СОВРЕМЕННАЯ БОТАНИКА - П. РЕЙВН - 1990
РАЗДЕЛ I. РАСТИТЕЛЬНАЯ КЛЕТКА
ГЛАВА 2. ЭУКАРИОТИЧЕСКАЯ КЛЕТКА
Растительная клетка
Растительная клетка состоит, как правило, из более или менее жесткой клеточной оболочки1 и протопласта. Термин протопласт происходит от слова протоплазма, которое долго применялось для обозначения живого содержимого клетки. Протопласт — это протоплазма индивидуальной клетки; в растительной клетке — протоплазма, ограниченная клеточной оболочкой.
1Наряду с термином «клеточная оболочка» встречается и равнозначный — «клеточная стенка». Однако в русской литературе по цитологии и анатомии растений термин «клеточная оболочка» принят как приоритетный. — Прим. перев.
Протопласт состоит из цитоплазмы и ядра (см. табл. 2-1). В цитоплазме находятся органеллы (такие, как рибосомы, микротрубочки, пластиды и митохондрии) и мембранные системы (эндоплазматический ретикулум и диктиосомы). Их можно подробно рассмотреть только в электронный микроскоп. Кроме того, цитоплазма включает в себя цитоплазматический матрикс, или основное вещество, в которое погружены органеллы и мембранные системы. Цитоплазма отделена от клеточной оболочки плазматической мембраной, которая представляет собой элементарную мембрану. В отличие от большинства животных клеток в цитоплазме растительных клеток содержатся одна или несколько вакуолей. Вакуоли — это пузырьки, заполненные жидкостью и окруженные элементарной мембраной, называемой тонопластом.
В живой растительной клетке основное вещество находится в постоянном (рис. 2-4) движении. Можно видеть, как органеллы и другие включения вовлекаются в это движение, называемое током цитоплазмы, или циклозом. Циклоз прекращается в мертвых клетках. Он, несомненно, облегчает передвижение веществ в клетке и обмен ими между клеткой и окружающей средой. Однако основная функция циклоза неизвестна.
Таблица 2-1. Компоненты растительной клетки

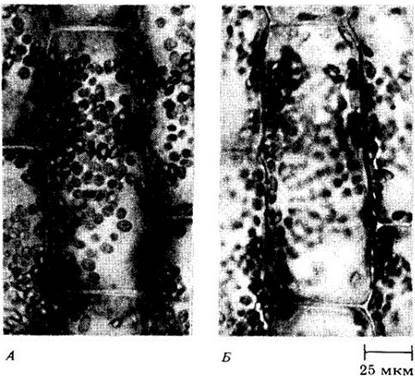
Рис. 2-4. Клетка элодеи. А. Поверхность клетки. Б. Середина клетки. Многочисленные дископодобные структуры — это хлоропласты, расположенные вдоль клеточной оболочки. При взгляде сверху (А) хлоропласты имеют округлые очертания. Б. Хлоропласты обращены своими широкими плоскостями к стенке и кажутся удлиненными. Отметим отсутствие хлоропластов в центре клетки (Б), т. е. в вакуоли
Плазматическая мембрана
Среди многочисленных мембран клетки в плазматической мембране наиболее ясно выражена трехслойность (темный — светлый — темный слои). Для растительных клеток обычны инвагинации (впячивания) плазматической мембраны.
Плазматическая мембрана выполняет следующие функции: (7) участвует в обмене веществ между клеткой и окружающей средой (см. гл. 4); (2) координирует синтез и сборку 'целлюлозных микрофибрилл клеточной оболочки; (3) передает гормональные и внешние сигналы, контролирующие рост и дифференцировку клеток.
Ядро
Ядро — часто наиболее заметная структура в цитоплазме эукариотической клетки. Ядро выполняет две важные функции: (7) контролирует жизнедеятельность клетки, определяя, какие белки и в какое время должны синтезироваться (см. гл. 8); (2) хранит генетическую информацию и передает ее дочерним клеткам в процессе клеточного деления.
Ядро эукариотической клетки окружено двумя элементарными мембранами, образующими ядерную оболочку. Она пронизана многочисленными порами диаметром от 30 до 100 нм, видимыми в электронный микроскоп (см. рис. 2-5). Поры — это не просто отверстия в оболочке: они имеют сложную структуру. Наружная мембрана ядерной оболочки в некоторых местах объединяется с эндоплазматическим ретикулумом. Ядерную оболочку можно рассматривать как специализированную, локально дифференцированную часть эндоплазматического ретикулума.
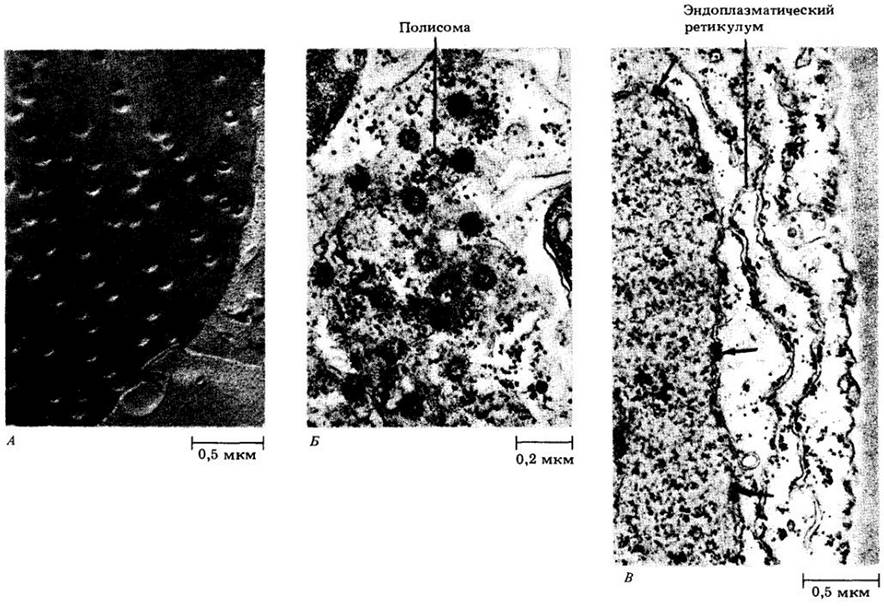
Рис. 2-5. Ядерные поры (А), видимые на поверхности ядерной оболочки в клетке кончика корня лука Аllium сера. Препарат получен техникой замораживания- травления. Б, В. Электронные микрофотографии ядер не имеющего семян сосудистого растения Selaginella kraus-siana. Б. Поры видны с поверхности: В — на срезе (отмечены стрелками). Обратите внимание на полисомы (спирали из гранул) на поверхности ядерной оболочки (Б) и шероховатый эндоплазматический ретикулум, расположенный параллельно ядерной оболочке (В)
В ядре, окрашенном специальными красителями, можно различить тонкие нити и глыбки хроматина и нуклеоплазму, или основное вещество ядра. Хроматин состоит из ДНК, связанной с большим количеством специальных белков — гистонов. В процессе клеточного деления хроматин все более уплотняется и в конце концов собирается в хромосомы. Недавно было высказано предположение, что в неделящихся (интерфазных) ядрах хромосомы (хроматин) прикреплены к ядерной оболочке в одном или нескольких местах. Генетическая информация у прокариот и эукариот закодирована в молекулах ДНК. Содержание ДНК в одной клетке эукариот значительно выше, чем в бактериальной клетке. У бактерий молекулы ДНК свободно располагаются в цитоплазме, а у эукариот они организованы в хромосомы, находящиеся в ядре.
Организмы различаются по числу хромосом в соматических (неполовых) клетках. Пустынный однолетник Haplopappus gracilis имеет 4 хромосомы; капуста — 20; подсолнечник — 34; пшеница — 42; человек — 46; а один из видов папоротника Ophioglossum — около 1250. Половые клетки, или гаметы, имеют только половину количества хромосом, характерного для соматических клеток организма. Число хромосом в гаметах называют гаплоидным (одинарным), в соматических клетках — диплоидным (двойным). Клетки, имеющие более двух наборов хромосом, называют полиплоидными.
Под световым микроскопом в ядре можно различить сферические структуры — ядрышки. В каждом ядре имеется одно или несколько ядрышек, которые заметны в неделящихся ядрах (рис. 2-1). В ядрышках синтезируются рибосомные РНК. Обычно в ядрах диплоидных организмов имеются два ядрышка, по одному для каждого гаплоидного набора хромосом. Состоя в основном из белка, ядрышки содержат около 5% РНК.
Пластиды
Наряду с вакуолями и клеточной оболочкой пластиды — характерные компоненты растительных клеток. Каждая пластида окружена собственной оболочкой, состоящей из двух элементарных мембран. Внутри пластиды различают мембранную систему и более или менее гомогенное вещество — строму. Зрелые пластиды обычно классифицируют на основании содержащихся в них пигментов.
Хлоропласты, в которых протекает фотосинтез (см. гл. 7), содержат хлорофиллы и каротиноиды. Хлоропласты растений обычно имеют форму диска диаметром от 4 до 5 мкм. В одной клетке мезофилла («середины листа») может находиться 40 — 50 хлоропластов; в квадратном миллиметре листа — около 500 000. В цитоплазме хлоропласты обычно располагаются параллельно клеточной оболочке, как показано на рис. 2-6.
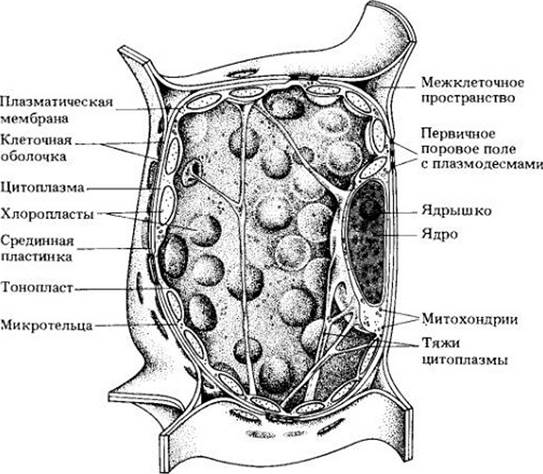
Рис. 2-6. Трехмерная схема растительной клетки, содержащей хлоропласты. Многочисленные хлоропласты располагаются в цитоплазме вдоль клеточной оболочки, к которой они обращены своими широкими поверхностями. Основной объем клетки занят вакуолью, пронизанной тяжами цитоплазмы. Ядро расположено вдоль оболочки, в других случаях оно может быть подвешено на тяжах цитоплазмы в центре вакуоли и окружено тонким слоем цитоплазмы
Внутренняя структура хлоропласта довольно сложна (рис. 2-7). Строма пронизана развитой системой мембран, имеющих форму плоских пузырьков, называемых тилакоидами. Каждый тилакоид, так же, как и оболочка хлоропласта, состоит из двух мембран. Считается, что тилакоиды образуют единую систему. Как правило, тилакоиды собраны в стопки —так называемые граны, напоминающие столбики монет. Тилакоиды отдельных гран связаны друг с другом тилакоидами стромы, или межгранными тилакоидами. Хлорофиллы и каротиноиды встроены в тилакоидные мембраны. Хлоропласты зеленых водорослей и растений часто содержат зерна крахмала и мелкие липидные (жировые) капли. Крахмальные зерна — это временные хранилища продуктов фотосинтеза (рис. 2-1). Они могут исчезнуть из хлоропластов растения, находящегося в темноте всего лишь 24 ч, и появиться вновь уже через 3 — 4 ч после переноса растения на свет.
Рис. 2-7. А. Хлоропласт листа кукурузы (Zea mays). Б. Гркны, состоящие из стопок дископодобных тилакоидов. Тилакоиды гран соединяются посредством других тилакоидов, обычно называемых тилакоидами стромы

Хлоропласты — полуавтономные органеллы и в некотором отношении напоминают бактерии. Например, рибосомы как бактерий, так и хлоропластов на одну треть меньше рибосом эукариот. Синтез белка на рибосомах бактерий и хлоропластов подавляется антибиотиком хлорамфениколом, не оказывающим подобного действия в клетках эукариот. Кроме того, и бактерии, и хлоропласты имеют один или несколько нуклеоидов — светлых, лишенных гран участков, содержащих тяжи ДНК. ДНК пластид и бактерий организована сходным образом, а именно: не окружена мембраной, не связана с гистонами и обычно существует в кольцевой форме.
Генетический код пластидной ДНК в настоящее время изучается в нескольких лабораториях. В изолированных хлоропластах осуществляется синтез РНК, который, как будет показано в гл. 8, обычно контролируется только хромосомной ДНК. Образование хлоропластов и синтез находящихся в них пигментов в значительной степени контролируются хромосомной ДНК, малопонятным образом взаимодействующей с ДНК хлоропластов. Тем не менее в отсутствие собственной ДНК хлоропласты не формируются.
Хлоропласты могут считаться основными клеточными органеллами, поскольку первыми стоят в цепи преобразования солнечной энергии, в результате которого мы получаем пищу и топливо. В хлоропластах протекает не только фотосинтез. Они участвуют и в синтезе аминокислот и жирных кислот, служат хранилищем временных запасов крахмала.
Хромопласты (от греч. chroma — цвет) — пигментированные пластиды (рис. 2-8). Многообразные по форме хромопласты не имеют хлорофилла, но синтезируют и накапливают каротиноиды, которые придают желтую, оранжевую и красную окраску цветкам, старым листьям (см. гл. 7), плодам и корням. Хромопласты могут развиваться из зеленых хлоропластов; последние при этом теряют хлорофилл и внутренние мембранные структуры и накапливают каротиноиды. Это происходит, например, при созревании многих фруктов. Точная функция хромопластов неизвестна, хотя в некоторых случаях они привлекают насекомых и других животных, с которыми вместе эволюционировали (см. гл. 29).
Рис. 2-8. Хромопласты из лепестка ноготков (Tagetes). Каждый хромопласт содержит многочисленные липидные капли, в которых откладываются пигменты, определяющие цвет лепестков
Лейкопласты (рис. 2-9) — непигментированные пластиды. Некоторые синтезируют крахмал (амилопласты; рис. 2- 10 и 2-11), другие, по-видимому, способны к образованию разных веществ, в том числе липидов и белков. На свету лейкопласты могут превращаться в хлоропласты.
Рис. 2-9. Лейкопласты, группирующиеся вокруг ядра в эпидермальных клетках листа Zebrina

Рис. 2-10. На микрофотографии, сделанной с помощью сканирующего электронного микроскопа, видны сферические и яйцевидные крахмальные зерна в клетке картофеля (Solanит tuberosum). Крахмальные зерна образуются в амилопластах, по одному зерну в пластиде

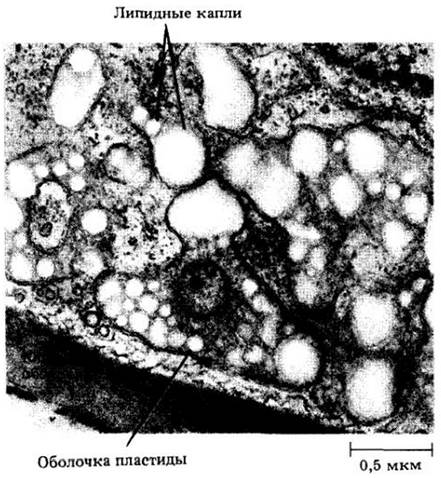
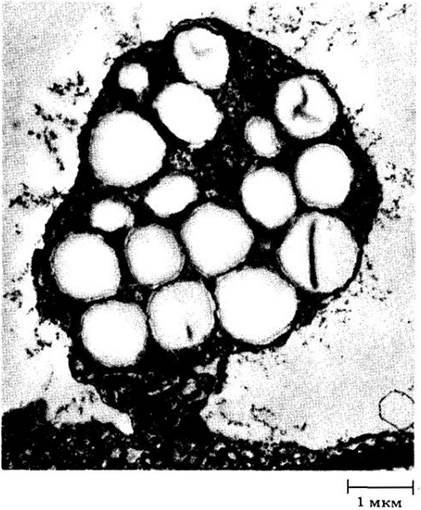
Рис. 2-11. Амилопласт из зародышевого мешка сои (Glycine max). Округлые светлые тельца — крахмальные зерна, более мелкие темные — липидные капли
Пропластиды — мелкие бесцветные или бледно-зеленые недифференцированные пластиды, которые находятся в меристематических (делящихся) клетках корней и побегов. Они являются предшественниками других, более дифференцированных пластид — хлоропластов, хромопластов и амилопластов (рис. 2-12). Если развитие пропластид в более дифференцированные структуры задерживается из-за отсутствия света, в них может появиться одно или несколько проламеллярных телец, представляющих собой полукристаллические скопления трубчатых мембран (рис. 2-13). Пластиды, содержащие проламеллярные тельца, называют этиопластами. Этиопласты превращаются в хлоропласты на свету; при этом мембраны проламеллярных телец формируют тилакоиды. Этиопласты образуются в листьях растений, находящихся в темноте. Пропластиды зародышей семян вначале превращаются в этиопласты, из которых на свету затем развиваются хлоропласты. Для пластид характерны относительно легкие переходы от одного типа к другому.
Рис. 2-12. Цикл развития пластид по J. М. Whatley. Последовательные стадии развития пластид. Стадии 1 — 3: эопласт (ео — «ранний*), амилопласт и амебоидную пластиду можно считать незелеными пропластидными стадиями. Стадия 4: до гранильная пластида может быть зеленой или незеленой. В отсутствие света догранальная стадия может быть представлена этиопластом (стадия 4'). Хромопласты (У) могут образоваться из пластид нескольких типов
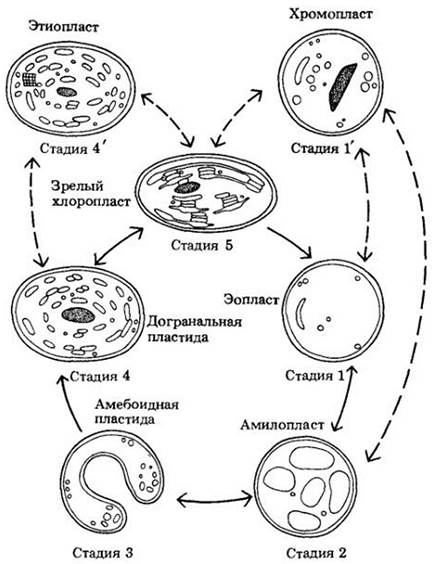
Рис. 2-13. Этиопласт с полукристаллическим проламеллярным тельцем, построенным из тубулярных мембран, в клетке листа выросшего в темноте растения. На свету мембраны проламеллярного тельца превращаются в тилакоиды

Пластиды размножаются делением надвое, и в этом отношении они также напоминают бактерии. В меристематичес- ких клетках 'ремя деления пропластид приблизительно совпадает с временем деления клеток. Однако в зрелых клетках Обльшая часть пластид образуется в результате деления зрелых пластид.