаются, оставляя заполненный жидкостью канал, по которому спермий плывет к яйцеклетке. Продолговатый или сферический антеридий (обычно на ножке) состоит из стерильной оболочки толщиной в одну клетку, окружающей многочисленные сперматогенные клетки (рис. 16-6,5). Из каждой такой клетки формируется один двужгутиковый спермий.
Рис. 16-6. Гаметангии маршанции. А. Несколько архегониев на разных стадиях развития. Б. Незрелый антеридий. У маршанции архегонии и антеридии образуются на разных гаметофитах
У печеночников и антоцеротовых спермии часто действительно плывут к архегонию по сплошной водной пленке. Приблизившись, они привлекаются в шейку архегония выделяемыми им химическими веществами. У мхов антеридии часто скучены и окружены листовидными структурами, образующими чашу (рис. 16-7). При этом спермии выходят в воду внутри такой чаши, а затем рассеиваются при попадании туда дождевых капель. Капельки воды с большим количеством спермиев могут переносить от растения к растению насекомые. Поскольку спермии такими способами могут достигать даже удаленных на относительно большие расстояния изолированных женских гаметофитов, последние часто образуют множество спорофитов.
Рис. 16-7. Листостебельные мужские гаметофиты мха Polytrichum piliferum, на которых видны собранные в головки зрелые антеридии. Спермии выходят в воду, скапливающуюся внутри этих окруженных листьями головок, и затем выплескиваются из них с каплями дождя, иногда попадая на другой гаметофит с архегониями

Оплодотворенная яйцеклетка, или зигота, остается в брюшке архегония и развивается там в зародыш. Некоторое время клетки брюшка делятся параллельно с ростом находящегося внутри молодого спорофита. Разросшийся архегоний образует так называемый колпачок (калиптру). Зрелый спорофит многих моховидных состоит из коробочки (спорангия) на ножке (спорофоре), переходящей в «стопу», закрепленную в архегонии (рис. 16-8).
Рис. 16-8. Почти зрелый спорофит маршанции. Заметны элатеры — спиральные нитчатые структуры внутри заполненной спорами коробочки, способствующие рассеиванию спор

Клетки молодых и созревающих спорофитов обычно содержат хлоропласты и фотосинтезируют, но к тому времени, когда в коробочке происходит мейоз и образуются споры, хлорофилл, как правило, исчезает. У большинства мхов калиптра по мере удлинения ножки выносится вверх месте с коробочкой; этот защитный колпачок отпадает перед рассеиванием спор, и они выбрасываются наружу при спонтанном растрескивании спорангия.
Моховидные традиционно разделяются на три класса Hepaticae (печеночники, 6000 видов), Anthocerotae (антоцеротовые, 100 видов) и Мusci (листостебельные мхи, 9500 видов). Эти три группы резко различаются между собой, а общие их признаки в общих чертах характерны для просто организованных растений. Большинство мхов имеют проводящую ткань и вполне могут считаться редуцированными сосудистыми растениями, с которыми у них, по-видимому, был общий предок, тогда как печеночники и антоцеротовые отделились от этой эволюционной линии раньше. Доказательств происхождения печеночников от предков с проводящей тканью, а также их близкого родства с мхами нет. Антоцеротовые сильно отличаются от обеих этих групп и далеки от любой известной группы организмов. Учитывая это, все большее число ботаников считает три группы моховидных разными отделами, сходство которых — не доказательство родства, а результат конвергентной эволюции или сохранения примитивных признаков. Однако в этой книге мы по традиции оставляем их в одном отделе.
Класс Hepaticae
Печеночники — это мелкие растения, обычно менее заметные, чем мхи. Их название возникло в IX в., когда считалось, что из-за напоминающих печень очертаний гаметофита у некоторых родов этих растений они могут быть полезны при лечении болезней этого органа (согласно средневековым медицинским представлениям, внешний вид растения говорит о его специфических целебных свойствах).
Гаметофиты некоторых печеночников — уплощенные в дорсовентральном направлении слоевища, или талломы (этот термин традиционно используется для обозначения недифференцированных тел растений, обычно с четко выраженными верхней и нижней поверхностями), нарастающие апикальной меристемой. Однако у большинства видов класса они облиственные. Рост связан с делением единственной апикальной клетки, похожей на перевернутую пирамиду с основанием и тремя боковыми гранями, отделяющими дочерние клетки. Ризоиды печеночников одноклеточные в отличие от мхов, у которых они состоят из нескольких клеток. Гаметофиты развиваются непосредственно из спор. Спорофиты большинства печеночников обычно проще, чем у мхов, а их коробочки имеют более разнообразные механизмы рассеивания спор.
Слоевищные печеночники
Слоевищные (необлиственные) печеночники весьма разнообразны. Они встречаются по влажным тенистым берегам рек и в других пригодных для них местах, таких, например, как цветочные горшки в неотапливаемых оранжереях. Слоевище образовано большим числом клеточных слоев (около 30 на срединном ребре и примерно 10 в более тонких местах) и четко дифференцировано на тонкую, богатую хлорофиллом верхнюю (дорсальную) и более толстую, бесцветную нижнюю (вентральную) части (рис. 16-9, А). Нижняя поверхность несет два типа ризоидов, а также ряды чешуек. Верхняя поверхность разделена на приподнятые участки, каждый из которых указывает размеры лежащей под ним воздушной камеры и имеет крупную ведущую в нее нору (рис. 16-9, Б).
Рис. 16-9. А. Поперечный разрез гаметофита слоевищного печеночника маршанции. Видны многочисленные несущие хлоропласты клетки верхних слоев, несколько слоев бесцветных клеток под ними и ризоиды, прикрепляющие тело растения к субстрату. Газообмен в воздушных камерах дорсального ассимиляционного слоя осуществляется через поры. Специализированные клетки, окружающие пору, обычно расположены друг над другом 4 — 5 «венцами» из 4 клеток каждый, образуя в результате бочонковидную структуру. В условиях сухости клетки самого нижнего «венца», обычно вдающиеся внутрь поры, смыкаются, а при увеличении важности расходятся. Таким образом поры выполняют функцию, сходную с функцией устьиц у сосудистых растений. Б. Микрофотография под сканирующим электронным микроскопом поры на дорсальной поверхности гаметофита маршанции
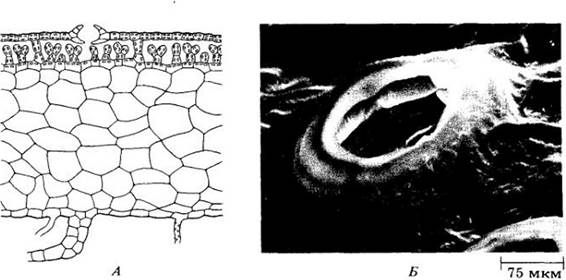
Рис. 16-10. Гаметофиты маршанции. Антеридии (А) и архегонии (Б) приподняты над слоевищем на ножках

Рис. 16-11. Зрелые споры и элитеры из коробочки маршанции
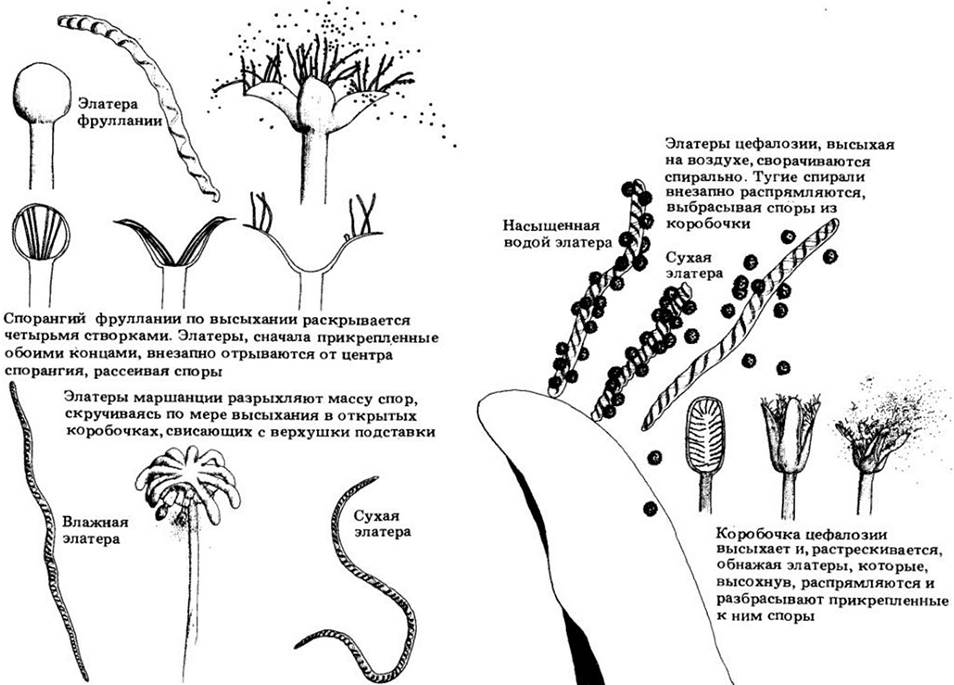
Один из наиболее известных печеночников — маршанция (Marchantía), широко распространенный наземный род, растущий на влажной почве и скалах (рис. 16-10). Дихотомически ветвящиеся гаметофиты достигают в длину большей частью от одного до нескольких сантиметров, а гаметангии сосредоточены на специализированных прямостоячих структурах, называемых гаметофорами (подставками). Гаметофиты маршанции однополые; мужские и женские особи легко распознаются по разной форме подставок. У антеридиев они имеют дисковидные головки и называются антеридиофорами, а у архегониев эти головки пальчатые, и подставки называются архегониофорами (рис. 16-10). Жизненный цикл маршанции изображен на рис. 16-13. У этого рода спорофитнос поколение состоит из стопы, короткой ножки и коробочки, или спорангия (см. рис. 16-8). Кроме спор зрелый спорангий содержит удлиненные клетки, элатеры, со спиральными гигроскопичными (поглощающими влагу) утолщениями стенок (рис. 16-8 и 16-11; см. также приложение «Рассеивание спор у печеночников»). Стенки этих клеток чувствительны к малейшим изменениям влажности, в ответ на которые скручиваются и раскручиваются. Это способствует рассеиванию спор после растрескивания коробочки на ряд лепестковидных сегментов.
Основной способ бесполого размножения печеночников — фрагментация таллома. Другой широко распространенный путь такого размножения у этого класса, а также у мхов — образование гемм (выводковых почек) — многоклеточных телец, которые могут давать начало новым гаметофитам. У маршанции геммы образуются в особых чашевидных структурах, выводковых корзинках, на дорсальной стороне гаметофита (рис. 16-12). Выводковые почки распространяются главным образом за счет вымывания их брызгами дождя.
Рис. 16-12. А. Гаметофиты марцианции с чашевидными выводковыми корзинками, содержащими выводковые почки, которые выплескиваются наружу дождем и могут дать начало новым гамета фитам. Каждый из них будет генетически идентичен родительскому растению, от которого путем митоза происходят выводковые почки. Б. Продольный срез выводковой корзинки

Облиственные печеночники
Эта неоднородная группа объединяет более 4000 из 6000 известных видов класса (рис. 16-14). Она особенно обильно представлена в тропиках и субтропиках, в районах с большим количеством осадков или высокой влажностью (рис. 16-15), но достаточно многочисленна и в умеренных областях. Эти растения обычно сильно ветвятся и образуют небольшие дернинки.
Листья печеночников, как и у мхов, обычно состоят всего из одного слоя недифференцированных клеток и у многих родов расположены двурядно с третьим рядом редуцированных листьев по нижней поверхности гаметофита. Листья часто двулопастные, причем каждый нарастает двумя апикальными точками. У Frullania, обычного накорового печеночника, листья состоят из крупной цельной дорсальной лопасти и мелкой шлемовидной вентральной (рис. 16-14, В).
У облиственных печеночников антеридии обычно находятся в сумковидном разрастании, андроцее, на нижней части видоизмененного листа. Развивающийся спорофит, как и архегоний, из которого он вырастает, имеет своеобразный трубчатый чехол, периантий (рис. 16-14, В).
Класс Аnthocerotae
Этот класс включает всего около 100 видов, объединяемых в шесть родов. Из них наиболее известен антоцерос (Аnthoceros), встречающийся по всему миру, обычно во влажных затененных местах. Гаметофиты антоцероса и слоевищного печеночника внешне сходны (рис. 16-16, А), но многие признаки свидетельствуют против близкого родства этих растений. Например, в первом случае каждая клетка, как у многих водорослей, обычно содержит единственный крупный хлоропласт, а не много мелких дисковидных хлоропластов, как у всех других растений. В каждом хлоропласте находится пиреноид, делающий сходство с водорослями еще более выраженным. Строение гаметофитов четко дорсовентральное, часто розетковидное с диаметром обычно 1 — 2 см. Крупные внутренние полости гаметофита антоцероса заполнены слизью, а не воздухом, как в случае слоевищных печеночников. В этих полостях со слизью обычно живут цианобактерии рода Nostoc, фиксирующие азот и снабжающие им своих растений-хозяев.
У некоторых видов антоцероса гаметофиты однополые, у других — обоеполые. Антеридии и архегонии погружены в ткань дорсальной поверхности гаметофита, причем антеридии располагаются группами в особых камерах. На одном растении может развиваться множество спорофитов.
Прямостоячий вытянутый спорофит антоцероса состоит из стопы и длинного цилиндрического спорангия (рис. 16-16, Б и 16-17, А). На очень ранней стадии развития между стопой и спорангием закладывается меристема, или зона активно делящихся клеток, функционирующая до тех пор, пока условия благоприятны для роста. В результате спорофит продолжает долгое время вытягиваться. Он зеленый, так как имеет несколько слоев фотосинтезирующих клеток, и покрыт кутикулой с устьицами (рис. 16-16, В). Созревание спор, а затем растрескивание спорангия начинаются от его верхушки и распространяются к основанию (рис. 16-16, Г, Д). Среди спор присутствуют стерильные вытянутые часто многоклеточные образования, напоминающие элатеры печеночников (рис. 16-17, Б). Растрескивается спорангий по всей длине на лентовидные половины.
Класс Musci
Во многих группах растений некоторые представители в народе неправильно называются мхами. Так, «олений мох» и «исландский мох» — лишайники, «бородатый мох» — сосудистое растение, «ирландский мох» — водоросль. Настоящие же мхи относятся к классу Мша, который состоит из трех подклассов: Вryidae (настоящие, или бриевые, мхи), Sphagnidae (сфагновые, или торфяные, мхи) и Аndreaeidae (андреевые мхи).
Настоящие мхи
Гаметофиты всех мхов представлены двумя различными фазами: протонемой (от греческих слов protos — первый и nema — нить), развивающейся непосредственно из прорастающей споры, и листостебельным гаметофитом. У настоящих мхов протонема состоит из одного слоя клеток, ветвится и напоминает нитчатую зеленую водоросль (рис. 16-18). Листостебельные гаметофиты развиваются из крошечных почковидных структур на ветвях протонем. У немногих родов мхов протонема долгоживущая и выполняет главную фотосинтетическую роль, тогда как листостебельные побеги очень мелкие. Протонемы, характерные для всех мхов, обнаружены также у некоторых печеночников.
У настоящих мхов листостебельный гаметофит обычно прямостоячий, а нс дорсовентрально уплощенный, как у облиственных печеночников, однако нарастает также из апикальной клетки, напоминающей перевернутую трехгранную пирамиду. Хотя первоначально образуются три ряда листьев, последующее скручивание оси приводит к смещению этих рядов и видимости спирального листорасположения. У некоторых родов (например, у водного мха Fontinalis) исходное трехрядное расположение листьев еще заметно и у зрелого гаметофита.
Гаметофиты мхов имеют сложное строение и длину от 0,5 мм до 50 см и более. У всех них — многоклеточные ризоиды, а листья обычно толщиной всего в один слой клеток, кроме средней жилки (отсутствующей у некоторых родов). Как упоминалось выше, в стебле многих мхов присутствует центральный тяж водопроводящих гидроидов, а у некоторых и проводящие органические вещества лептоиды.
Для гаметофитов мхов характерны два способа роста. У одних (часто образующих густые подушки) мхов гаметофиты прямостоячие и маловетвистые, обычно с верхушечными спорофитами. У других — гаметофиты сильно ветвящиеся («перистые»), растения ползучие, спорофиты боковые (рис. 16-19). Второй тип роста встречается у многих мхов, массами свисающих с ветвей деревьев во влажных областях.
Созревая, большинство листостебельных гаметофитов образует гаметангии либо на верхушке главной оси, либо на боковой ветви. У некоторых родов гаметофиты однополые (рис. 16-20), тогда как у других архегонии и антеридии возникают на одной и той же особи.
Рис. 16-13. Жизненный цикл маршанции, широко распространенного слоевищного печеночника. Для маршанции, как и для других печеночников, характерно чередование поколений с доминированием гаметофита (n) над спорофитом (2n). Два важнейших события в жизненном цикле — мейоз и сингамия. Гаметофитное поколение начинается с мейоза, дающего гаплоидные споры. Одни из них прорастают в мужские гаметофиты, другие — в женские. Спорофитное поколение начинается с сингамии, дающей диплоидную зиготу. Подвижным сперминм для достижения яйцеклетки требуется водная среда. Внутри калиптры, или брюшка, архегония зигота развивается в зародыш, или молодой спорофит. По мере роста зародыша калиптра тоже увеличивается в размере. В конечном итоге созревающий спорофит разрывает калиптру, вынося спорангий во внешнюю среду. Спорофит в течение всей жизни прикреплен к гаметофиту расширенной стопой. Подробности строения этого печеночника показаны на рис. 16-6, 16-8 — 16-12

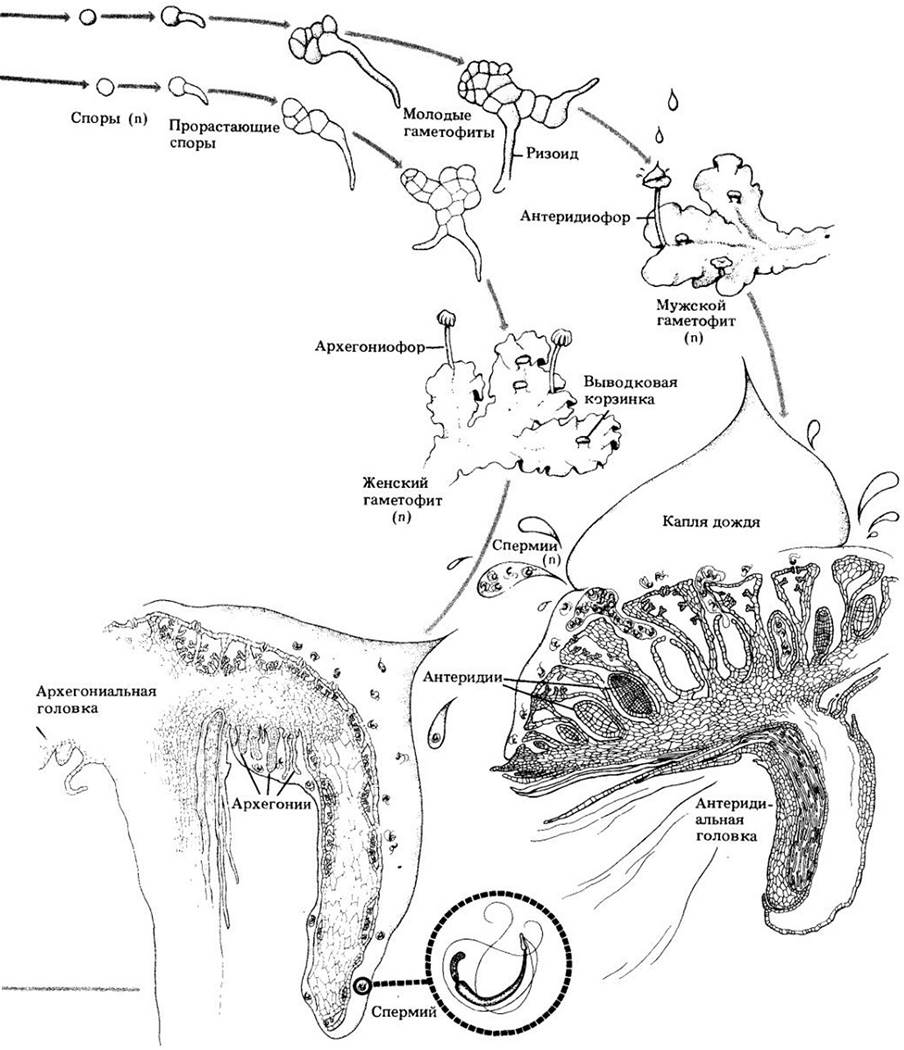
Рис. 16-14. Облиственные печеночники. А. Clasmatocolea puccionana. Видно характерное расположение листьев. Б. Кончик веточки Сlasmatocolea humilis. Видны коробочка и длинная ножка спорофита. В. Часть ветви Frullania с характерным листорасположением
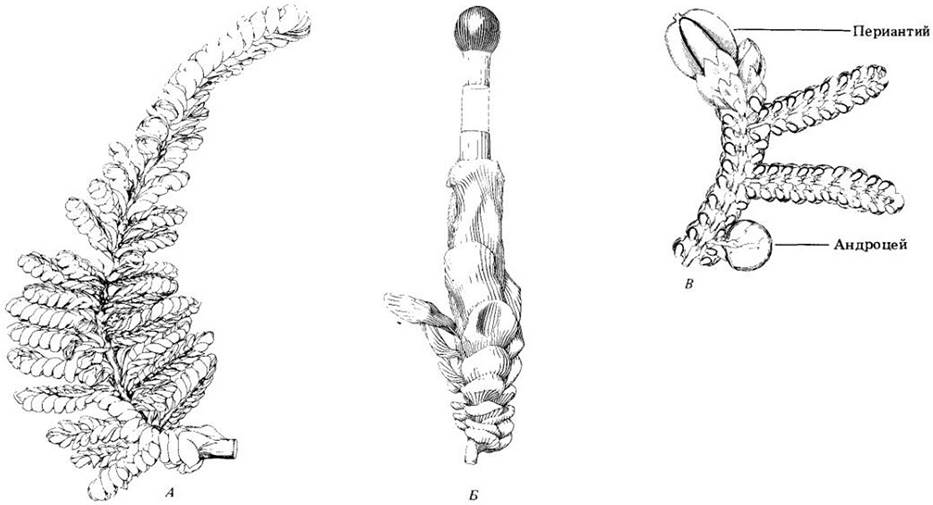
Спорофитам у видов умеренных областей обычно требуется от 6 до 18 месяцев для достижения зрелости; они возникают на гаметофитах (см. рис. 16-3). Коробочки поднимаются на ножке, которая иногда достигает 15 — 20 см длины, однако у некоторых видов совсем отсутствует. Короткая стопа в ее основании погружена в ткани гаметофита. Ножка обычно вытягивается уже на ранней стадии развития спорофита, который в это время способен к фотосинтезу, поэтому, как правило, меньше зависит от гаметофита в отношении питания, чем у печеночников (у них он до созревания обычно остается окружен тканями гаметофита). Устьица, отсутствующие на спорофитах печеночников, в норме присутствуют здесь у мхов. Кроме того, спорофиты последних сложнее организованы внутренне. Ножки большинства из них имеют центральный тяж гидроидов и для многих характерны и проводящие органические вещества лептоиды (см. рис. 16-5).
Рис. 16-15. Облиственный печеночник, растущий на листе вечнозеленого дерева в дождевом лесу бассейна Амазонки близ Манауса (Бразилия)

Рис. 16-16. Антоцерос. А. Гаметофит с прикрепленными к нему спорофитами. Б. Зрелый спорангий расщепляется, высвобождая споры. В. Устьице. На зеленых фотосинтезирующих спорофитах антоцеротовых они встречаются в большом количестве. Г. Развивающиеся споры. Д. Зрелые споры

Рис. 16-17. Антоцерос. А. Поперечный срез спорофита, показывающий его стопу, погруженную в ткань гаметофита. Б. Продольный срез спорангия, показывающий тетрады спор с элатеровидными образованиями среди них. Колонка в центре нижней части спорангия состоит из ткани, возможно, функционирующей как проводящая

Рис. 16-18. Протонема мха с почковидной структурой. Протонема — первая стадия гаметофитного поколения мхов и некоторых печеночников. Она часто напоминает нитчатые зеленые водоросли

Рис. 16-19. Две основные формы роста гаметофитов у мхов. А. «Подушковидный» (на примере Роlytrichum juniperinum); гаметофиты прямостоячие и маловетвистые. Над гаметофитами возвышаются спорофиты — коробочки на длинных тонких ножках. Б. «Перистый», со стелющимися ковром гаметофитами (на примере Thuidium delicatulum)

Рис. 16-20. Гаметангии мха Mnium. А. Продольный срез архегониальной головки; видны окрашенные в лиловый цвет архегонии, окруженные стерильными образованиями, — парафизами. Б. Продольный срез антеридиальной головки; видны антеридии, окруженные парафизами
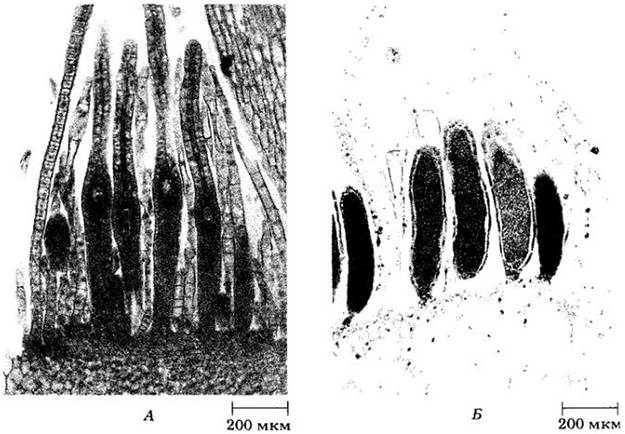
Спорофит мха, созревая, постепенно теряет способность к фотосинтезу и становится сначала желтым, затем оранжевым и наконец бурым. В конце концов крышечка (оперкулум) коробочки опадает, открывая кольцо зубцов — перистом, окружающий отверстие (рис. 16-21). Зубцы перистома образуются путем расщепления клеточного слоя вблизи верхушки коробочки по наименее прочным местам. Этот аппарат, регулирующий рассеивание спор, характерен для подкласса Вryidae и отсутствует в двух других подклассах. Каждая коробочка выбрасывает до 50 млн. гаплоидных спор, каждая из которых способна дать начало новому гаметофиту. Типичный жизненный цикл бриевых мхов приведен на рис. 16-22.
Рис. 16-21. Зубцы перистома у настоящих мхов. А. У Brachythecium перистом состоит из двух колец зубцов и открывается, высвобождая споры, в ответ на изменение влажности воздуха. Во влажных условиях зубцы обоих колец сцеплены между собой, а когда коробочка высыхает, наружные отгибаются и споры могут рассеиваться ветром. Б. Микрофотография (сканирующий электронный микроскоп) зубцов перистома рода Orthotrichum в сухую погоду; видны загнутые внутрь зубцы внутреннего круга и отогнутые наружные
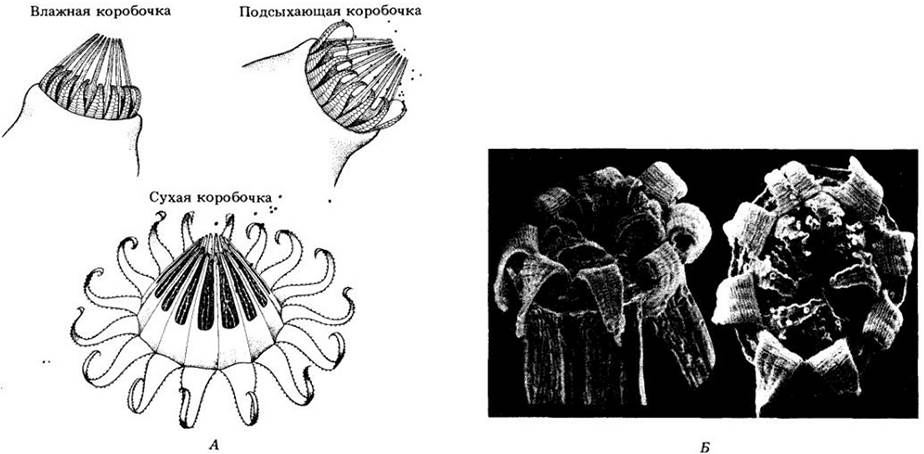
Рис. 16-22. В жизненном цикле бриевого мха споры высвобождаются из коробочки, открывающейся в результате сбрасывания крышечки (оперкулума).
Гаплоидная спора прорастает в разветвленную нитчатую протонему, из которой развивается листостебельный гаметофит. Спермии, выходящие из зрелого антеридия, могут оказаться вблизи архегония; в этом случае они проникают, привлеченные определенными веществами, в канал шейки. Внутри архегония один из спермиев сливается с яйцеклеткой, давая зиготу, которая делится митотически, формируя спорофит. Одновременно брюшко архегония разделяется, образуя калиптру. Спорофит состоит из коробочки, обычно быстро выносящейся вверх на ножке (также его часть), и стопы, посредством которой он получает питательные вещества от гаметофита. Внутри коробочки происходит мейоз с образованием гаплоидных спор. На рисунке представлен жизненный цикл одного из видов рода Роlytrichum.
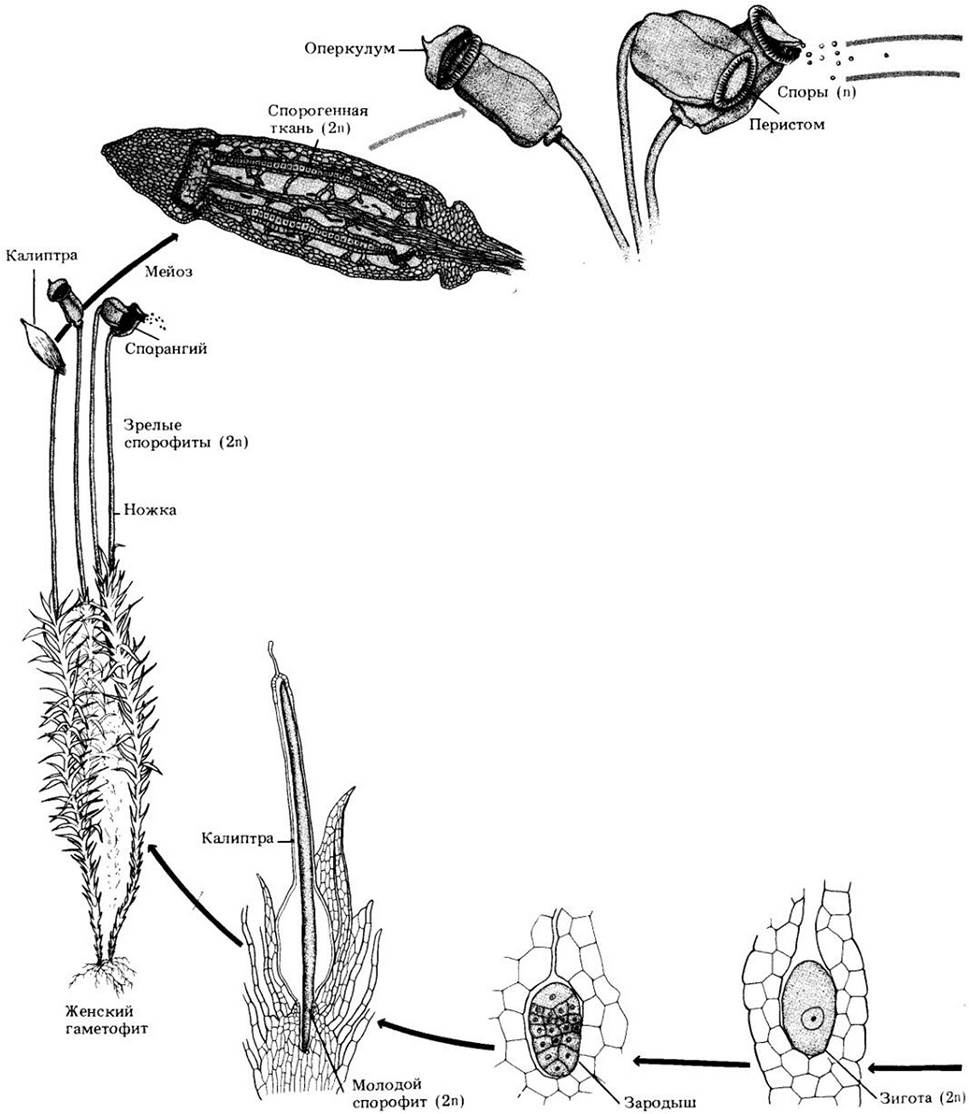
Бесполое размножение этих мхов обычно происходит путем фрагментации. В сущности, любая часть гаметофита, включая стерильные части половых органов, способна к регенерации, а многие виды образуют выводковые почки, которые могут дать начало новым гаметофитам.
Сфагновые мхи
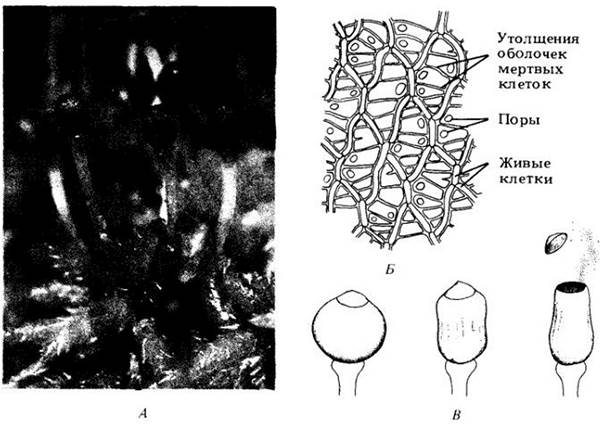
Примерно 350 видов мхов, относящихся к единственному роду сфагнум (Sphagnum), составляют четко очерченную группу, очень давно отделившуюся от основной линии эволюции этого отдела (рис. 16-23). Стебли листостебельного гаметофита у них несут мутовки ветвей, часто по пять в узле, более тесно скученные у верхушки растения, так что возникает своего рода головка. Гаметофиты образуют крупные светло-зеленые или иногда красноватые куртинки на болотистой почве. Они возникают из выводковых почек по краю пластинчатой, а не нитчатой протонемы. Листья сфагнума без средней жилки; у взрослых растений нет ризоидов. На типичных для них болотистых местах эти растения почти всегда имеют хороший тургор, поэтому прямостоячие; обычно они тесно сближены друг с другом. Листья этого рода состоят из широких мертвых клеток, окруженных узкими зелеными или изредка красными живыми клетками (рис. 16-23, Б). Первые имеют поры и утолщения стенок; они легко наполняются водой, так что водоудерживающая способность сфагновых мхов примерно в 20 раз превышает их сухую массу (для сравнения: хлопковая вата может поглотить воды только в 4 — 6 раз больше собственной сухой массы).
Рис. 16-23. Торфяной мох Sphagnum. А. Гаметофит с многочисленными прикрепленными к нему спорофитами. Некоторые из коробочек (например, две слева) уже рассеяли споры. Б. Структура листа. Крупные мертвые клетки окружены более мелкими живыми клетками с хлоропластами. В. Вскрывание коробочки. По мере высыхания она заполняется воздухом. Устьица, через которые входит воздух, закрываются при высыхании. По мере дальнейшего сжимания коробочки возрастающее давление с силой скидывает крышечку и выбрасывает в воздух облачко спор
Благодаря своим превосходным абсорбирующим свойствам эти мхи использовались в Европе с 1880-х гг. как перевязочный материал для ран и нарывов, однако со времени первой мировой войны были почти полностью вытеснены в этом смысле хлопком, вероятно, из-за более опрятного вида изделий из него. Садоводы смешивают торфяной мох с почвой для увеличения ее водоудерживающей способности и кислотности.
Спорофиты сфагнума (рис. 16-23, А) также очень своеобразны: от красных до черновато-бурых по цвету коробочки почти сферические и приподнимаются на ложноножке (псевдоподии), являющейся частью гаметофита и достигающей 3 мм длины. Ножка самого спорофита очень короткая. Рассеивание спор происходит очень эффективным способом (рис. 16-23, В). На верхушке коробочки дисковидный оперкулум отделен от остальной ес части круговой бороздкой. Когда коробочка созревает, ее внутренние ткани сморщиваются и воздух втягивается внутрь, возможно через незакрывающиеся устьица. Когда стенка высыхает, он остается внутри, как в ловушке. Дальнейшее сжатие созревающей коробочки вызывает увеличение внутреннего давления, и оперкулум в конце концов отскакивает со слышимым треском. Это происходит в теплую солнечную погоду. Выходящий наружу газ выносит из коробочки облачко спор. Наиболее характерные признаки подкласса Sphagnidae — отсутствие перистома и своеобразная морфология гаметофита.
Экология сфагнума
Представители этого рода образуют обширные торфяные болота, являющиеся отличительной чертой холодных и умеренных областей земного шара. Мхи способствуют увеличению кислотности своего местообитания, высвобождая ионы водорода, поэтому pH в центре болота часто ниже 4, что очень необычно для естественной среды. Торф образуется в результате накопления и уплотнения самих сфагновых мхов, а также осок, злаков и других растений, которые встречаются вместе с ними. В Ирландии и некоторых других северных областях сухой торф широко используется как топливо. По самым скромным оценкам торфяники покрывают нс менее 1 % земной суши, что эквивалентно половине площади США. Поскольку мировые запасы торфа так велики, с каждым годом растет интерес к нему как возможному важному источнику промышленной энергии. Однако при этом необходимо тщательно изучить потенциальное влияние добычи торфа на те области, где она ведется, а также последствия выделения в атмосферу большого количества двуокиси углерода при сжигании такого горючего.
Андреевые мхи
Род андрея (Andreae) состоит примерно из 100 видов мелких черновато-зеленых или темно-красновато-бурых, образующих дерновинки наскальных мхов (рис. 16-24), которые по- своему не менее своеобразны, чем сфагнум. Представители этой группы встречаются в горных или арктических областях, часто на гранитных скалах. Хотя их гаметофит очень похож на гаметофит настоящих мхов, он возникает из толстой многолопастной протонемы, а нс из нитчатой структуры. У спорофита нет настоящей ножки, и он возвышается над листьями на стебельке из ткани гаметофита, т. с. на ложноножке, как у сфагновых мхов. По мелкой коробочке андреи проходят четыре вертикальных ряда менее прочных клеток, вдоль которых она вскрывается, но остается целой выше и ниже линий растрескивания. Образующиеся четыре створки очень чувствительны к влажности окружающего воздуха, широко раздвигаясь, когда он сухой (споры при этом могут далеко разноситься ветром), и смыкаясь, если он влажный. Этот механизм рассеивания спор за счет продольного расщепления коробочки отличается от наблюдаемого у всех других мхов (рис. 16-24).
Рис. 16-24. А — Andreae rothii на гранитных скалах в Девоне, Англия. Б — спорангий рода Andreaea сжимается при высыхании, расщепляясь на четыре створки, что позволяет спорам рассеиваться в сухую погоду

Andreaeobryum — второй род андреевых мхов, открытый на Аляске в 1976 г. — отличается от Andreaea присутствием настоящей ножки у спорофита и расщеплением коробочки до верхушки.
А — Splachnum luteum. Растения этого вида с кремовыми апофизами напоминают мелкие цветы. Б — окраска растений и выделяемые ими химические вещества привлекают насекомых. В — Splachnum rubrum, сфотографированный на лосином помете в провинции Альберта (Канада)