СОВРЕМЕННАЯ БОТАНИКА - П. РЕЙВН - 1990
РАЗДЕЛ I. РАСТИТЕЛЬНАЯ КЛЕТКА
ГЛАВА 2. ЭУКАРИОТИЧЕСКАЯ КЛЕТКА
Митохондрии
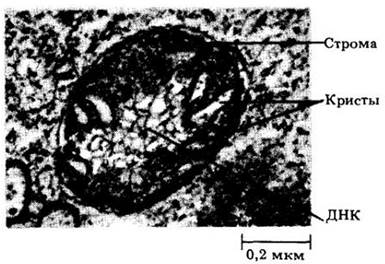
Как и хлоропласты, митохондрии окружены двумя элементарными мембранами (рис. 2-14 и 2-15). Внутренняя мембрана образует множество складок и выступов, называемых кристами, которые значительно увеличивают внутреннюю поверхность митохондрии. Митохондрии обычно мельче, чем пластиды, имеют около половины микрометра в диаметре и очень разнообразны по длине и форме. Как правило, митохондрии плохо видны под световым микроскопом, но их можно хорошо рассмотреть с помощью электронного микроскопа.
Рис. 2-14. Разрез митохондрии из клетки листа шпината (Spinacia oleracea). Видны тяжи ДНК. Оболочка митохондрии состоит из двух отдельных мембран. Внутренняя мембрана образует кристы, погруженные в строму. Мелкие гранулы в строме — рибосомы
В митохондриях осуществляется процесс дыхания, в результате которого органические молекулы расщепляются с высвобождением энергии и передачей ее молекулам АТР (аденозинтрифосфата), основного резерва энергии всех эукариотических клеток. (Процесс дыхания обсуждается ниже, в гл. 6.) Большинство растительных клеток содержит сотни и тысячи митохондрий, их число в одной клетке определяется потребностью клетки в АТР.
С помощью прерывистой съемки можно увидеть, что митохондрии находятся в постоянном движении. Они поворачиваются, изгибаются, перемещаются из одной части клетки в другую, а кроме того, сливаются друг с другом и делятся. Митохондрии обычно собираются там, где нужна энергия.
Если плазматическая мембрана активно переносит вещества из клетки или в клетку, то митохондрии располагаются вдоль поверхности мембраны. У подвижных одноклеточных водорослей митохондрии, как правило, скапливаются у оснований жгутиков и, по-видимому, поставляют энергию, необходимую для их движения.
Митохондрии, подобно пластидам, являются полуавтономными органеллами, содержащими компоненты, необходимые для синтеза собственных белков. Внутренняя мембрана окружает жидкий матрикс, в котором находятся белки, РНК, тяжи ДНК, рибосомы, сходные с бактериальными, и различные растворенные вещества. ДНК существует в виде кольцевых молекул, располагающихся в одном или нескольких нуклеоидах (рис. 2-14).
Происхождение митохондрий и хлоропластов
На основании сходства бактерий с митохондриями и хлоропластами эукариотических клеток можно предположить, что митохондрии и хлоропласты произошли от бактерий, которые нашли «убежище» в более крупных гетеротрофных клетках — предшественниках эукариот. Бактерии, имеющие возможность использовать молекулярный кислород для окисления питательных веществ и использовать энергию света, придавали эти полезные свойства более крупным клеткам, которые с такими «помощниками» имели явное преимущество перед своими современниками и, несомненно, вскоре размножились за их счет. Все ныне живущие эукариоты, за очень малым исключением, содержат митохондрии, а все автотрофные эукариоты содержат также хлоропласты. По- видимому, они были приобретены в результате независимых случаев симбиоза (симбиозом называется тесное объединение двух или более организмов, которое может быть, но не обязательно полезно для каждого). Более крупные и более сложные клетки эукариот, по-видимому, защищали свои симбиотические органеллы от неблагоприятных внешних воздействий. В результате эукариоты смогли захватить сушу и кислые воды, в которых отсутствуют прокариотические цианобактерии (или «сине-зеленые водоросли»), но в изобилии живут эукариотические зеленые водоросли.
Микротельца
В отличие от пластид и митохондрий, которые ограничены двумя мембранами, микротельца представляют собой сферические органеллы, окруженные одной мембраной. Их диаметр колеблется от 0,5 до 1,5 мкм. Микротельца имеют гранулярное содержимое, иногда в них встречаются и кристаллические белковые включения (рис. 2-15). Микротельца обычно связаны с одним или двумя участками эндоплазматического ретикулума.
Рис. 2-15. Органеллы в клетке листа табака (Nicotiana tabacum). Микротельце с крупным кристаллическим включением окружено одной мембраной и тем самым отличается от двух митохондрий и пластиды, окруженных двумя мембранами. Двойную мембрану пластиды видно только в нижней части микрофотографии. Часть вакуоли (светлый участок справа вверху) отделена от остальной цитоплазмы одиночной мембраной, тонопластом
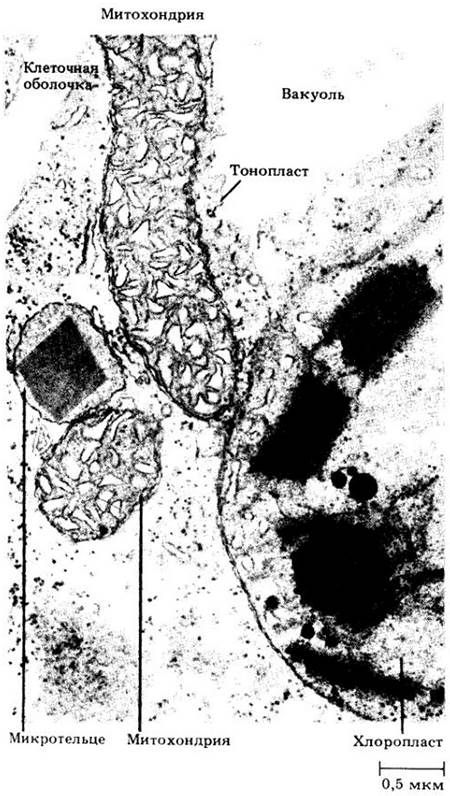
Некоторые микротельца, называемые пероксисомами, играют важную роль в метаболизме гликолевой кислоты, имеющем непосредственное отношение к фотодыханию (см. с. 105). В зеленых листьях они обычно связаны с митохондриями и хлоропластами (рис. 2-15). Другие микротельца, называемые глиоксисомами, содержат ферменты, необходимые для превращения жиров в углеводы, что происходит во многих семенах во время прорастания (см. с.92).
Вакуоли
Вакуоли — это ограниченные мембраной участки клетки, заполненные жидкостью, которая называется клеточным соком. Они окружены тонопластом, или вакуолярной мембраной (рис. 2-6 и 2-15).
Молодая растительная клетка обычно содержит многочисленные мелкие вакуоли, которые увеличиваются в размерах и сливаются в одну вакуоль по мере старения клетки. В зрелой клетке до 90% ее объема может быть занято вакуолью, тогда как цитоплазма в виде тонкого периферического слоя прижата к клеточной оболочке (рис. 2-6). Заполняя большую часть клетки «дешевым» вакуолярным содержимым, растения экономят потребляющую азот «дорогую» цитоплазму, в то время как ее поверхность остается обширной. В основном увеличение размера клетки происходит за счет роста вакуоли. В результате этого возникает тургорное давление и поддерживается упругость ткани, в чем и заключается одна из главных функций вакуоли и тонопласта (см. гл. 4).
Основной компонент клеточного сока — вода, остальные варьируют в зависимости от типа растения и его физиологического состояния. Обычно вакуоли содержат соли и сахара, иногда — растворимые белки. Тонопласт играет важную роль в активном транспорте и накоплении в вакуоли некоторых ионов. Концентрация ионов в клеточном соке, таким образом, может быть выше, чем в окружающей цитоплазме. При высоком содержании некоторых веществ в вакуолях могут образоваться кристаллы. Особенно часто встречаются кристаллы оксалата кальция, имеющие различную форму (рис. 2-16 и 2-17). Обычно содержимое вакуолей слабокислое, иногда — очень кислое, как в плодах лимона.
Рис. 2-16. Друзы кристаллов оксалата кальция в эпидермальных клетках семядоли церциса канадского (Cercis canadensis). Микрофотография сделана с помощью сканирующего электронного микроскопа

Рис. 2-17. Рафиды — игольчатые кристаллы оксалата кальция — в вакуоли клетки листа Sansevieria. Тонопласт на этой микрофотографии не виден. Гранулярная субстанция, окружающая кристаллы, — цитоплазма
Вакуоли — места накопления различных метаболитов (продуктов метаболизма), таких, как запасные белки в семенах и яблочная кислота у САМ-растений (см. с. 106). Они удаляют из цитоплазмы и ядовитые вторичные продукты метаболизма, например, алкалоид никотин (см. гл. 29).
В вакуолях часто откладываются пигменты. Голубой, фиолетовый, пурпурный, темно-красный и пунцовый цвета придают растительным клеткам пигменты из группы антоцианов (см. гл. 29). В отличие от большинства других растительных пигментов антоцианы легко растворяются в воде и содержатся в клеточном соке. Они определяют красную и голубую окраску многих овощей (редис, турнепс, капуста), фруктов (виноград, сливы, вишни) и множества цветов (васильки, герани, дельфиниумы, розы и пионы). Иногда эти пигменты маскируют в листьях хлорофилл, как, например, у декоративного красного клена. Антоцианы окрашивают осенние листья в ярко-красный цвет. Они образуются в холодную солнечную погоду, когда в листьях прекращается синтез хлорофилла. По мере разрушения имеющегося хлорофилла проявляются антоцианы. В листьях, где эти пигменты не образуются, после разрушения хлорофилла становятся заметными желто-оранжевые каротиноиды хлоропластов. Наиболее ярко окрашены листья холодной ясной осенью.
Вакуоли участвуют и в разрушении макромолекул, и в круговороте их компонентов в клетке. Отдельные органеллы, такие, как рибосомы, митохондрии и пластиды, могут попадать в вакуоли и там разрушаться. По этой переваривающей активности вакуоли сравнимы с лизосомами — органеллами, которые встречаются в клетках животных.
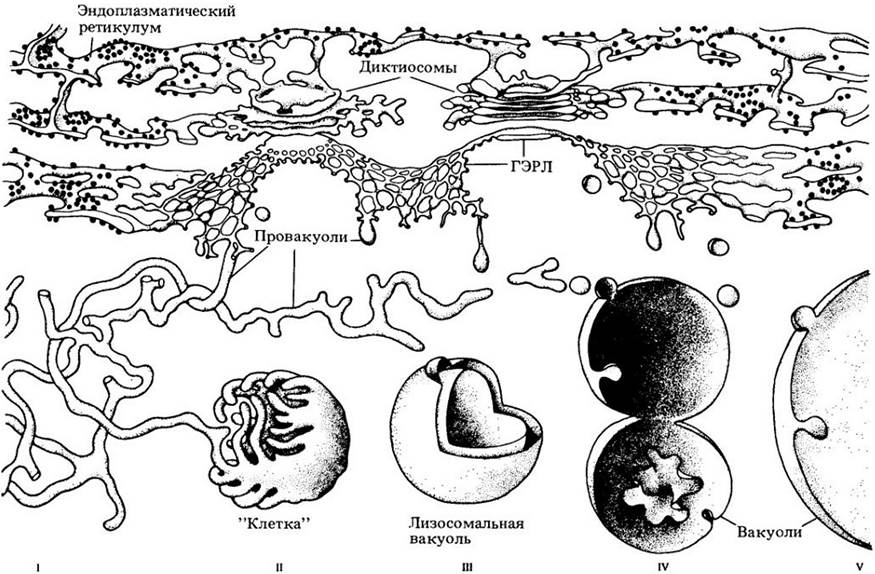
Давно высказано предположение, что вакуоли образуются из эндоплазматического ретикулума. Когда стало ясно, что вакуоли аналогичны лизосомам, были предприняты попытки определить, не образуются ли вакуоли, по крайней мере у некоторых растений, так же, как лизосомы. В клетках животных формирование лизосом связано со специализированным участком цитоплазмы, который был назван ГЭРЛ, т. е. связанный с аппаратом Гольджи и эндоплазматическим ретикулумом лизосомальный комплекс. ГЭРЛы были найдены в клетках кончика корня редиса и молочая и в семядолях зародыша маша (Phaseolus aureus). На рис. 2-18 показаны стадии образования вакуолей, связанные с ГЭРЛ. Аппарат Гольджи рассматривается ниже.
Рис. 2-18. Стадии образования вакуолей по F. Marty. Трубчатые провакуоли, которые возникают из комплекса ГЭРЛ, вначале выглядят как грушевидные пузырьки, затем разрастаются и разветвляются по всей клетке (I). Провакуоли отгораживают участки цитоплазмы, как прутья клетку (II). Трубчатые «прутья» каждой «клетки»
сливаются и полностью изолируют захваченную цитоплазму (III). Внутренняя мембрана «клетки» и захваченная цитоплазма перевариваются лизосомальными ферментами (IV). До этого момента пищеварительные ферменты содержались в сливающихся между собой трубочках. Наружная мембрана «клетки» остается интактной, и пищеварительная активность сохраняется внутри развивающейся вакуоли. Образовавшиеся многочисленные мелкие вакуоли сливаются (IV), и в конце концов возникает одна или несколько более крупных вакуолей, характерных для зрелой растительной клетки (V)
Рибосомы
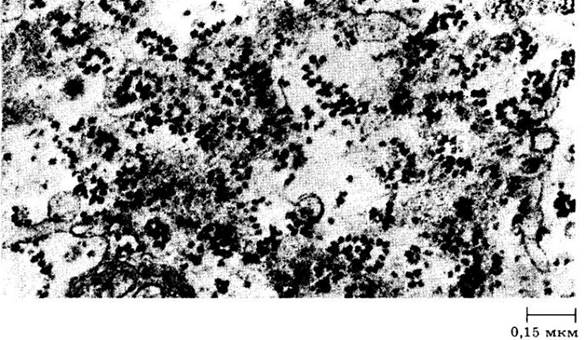
Рибосомы — маленькие частицы всего около 17 — 23 нм в диаметре, состоящие примерно из равных количеств белка и РНК. В рибосомах аминокислоты соединяются с образованием белков; они наиболее многочисленны в цитоплазме клеток с активным метаболизмом. Рибосомы располагаются в цитоплазме клетки свободно или же прикрепляются к эндоплазматическому ретикулуму. Обнаруживают их и в ядре. Как, указывалось выше, пластиды и митохондрии содержат рибосомы, подобные таковым у прокариот.
Рибосомы, активно участвующие в синтезе белка, образуют комплексы, называемые полирибосомами, или полисомами (рис. 2-19). Клетки, синтезирующие белки в больших количествах, имеют обширную систему полисом. Полисомы часто прикреплены к наружной поверхности оболочки ядра (рис. 2-5, Б).
Рис. 2-19. Группы рибосом (полисомы) на поверхности эндоплазматического ретикулума. Эндоплазматический ретикулум — это сеть мембран, пронизывающих цитоплазму эукариотической клетки и делящих ее на отдельные компартменты. На поверхности эндоплазматического ретикулума могут протекать различные химические реакции. Рибосомы — структуры, в которых аминокислоты объединяются в белки. На данной электронной микрофотографии показан участок паренхимной клетки листа папоротника Regnellidium diphyllum