СОВРЕМЕННАЯ БОТАНИКА - П. РЕЙВН - 1990
РАЗДЕЛ VIII. ЭВОЛЮЦИЯ
ГЛАВА 28. ПРОЦЕСС ЭВОЛЮЦИИ
В 1831 г. двадцатидвухлетним молодым человеком Чарльз Дарвин (рис. 28-1) отправился в пятилетнее путешествие в качестве натуралиста на британском военном корабле «Бигль». Об этом путешествии он написал книгу «Путешествие на «Бигле», которая представляет собой не только классическую работу по естественной истории, но и позволяет понять, каким образом Дарвин подошел к созданию теории эволюции, в основе которой лежит понятие о естественном отборе.
В те времена, когда Дарвин совершал свое историческое путешествие, большинство ученых, подобно людям, далеким от науки, все еще признавало теорию «отдельных актов творения». Согласно этой теории, каждая из множества форм различных живых организмов была создана или, иначе говоря, начала свое существование в ее современном виде. Некоторые ученые, например, Жан Батист дс Ламарк (1744 — 1829), высказывали сомнения относительно теории отдельных актов творения, но их доказательства были не настолько убедительны, чтобы поколебать веру большинства людей. Теорию отдельных актов творения нельзя считать собственно научной гипотезой, но тем не менее она прочно утвердилась во всей западной культуре.
Рис. 28-1. «Впоследствии, когда мы с Фицроем (капитаном «Бигля») стали близкими друзьями, я узнал, что мне чуть было не отказали в участии в путешествии из-за формы моего носа! Он... был убежден в том. что мог судить о характере человека по чертам его лица; он сомневался в том, чтобы кто-нибудь с таким носом, как мой, мог обладать достаточной энергией и решительностью для подобного путешествия. Я полагаю, что впоследствии он был рад, что мой нос ввел его в заблуждение» (Из книги Дарвина «Путешествие на «Бигле»)

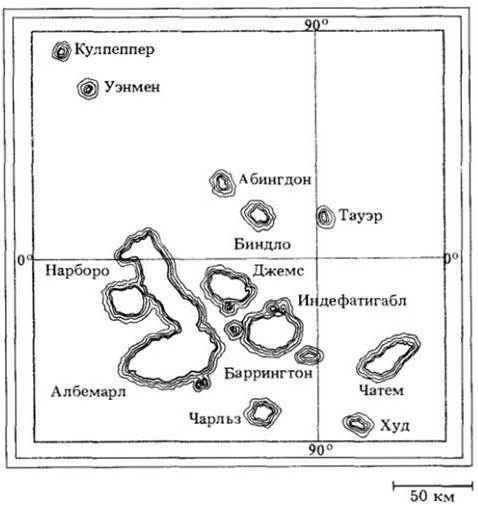
Теория Дарвина смогла произвести решительный переворот в научном мышлении уже только потому, что множество представленных ею доказательств было в высшей степени убедительно и не оставляло места для сколько-нибудь серьезной, научно обоснованной критики. Особенно важную роль в формировании идей Дарвина сыграли впечатления, полученные им на островах Галапагос — архипелаге, расположенном в экваториальных водах на расстоянии около 950 км от западного берега Южной Америки (рис. 28-2). На островах он пробыл пять недель и сделал два особо важных наблюдения. Во- первых, он заметил, что растения и животные этих островов хотя чем-то и отличались, но в целом были похожи на тех, которые обитали на соседнем южноамериканском материке. Если каждая форма растения или животного создавалась отдельно, как принято считать, и оставалась неизменной, то почему бы растениям и животным островов Галапагос не быть похожими на африканских, например, а не на южноамериканских? Или почему здесь нет совершенно уникальных организмов, не похожих ни на какие другие, живущие где-либо на земле? Во-вторых, люди, хорошо знавшие острова, замечали различия между организмами (например, гигантскими черепахами), обитающими на разных островах. Матросы, которые приносили этих черепах на борт корабля и сохраняли для пополнения запаса свежего мяса на время плавания в открытом морс, могли по одному их внешнему виду сказать, на каком острове обитает каждая черепаха. Если галапагосские черепахи были созданы одновременно, почему же они не все одинаковы?
Рис. 28-2. Острова Галапагос — небольшая группа вулканических островов в 950 км от берегов Эквадора. Со времени образования (начавшегося несколько миллионов лет тому назад) они время от времени заселялись растениями и животными, случайно заносимыми ветром или водой с материка. Некоторые из этих организмов сумели выжить, размножились и приспособились к жизни на этих суровых островах.
Дарвин стал задавать себе вопрос, нс произошли ли все черепахи и другие необычные животные и растения островов Галапагос в разнос время от тех организмов, которые существовали на южноамериканском материке? Достигнув этого отдаленного архипелага, они могли постепенно расселиться с одного острова на другой, мало-помалу изменяясь в соответствии с местными условиями и став, наконец, отдельными расами, легко различаемыми наблюдателем.
В 1838 г., вернувшись из своего исторического путешествия, Дарвин прочел книгу английского священника Томаса Мальтуса, озаглавленную «Опыт закона о населении». В этой книге, опубликованной в 1798 г., за 11 лет до рождения Дарвина, было высказано первое предупреждение о непомерном росте населения земного шара. Дарвин увидел, что доводы Мальтуса теоретически верны не только для человеческой популяции, но и для популяций других организмов. Например, одна пара слонов, размножающихся медленнее других животных, за 750 лет может создать популяцию, насчитывающую около 19 млн. особей, если все поколения будут жить достаточно долго и приносить потомство. Несмотря на это численность слонов на Земле остается постоянной; там, где 750 лет назад обитали две особи, их две и теперь. Но что определяет, какие два слона из возможных 19 млн. выживут?
Процесс, с помощью которого и происходит этот выбор. Дарвин назвал естественным отбором. Он использовал это понятие для противопоставления искусственному отбору,которым пользуются селекционеры, преднамеренно изменяющие признаки сортов культурных растений и пород домашних животных так, как это им нужно. При этом для дальнейших скрещиваний оставляют только тех особей, которые обладают желаемыми признаками. Дарвин заметил, что дикие организмы тоже изменчивы. Некоторые обладают признаками, позволяющими им производить больше потомства, чем другим, в тех же естественных условиях; в таком случае их признаки становятся более обычными в популяции, чем признаки особей, производящих меньше потомков. В конце концов эта тенденция может приводить к постепенным, но неуклонным изменениям частоты тех или иных признаков в популяциях.
При искусственном отборе селекционер может направить свои усилия на закрепление одного или нескольких, представляющих интерес признаков, например, величины плода. Однако при естественном отборе организм в целом должен приспособиться ко всем условиям окружающей среды, в которых он обитает. Разумно ожидать, что этот процесс требует длительного времени, и не случайно, что труды геолога Чарльза Лайеля, который утверждал, что Земля имеет гораздо больший возраст, чем считалось ранее, оказали глубокое влияние на Дарвина. Дарвину нужна была именно такая планета, которая могла быть ареной событий, в результате которых возникло существующее разнообразие живых организмов. Обнаружение разнообразных ископаемых остатков, с увеличением возраста все более и более отличавшихся от современных видов, тоже можно считать важнейшей предпосылкой возникновения теории эволюции. Процесс естественного отбора вскоре был признан учеными как основа для объяснения эволюции живого мира.
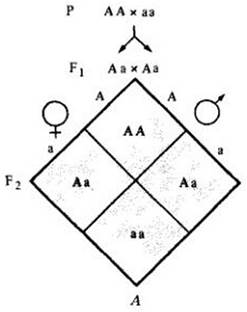
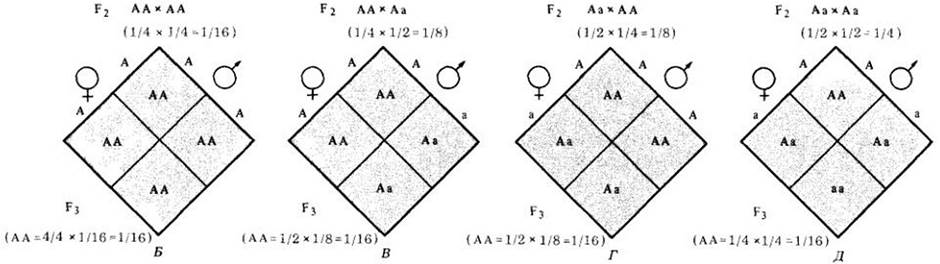
Рис. 28-3. А. Если две популяции, одна гомозиготная по доминантной аллели (АА), а другая гомозиготная по рецессивной (аа), скрещиваются между собой, все первое поколение (F1 будет похоже на доминантных родителей, даже если потомство гетерозиготно (Аа). Однако, когда в поколении F1 произойдут скрещивания, в поколении F2 произойдет менделевское (фенотипическое) расщепление 3:1 (в среднем 1/4 АА, 1/2 Аа и 1/4 аа). При случайных скрещиваниях в поколении F2 вероятность появления особей А А в поколении F3 составит (Б) при скрещивании особей А А — 4/4 (поскольку все потомство в F3 должно быть АА) умножить на 1/16 (потому что вероятность скрещивания АА с АА равна 1/4x114), т. е. 1/16; В — при скрещивании особей АА с Аа вероятность появления особей АА составит 1/2 х 1/8, или 1/16; Г — если особи Аа скрещиваются с особями АА, то вероятность появления особей АА снова будет 1/16; Д — если особи Аа скрестятся между собой, вероятность появления особей АА опять составит 1/16. Таким образом, в третьем поколении (F3) число особей АА составит в среднем 1/16 + 1/16 + 1/16 + 1/16, или 1/4, т. е. столько же, сколько и во втором поколении. Иначе говоря, половая рекомбинация не меняет соотношения различных аллелей в популяции
Поведение генов в популяциях
Закон Харди — Вайнберга
В XIX в., когда большинство биологов признавало своего рода смешанное наследование (т. е. смешение родительских признаков у потомков), было трудно понять, почему редкие признаки не «разбавляются» вплоть до окончательного исчезновения. Дарвин не мог решить эту проблему, потому что в его время мало было известно о механизмах наследственности.
После того как были переоткрыты законы Менделя (см. гл. 9), снова встал этот вопрос, выраженный в более современных понятиях: почему доминантные аллели в конце концов, не вытесняют рецессивные с последующей потерей изменчивости в целом? Этот вопрос чрезвычайно важен, поскольку касается сохранения наследственной изменчивости, лежащей в основе эволюции. Ответ на него, хотя и не сразу очевидный, заключается в правильном понимании дискретной природы гена. Соответствующие вычисления были одновременно представлены в 1908 г. английским математиком Г. X. Харди и немецким врачом Г. Вайнбергом.
Закон Харди — Вайнберга (как его теперь называют) гласит, что в крупных популяциях, где происходит свободное скрещивание, при отсутствии сил, изменяющих соотношение аллелей (что будет обсуждаться ниже), первоначальное соотношение доминантных и рецессивных аллелей будет сохраняться из поколения в поколение.
Для примера рассмотрим аллели одного гена — например гена А. Создадим искусственную популяцию, так что половина особей — гомозиготы АА, а другая половина — гомозиготы аа. На рис. 28-3 показано, что в третьем поколении (или в четвертом, или в пятом) доля особей АА (или аа, или Аа) останется в среднем такой же, какой была во втором поколении.
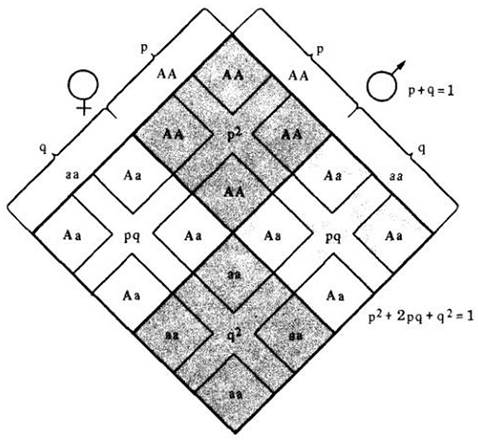
Для исследований генетики популяций элементы закона Харди —Вайнберга обычно выражаются в алгебраических терминах, где дроби, использованные на рис. 28-3, записываются как десятичные. Для гена, имеющего в генном пуле две аллели, сумма частоты (р) доминантной аллели и частоты (д) рецессивной аллели должна равняться 1; р+q =1. (Частота аллели, это просто доля аллели в генном пуле, отнесенная ко всем аллелям того же гена.) Иначе можно сказать, что если имеются только две аллели А и а определенного гена и если половина (0,5) аллелей в генном пуле — А, то другая половина должна быть а. Аналогично если 99 из 100 (или 0,99) — А, тогда 1 из 100 (или 0,01) будет а.
Как тогда мы найдем относительную долю особей АА, Аа и аа? Эти доли могут быть вычислены умножением частоты мужских особей А на частоту женских А [А2]; мужских А — на женских а [Аа]; мужских а — на женских А [Аа]; мужских а — на женских а [а2] или в алгебраической форме: р2+2рq+q2. Это уравнение является разложением бинома (Р+q)2, а если Р+1=1, тогда (р+q)2=1. Таким образом, если половина (0,50) генного пула — А и половина — а, тогда доля АА будет 0,25, доля Аа — 0,50 и доля аа — 0,25. Это полностью соответствует законам Менделя и только выражено в математических терминах:
А2+2Аа+а2 = (0,50)2+2(0,50)(0,50)+(0,50)2 = 0.25+2(0,25)+0,25=0,25+0,50+0,25 = 1.
Что произойдет в последующих поколениях? Как можно видеть из рис. 28-4, частота генотипов остается постоянной. Аналогично и частота генов остается постоянной при р(А)= =0,5 и q(а)=0,5. Таким образом, можно сказать, что закон Харди—Вайнберга предсказывает состояние «генетического равновесия».
Рис. 28-4. Возможные комбинации гамет в популяции, состоящей из особей с генотипами АA, Аа и аа, иллюстрирующие закон Харди — Вайнберга
Величина популяции. Закон Харди — Вайнберга действует только в больших популяциях. В маленьких популяциях случайные потери одного или нескольких отдельных генотипов — если, например, не состоится скрещивание, — могут привести к элиминации одной или большего числа аллелей в популяции.
Миграция. Дальнейшие нарушения вызываются миграцией в популяцию или из нее. Если особи с конкретными генетическими признаками покидают популяцию или входят в другую, с иным соотношением этих признаков, то частота отдельных аллелей и генотипов в обеих популяциях изменяется. Миграция характерна как для животных, так и для растений, в последнем случае посредством, например, распространения семян.
Мутирование. Если отдельный ген мутирует в другую аллельную форму с частотой, превышающей таковую для обратного процесса, то частота таких аллелей в популяции будет меняться.
Отбор. Отбор — это термин, употребляемый для обозначения неслучайного воспроизведения генотипов. В любой изменчивой популяции одни особи оставляют больше потомства, чем другие. В результате определенные аллели оказываются более распространенными в популяции, а другие — менее. Отбор — главный фактор, вызывающий отклонение от равновесия Харди — Вайнберга; он — и первопричина эволюционных изменений. Мутации, конечно, лежат в основе изменчивости организмов, но изменения в популяциях происходят в результате действия отбора на изменчивость, обусловленную мутациями.
Хотя отбор иногда считают созидающей силой, важно помнить, что это только регистрация явлений, которые уже произошли. Когда в данном поколении доля определенных аллелей выше, чем в предшествовавшем, и других объяснений этому нет, — говорят, что произошел отбор. В действительности отбор не является причиной изменений, происходящих при данной совокупности условий. Постоянно элиминируя из популяции определенные аллели, отбор направляет ход изменений признаков, возникших в результате мутаций и рекомбинаций. В итоге в последующих поколениях будет больше особей, лучше выживающих в данных условиях, чем подвергающихся элиминации. В необычных или экстремальных условиях в результате отбора может быть повышена численность генотипов, которых было меньше в основной части популяции. Что касается вредных рецессивных аллелей, даже приводящих в гомозиготном состоянии к гибели особи, то чем ниже их частота в популяции, тем слабее будет воздействие на них отбора. Такая взаимосвязь существует потому, что доля рецессивных аллелей в гомозиготном состоянии резко убывает по мере снижения частоты аллели в популяции. Короче говоря, чем ниже частота рецессивной аллели, тем менее ее гомозиготная форма доступна действию отбора. По мере падения частоты рецессивной аллели вероятность ее удаления из популяции уменьшается.
Реакция на отбор
Генетические факторы
Реакция популяции на отбор происходит на основе генетических законов (это обсуждалось в гл. 9). В принципе отбирается только фенотип, а взаимодействие фенотипа и окружающей среды определяет репродуктивный успех организма. Вследствие того, что почти все признаки естественных популяций детерминируются взаимодействием многих генов, фенотипически сходные особи могут обладать сильно различающимися генотипами. Когда какой-нибудь признак, например, высокий рост, подвергается строгому отбору, обычно происходит накопление аллелей, ответственных за выражение данного признака, и элиминация тех аллелей, которые этому препятствуют.
Однако отбор на полигенный признак — не просто накопление одного набора аллелей и элиминация других. Взаимодействия генов, например, эпистаз или плейотропия (см. гл. 9), во многом определяют, как пойдет отбор в популяции. Некоторые влияния одной аллели на другую могут быть полезными для данной особи или участвовать в формировании признака, который будет подхвачен отбором. Другие, напротив, могут оказаться вредными или работать против подлежащего отбору признака. Вследствие генетических взаимодействий фенотипические эффекты отдельных аллелей могут быть оценены только в конкретном генетическом окружении. Отбор по конкретным аллелям изменяет характер проявления других аллелей в популяции, причем взаимные влияния отбираемых аллелей друг на друга постепенно меняются. Таким образом, значение аллелей в детерминации отдельных признаков или приспособленности организма как целого также меняется.
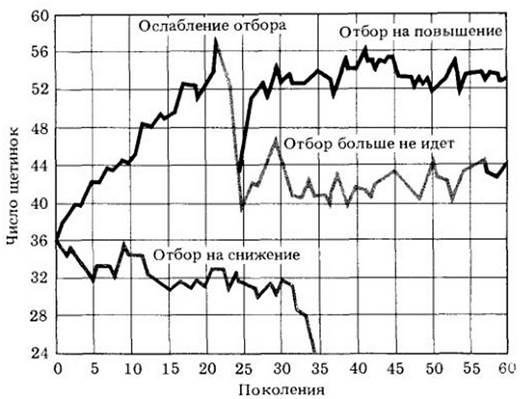
Другой важной стороной природы селективного изменения является необходимость создания «работающего» организма. Строгий отбор по одному признаку может не идти потому, что приведет к накоплению такого количества побочных эффектов, что организм не будет способен к выживанию и воспроизведению. Данные, полученные на основе опытов по разведению плодовой мушки, представленные на рис. 28-5, свидетельствуют о том, что в результате отбора популяция достигает нового внутреннего генетического баланса. При этом возможно появление фертильных и по-своему нормальных особей с 42 абдоминальными щетинками вместо 36. Баланс, который может рассматриваться как мера устойчивости популяции к генетическим изменениям, назван М. Лернером из Калифорнийского университета в Беркли генетическим гомеостазом. В физике гомеостаз — это своего рода динамическое равновесие, т. е. устойчивость к изменениям. Генетический гомеостаз стремится сохранить популяцию как целое путем создания высокой доли особей, хорошо приспособленных к данным условиям.
Рис. 28-5. Эксперимент, в котором число абдоминальных щетинок у мухи Drosophila melanogaster подвергалось искусственному отбору. В двух родительских линиях среднее число щетинок на вентральной поверхности брюшка составляло 36. В одной группе скрещиваний отбор велся на повышение числа щетинок, а в другой — на снижение их числа. В первом случае число щетинок достигло 56 за 21 поколений, но вскоре наступила стерильность. Отбор был прерван после 21 -го поколения и возобновлен после 25-го. Теперь прежнее высокое число щетинок восстановлюсь без потери плодовитости. Во втором случае не удалось достичь нового генетического баланса, и данная группа прекратила свое существование из-за стерильности. Отбор по одному «нейтральному» признаку вместе с генами, от которых зависит проявление данного признака, несомненно, оказывал сильное влияние на генетическую структуру организмов
Фенотипические факторы
Иного рода ограничения отбора связаны с тем, что организмам приходится сталкиваться с противоречивыми потребностями. Например, длинный, ярко окрашенный хвост самца павлина способствует брачному успеху птицы. Поэтому для нее обладание длинным и как можно более красочным оперением — это очевидное преимущество. С другой стороны, птица должна быть защищена от своих врагов, чему отнюдь не способствует яркое хвостовое оперение. Естественный отбор находит некоторое равновесие между двумя селективными направлениями, и павлины, обладающие слишком коротким и тусклым хвостовым оперением или слишком длинным и ярким, нс оставят столько же потомства, сколько птицы с хвостом, «удобным» во всех отношениях.
Еще аналогичный пример. В суровых альпийских условиях растения должны расти и фотосинтезировать быстро, чтобы успеть запасти достаточно углеводов на длинную и трудную зиму. Поэтому весной они должны реагировать возобновлением метаболической активности на появление самых первых благоприятных для роста условий. Однако если реакция будет настолько тонкой, что растения отреагируют даже на временные оттепели в середине зимы, то они элиминируются из популяции. В подобном же положении находятся пустынные растения: их семена не должны прорастать, пока не наступят благоприятные для выживания проростков условия.
Интересную эволюционную стратегию подметил Д. Джэнзен из Пенсильванского университета среди тропических древесных представителей семейства бобовых (Fabaсеае). В Центральной Америке он обнаружил две группы видов: у одних было мало крупных семян, у других — много мелких. Эти растения ежегодно могли уделять ограниченное количество запасных веществ на образование семян и поэтому образовывать либо много мелких, либо мало крупных. Почему одни виды пошли по первому пути, а другие по второму?
Крупные семена предоставляют проросткам более обильную пищу. Поскольку каждое родительское дерево или куст в среднем замещается только одним деревом или кустом, то, казалось бы, для них более выгодно производить мало крупных семян с обильным запасом питательных веществ. Но в тропиках это не всегда возможно из-за очень интенсивного поедания семян бобовых насекомыми, особенно жуками зерновками из семейства Bruchidae. Взрослые жуки откладывают свои яйца в плоды, а их личинки завершают свое развитие внутри семян. Этих жуков часто бывает так мною, что они фактически уничтожают полностью весь ежегодный урожай семян отдельных видов бобовых, кроме тех немногих семян, которые успели, например, унести птицы, прежде чем жуки обнаружили плоды и набросились на них. Джэнзен изучал распространение жуков-зерновок среди крупносемянных и мелкосемянных групп бобовых. К своему удивлению, он обнаружил, что мелкосемянные виды сильнее атаковались жуками, так что оставалось лишь немного жизнеспособных семян, тогда как почти все крупносемянные виды оставались нетронутыми. Затем он выяснил, что именно крупные семена, а не мелкие содержат химические вещества, которые, очевидно, защищают их от жуков.
На основании полученных результатов становится ясно, что перед бобовыми стоял выбор (с эволюционной точки зрения) одной их двух возможностей: либо производить очень много мелких семян с относительно ограниченным запасом питательных веществ для проростков, но с большими шансами на то, что какие-то семена не будут съедены жуками; либо производить крупные семена в меньшем количестве с хорошим обеспечением проростков. Однако выбор последней альтернативы будет успешен только в том случае, если семена будут как-нибудь защищены от поедания жуками. Это пример своего рода эволюционного компромисса, что часто встречается в природе.
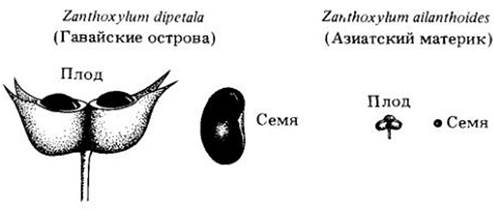
В свете разумного объяснения ситуации с тропическими бобовыми, сделанного Джэнзеном, приобретает интерес то обстоятельство, что семена и плоды растений на океанических островах часто оказываются крупнее, чем у родственных видов на материке (рис. 28-6). Виды вредителей, поражающих растения материка, часто отсутствуют на островах. Вполне вероятно, что тем самым растениям предоставляется благоприятная возможность производить более крупные плоды и семена, которые создают лучшие условия для начального развития проростков, поскольку содержат больше запасных веществ. Такая изоляция от вредителей (так же, как и биохимическая защита крупносемянных бобовых тропической Америки) является фактором, позволившим отбору на крупносемянность, осуществляющемуся на иной основе, достичь здесь большего успеха, чем где-нибудь в другом месте.
Рис. 28-6. Островные виды, изолированные от своих естественных врагов, могут образовывать более крупные плоды и семена. Показанные здесь плоды и семена принадлежат двум родственным видами Zanthoxylum из семейства рутовых (Rutacea); один из них найден на Гавайях, другой — на азиатском материке
Изменения в естественных популяциях
В последние годы было проведено много работ по изучению изменений признаков в природных популяциях. Климатические изменения и естественные катастрофы были постоянными явлениями в мире с самого начала его возникновения, и популяции организмов, существовавшие в ранние исторические эпохи, реагировали на эти изменения так же, как и теперь. Природа и скорость некоторых изменений таковы, что мы можем говорить о них как об «эволюции в действии». Неудивительно, что многие из относительно быстрых изменений, наблюдавшихся в течение последнего столетия или около того, обусловлены деятельностью человека, потому что в последние столетия именно практическая деятельность людей стала экологически доминирующим фактором, не имеющим аналога в истории жизни на Земле.
Некоторые из наиболее ярких эволюционных изменений произошли в популяциях бактерий, у которых имеется только одна хромосома (т. е. они гаплоидны). Мутации бактерий немедленно проявляются в фенотипе и сразу же подвергаются действию отбора. У всех гаплоидных организмов происходит быстрая смена поколений, что, вероятно, существенно для успешных изменений, поскольку здесь нет запаса изменчивости для рекомбинаций, как у диплоидных организмов.
У растений мощный отбор приводит к быстрым изменениям естественных популяций. Например, в Мэриленде растения той части пастбища, где производился выпас, были гораздо ниже растений другой части, где выпаса не было. Считали, что это прямой результат потравы. Для проверки этой гипотезы низкие растения пересаживались в иные условия. Предполагалось, что при отсутствии потравы они в скором времени станут высокими, такими же, как растения той части пастбища, где скот не выпасался. С некоторыми растениями так и произошло, но клевер ползучий (Trifolium repens), мятлик луговой (Роа pratensis) и ежа сборная (Dactylis glomerata) оставались низкими, т. е. в этих популяциях под селективным воздействием выпаса скота произошли генетические изменения. Аналогичный пример представлен на рис. 28-7.
Рис. 28-7. Prunella vulgaris — растение семейства губоцветных, широко распространенное в лесах, на лугах и газонах в умеренной климатической зоне. Большинство популяций представлено прямостоячими растениями (A), которые растут на открытых, часто сыроватых и заросших травой местах, повсеместно в районах с прохладным климатом. Однако газонные популяции всегда представлены стелющимися формами (Б), растущими, например, в Беркли (Калифорния). Прямостоячие растения Prunella vulgaris не могут выжить на газонах, потому что повреждаются при вытаптывании и не способны отрастать за счет нижних ветвей, что необходимо для выживания. Когда газонные растения выращиваются на опытных участках, некоторые из них остаются стелющимися, тогда как другие растут прямо. Стелющаяся форма роста детерминируется генетически в первом случае и внешними условиями — во втором

В Уэльсе отвалы вокруг заброшенных свинцовых рудников богаты свинцом и почти лишены растительности. Один вид злака (Agrostis tenuis) заселил почвы около рудников, которые могли содержать до 1% свинца и 0,03% цинка. Был проведен такой опыт: группы растений Agrostis tenuis с рудничных отвалов и соседних пастбищ выращивались вместе на обыкновенной почве и на взятой около рудника. На обыкновенной почве растения A. tenuis, росшие прежде около рудника, росли явно медленнее и были меньше, чем растения с пастбищ. Однако на почве с рудника они росли нормально, а растения с пастбищ не росли совсем. Половина пастбищных растений погибла через три месяца и имела уродливые корни, которые редко достигали 2 мм в длину. Однако несколько растений с пастбищ (3 из 60) обнаружили некоторую устойчивость к воздействиям почвы, богатой свинцом. Они, несомненно, были похожи на растения, в свое время давшие устойчивую к свинцу линию A. tenuis. Руднику было менее 100 лет, так что устойчивая к свинцу раса образовалась за относительно короткий промежуток времени. Растения отбирались из генетически разнообразных популяций соседних местообитаний. Под действием отбора возникла отдельная раса.
Бесполое размножение и эволюция
При бесполом размножении потомство идентично своему единственному родителю. У растений есть много способов бесполого размножения — от развития неоплодотворенной яйцеклетки до разделения родительского организма на отдельные части. Однако во всех случаях новые растения образуются в результате митоза и поэтому генетически идентичны родительскому.
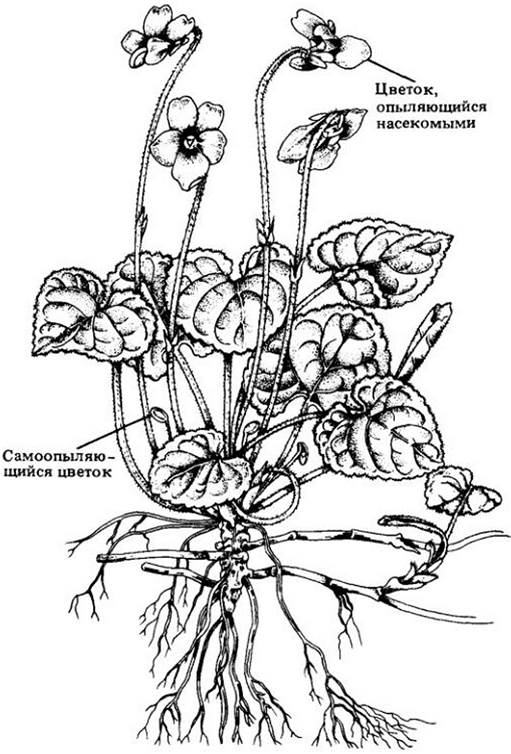
Растения часто размножаются и половым, и бесполым путями, «страхуя» свой эволюционный успех (рис. 28-8), но многие виды размножаются только бесполым путем. Однако и в этом случае ясно, что их предки были способны к половому размножению. Таким образом, вегетативное размножение — это альтернативный «выбор», сделанный под давлением отбора, когда требовалось единообразие популяций. Этот выбор сильно ограничивает способность популяций приспосабливаться к изменяющимся условиям. Однако в разнообразных местообитаниях можно встретить различные популяции одного вида с бесполым воспроизведением. Обладая постоянными признаками, они могут резко отличаться одна от другой и быть лучше приспособленными к росту в конкретных условиях.
Рис. 28-8. Фиалки размножаются половым и бесполым путями. Более крупные цветки перекрестно опыляются насекомыми, и семена могут разноситься на некоторое расстояние ветром или водой. Более мелкие цветки, расположенные ближе к земле, самоопыляются и никогда не раскрываются. Семена таких цветков высыпаются вблизи родительского растения; из них вырастают фиалки, генетически сходные с родительским организмом. Предполагается, что им чем-то лучше расти около материнского растения. Оба указанных выше способа размножения — половые, включающие рекомбинацию. Бесполое размножение фиалок осуществляется ползучими горизонтальными побегами — столонами или корневищами — дающими начало новым, генетически идентичным растениям
Дивергенция популяций
Распространение отдельных видов растений и животных по ареалу никогда не бывает сплошным. Местообитания видов так или иначе разделяются озерами и ручьями, вершинами гор, освещенными солнцем полянами в лесу или участками почвы определенного состава, а раз это так, что перемещение аллелей из одной изолированной популяции в другую более или менее ограничено, и потому популяции могут по- разному реагировать на требования отбора в соответствии с конкретными условиями обитания. Даже если бы нормальный поток генов (т. е. перемещение аллелей из одной популяции в другую вследствие миграции особей) мог предоставить источник изменчивости отдаленным популяциям, то и тогда он не смог бы полностью нейтрализовать эффекты отбора в рамках местных условий.
Действительное разобщение популяций одного и того же вида зависит от расстояний, на которые данные растения способны расселяться, однако эти расстояния часто бывают очень малы. Для некоторых насекомоопыляемых растений, произрастающих в областях умеренного климата, 300 м может быть достаточно, чтобы надежно изолировать две популяции. Редко более 1% пыльцы, попавшей на данное растение, приносится издалека. У ветроопыляемых растений при нормальных условиях очень мало пыльцы переносится на расстояние свыше 50 м от родительского растения. Для пыльцы таких растений вероятность достигнуть восприимчивого рыльца растения, находящегося на далеком расстоянии, тем меньше, чем больше это расстояние. Не следует, конечно, говорить, что две сосны, отстоящие друг от друга на 50 м, не имеют генетического контакта. Вернее, будет сказать, что на потомках больше будут сказываться требования местных условий, чем аллели, поступившие издалека.
Любая пара разобщенных популяций будет расходиться в своих признаках вследствие различной направленности отбора. Если восстановится их контакт, то они или сольются, или накопившиеся между ними различия приведут к некоторой генетической изоляции. Генетическая, или репродуктивная, изоляция происходит разными путями, о чем речь будет идти ниже.
Экотипическая и клинальная изменчивость
Пластичность развития — это тенденция особей с течением времени изменяться в ответ на изменение условий окружающей среды (при этом между генетически однородными организмами могут возникать определенные различия). Такая пластичность более характерна для растений, чем для животных, поскольку свойственная первым открытая система роста гораздо легче может быть тем или иным образом изменена, и вследствие этого отдельные растения поразительно различаются между собой.
Каждый садовник знает, что факторы внешней среды могут вызвать глубокие изменения фенотипа у различных видов растений. Листья, развивающиеся в тени, тоньше и шире; они имеют воздушные полости большего объема, более тонкий слой палисадной ткани и меньше устьиц, чем листья того же растения, развивающиеся на солнце (см. с. 60). На форму листьев может оказывать влияние и продолжительность дня, т. с. та часть 24-часового периода, в течение которого растение находится на свету. У Kalanchoe, например, растения, выросшие в условиях короткого дня (8 ч света), имеют мелкие суккулентные листья с гладкими краями, тогда как растения, выросшие в условиях длинного дня (16 ч света), имеют широкие тонкие листья с выемчатыми краями. В свете этих наблюдений неудивительно, что примерно до 1930 г. многие ученые предполагали, что большая доля изменчивости, наблюдаемая в природе, обусловлена прямым воздействием внешних условий и нс имеет под собой генетической основы.
Как же контролируются различия между расами растений, произрастающих в различных местах, — генетически или посредством внешних условий? Первый определенный ответ на этот вопрос в 20-х гг. дал шведский ботаник Гёте Турессон. Среди растений многих видов, произрастающих на юге Швеции, он обнаружил неодинаковые расы. Отдельные растения, представляющие эти расы, он пересадил на свои опытные плантации. Таким образом Турессон исследовал 31 вид. У большинства из них наблюдавшиеся в природе различия находились под генетическим контролем; у очень немногих они были результатом прямого воздействия внешних условий. Такие различия, как рост, время цветения, цвет листьев и т. д., обычно контролировались генетически. Турессон назвал такие расы, образовавшиеся в конкретных местах, экотипами.
В изучение экотипов особый вклад вносят работы Дж. Клозена, Д. Кека и У. Хайзи, проведенные под эгидой Института Карнеги (Вашингтон) в Отделе биологии растений Станфордского университета (Калифорния). Эти исследователи проводили опыты с некоторыми видами растений, характерными для западных областей США. Они организовали станции для пересадки растений в трех пунктах Калифорнии (Станфорд, Матер и Тимберлайн, рис. 28-9). Они расширили опыты Турессона, работая преимущественно с растениями, способными размножаться бесполым путем, так что генетически идентичные особи могли быть выращены во всех трех пунктах.
Природным условиям западных штатов свойственны большие контрасты, и поэтому неудивительно, что многие виды растений представлены здесь резко очерченными экотипами. Одним из видов, изученных группой из Института Карнеги, было многолетнее травянистое растение Potentilla glandulosa, находящееся в близком родстве с земляникой (Fragaria). Оно встречается в различных климатических зонах Калифорнии, и его естественные популяции можно обнаружить вблизи каждой из трех упомянутых опытных станций. Когда растения Р. glandulosa разного экологического происхождения выращивались бок о бок в питомниках, заложенных учеными из Института Карнеги на трех опытных станциях по пересадке растений, то выявился ряд экологических различий между исходными линиями. Можно было четко выделить четыре экотипа Р. glandulosa, морфологические признаки каждого из которых строго соответствовали тем физиологическим реакциям, которые были критическими для выживания экотипов в их природных условиях.
Рис. 28-9. Питомники Института Карнеги в Калифорнии. А — в Станфорде, близко к уровню моря; Б — Матер, в центральной части гор Сьерра-Невада на высоте около 1400 м; В — Тимберлайн, на высоте 3050 м

Например, экотип Берегового хребта представлен растениями, которые в Станфорде и летом и зимой росли хорошо. Они выживали и в Матере, где подвергались воздействию зимних холодов на протяжении почти пяти месяцев. Здесь эти растения впадали в состояние зимнего покоя, но за вегетационный период успевали накопить достаточно запасных веществ, чтобы пережить продолжительный неблагоприятный сезон. В Тимберлайне растения этого экотипа почти неизменно погибали в первую же зиму; короткий вегетационный период на этой высокой отметке нс позволял им накопить достаточно запасных веществ. Растения с калифорнийского Берегового хребта, относящиеся к другим видам, образовывали экотипы со сходными физиологическими реакциями. В самом деле, часто бывает так, что линии совершенно разных видов, но растущих вместе в данных условиях, физиологически более сходны друг с другом, чем с растениями других популяций своего же вида.
Физиологические и морфологические признаки экотипов обычно имеют очень сложную генетическую основу, включающую десятки (а в некоторых случаях, вероятно, даже сотни) генов. Отчетливые экотипы характерны для тех областей, где местообитания тоже резко разграничены. С другой стороны, если условия меняются постепенно, без резких переходов, то и признаки растений соседних популяций могут меняться так же плавно. Такого рода постепенная смена признаков называется клином.
Клины часто встречаются у обитателей тех районов моря, где температура воды повышается или понижается постепенно, при изменении широты. Они характерны и для организмов, населяющих области, подобные восточной части США, где градиенты количества осадков могут распространяться на тысячи километров. Если образцы растений соседних популяций собирать вдоль клина, то различия часто бывают пропорциональны расстоянию между популяциями. Так же, но в меньшем масштабе изменяются сами популяции растений — либо постепенно, либо резко в зависимости от местных условий.
Физиологическая дифференциация
Чтобы понять, почему экотипы процветают в том или ином месте, надо понять физиологическую основу их экотипической дифференциации. Например, скандинавские линии золотарника (Solidago virgaurea) из затененных и освещенных местообитаний в условиях эксперимента обнаруживают различия в фотосинтетической реакции на интенсивность освещения во время роста. Растения из затененных мест растут быстро при низкой освещенности, тогда как скорость их роста заметно замедляется при высокой, и напротив, — растения из освещенных местообитаний растут быстро в условиях сильной освещенности и гораздо хуже при слабой.
В другом эксперименте изучались линии арктических и альпийских популяций травянистого растения Oxyria digyna с большим широтным диапазоном распространения — от Гренландии и Аляски на севере до Калифорнии и Колорадо на юге. Растения северных популяций содержали больше хлорофилла и отличались большей интенсивностью дыхания при всех температурных условиях по сравнению с южными растениями. У высокогорных растений из мест, близких к южным пределам распространения вида, при высоком уровне освещенности в условиях опыта фотосинтез был более интенсивным, чем у равнинных растений Крайнего Севера. Таким образом, каждая данная раса может наилучшим образом функционировать в своих собственных местообитаниях, какими бы ни были те условия — высокий уровень освещенности в высокогорьях или низкий на Крайнем Севере и т. д. Распространение Oxyria digyna по всему столь обширному ареалу и в столь широком диапазоне экологических условий стало возможным отчасти благодаря тому, что входящие в состав вида популяции различаются по своему метаболическому потенциалу.
Репродуктивная изоляция
Генетическая система популяции растений реагирует на отбор как единое целое. По мере того как отдельные популяции начинают все более и более отличаться друг от друга, признаки, ранее не препятствовавшие им скрещиваться между собой, могут измениться вместе с признаками, чей полезный эффект проявляется незамедлительно, например, с позволяющими этим популяциям хорошо расти в различных местах. По этой причине растения из сильно различающихся популяций могут утратить способность образовывать гибриды (т. с. потомство генетически несходных родителей), а если они и получаются, то часто оказываются бесплодными. Вообще, чем больше различий обнаруживают две популяции, тем меньше вероятность того, что они окажутся способными образовать гибридное потомство.
Когда две популяции становятся репродуктивно изолированными (т. е. неспособными к образованию плодов- гибридов), они больше не оказывают влияния на эволюцию друг друга, по крайней мере в генетическом отиошении. Поэтому такая изоляция — один из наиболее критических моментов в эволюционной дивергенции популяций и видообразовании.
Репродуктивная изоляция — основной критерий вида. Он, однако, не всегда применим, поскольку у некоторых групп растений очень разные внешне и экологически различные виды (в частности, у таких долгоживущих растений, как деревья и кустарники) иногда могут образовывать плодовитые гибриды. В противоположность этому гибриды между видами травянистых растений часто стерильны или образуются с трудом; подобная дифференциация нередко происходит даже между популяциями короткоживущих растений. Гибриды между такими популяциями могут быть полностью или частично стерильными. Поскольку между выделенными для различных групп растений видами имеются различия по генетическим признакам, возникают трудности с определением понятия вид. Очевидно, что для наиболее приемлемого решения вопроса о том, как классифицировать ту или иную группу организмов, недостаточно использовать только один критерий, но важно учитывать ряд факторов.
При одинаковом давлении отбора популяции однолетних растений, по-видимому, изменяются быстрее, чем популяции долгоживущих. Дело не только в том, что однолетники имеют более короткий жизненный цикл, но также и в том, что их существование зависит от ежегодного воспроизведения семенами, а это уже само по себе является фактором, ускоряющим эффект естественного отбора. В итоге такие популяции дивергируют быстрее. В тех областях, где однолетники способны успешно произрастать — пустынях или местах с сухим летом, как в Калифорнии, — они могут составлять треть и более от общего числа представленных там видов. В целом эти виды гораздо моложе (считая со времени их возникновения), чем виды деревьев, кустарников и многолетних травянистых растений, в сообществе с которыми они находятся.
Кроме стерильности гибридов или невозможности образовать гибридное потомство есть еще ряд факторов, которые способствуют четкому разграничению видов при их совместном произрастании. Прежде всего это факторы, которые предотвращают образование гибридов. Например, растения двух видов, способные к образованию плодовитых гибридов, могут встречаться в одном ареале, но в разных его участках. На востоке США дуб американский шарлаховый (Quercus coccínea) может занимать совместно с дубом бархатным (Q. velutina)обширный ареал. Оба вида опыляются ветром и в искусственных условиях легко образуют плодовитые гибриды. Несмотря на это, в природе их межвидовые гибриды обнаруживаются редко. Дело в том, что первый дуб встречается в относительно влажных низменных местах с кислыми почвами, а второй — на сухих, хорошо дренированных почвах. Только там, где местообитания изменились в результате пожаров или вырубки деревьев, гибриды встречаются чаще. Это можно объяснить тем, что они смогли лучше приспособиться к вновь создавшимся условиям, чем каждый из родителей.
Среди других механизмов, предотвращающих образование гибридов между видами, растущими вместе, важное место занимают сезонные различия во времени цветения. Если два вида нс цветут одновременно в природных условиях, они не образуют гибридов, даже если и растут бок о бок. Фотопериодические механизмы, обсуждавшиеся в гл. 25, смогут способствовать разобщению видов. Кроме того, два вида, встречающиеся вместе, могут отличаться системой опыления (подробно на этом мы остановимся в следующей главе). Если два вида посещаются и опыляются разными насекомыми, то гибрид получится лишь в том случае, если насекомые по ошибке посетят «не тот» цветок.
Группы родственных видов
Рассмотренные нами эволюционные процессы приводят к образованию групп родственных видов в разных географических областях. Такие группы животных на островах Галапагос, особенно упоминавшиеся выше черепахи, а также вьюрки стали знаменитыми благодаря той роли, которую сыграли в создании Дарвином теории эволюции. Видообразование такого типа называется адаптивной радиацией (см. приложение). Дифференциация на островах особенно поразительна тем, что в отсутствие конкуренции здесь, по-видимому, легче образуются формы, весьма необычные по сравнению с родственными видами на континентах. Очевидно, обстановка на островах благоприятствует значительным эволюционным изменениям, подобным тем, которые предшествуют возникновению новых родов и семейств. В таких местах признаки животных и растений способны меняться быстрее, чем на материке; кроме того, могут возникать признаки, которые никогда в других местах не встречаются. Подобные группы видов могут, конечно, возникать и в пределах материковых ареалов, демонстрируя самую неожиданную дифференциацию. Они являются неизбежным следствием обсуждаемых эволюционных процессов.
Роль гибридизации в эволюции
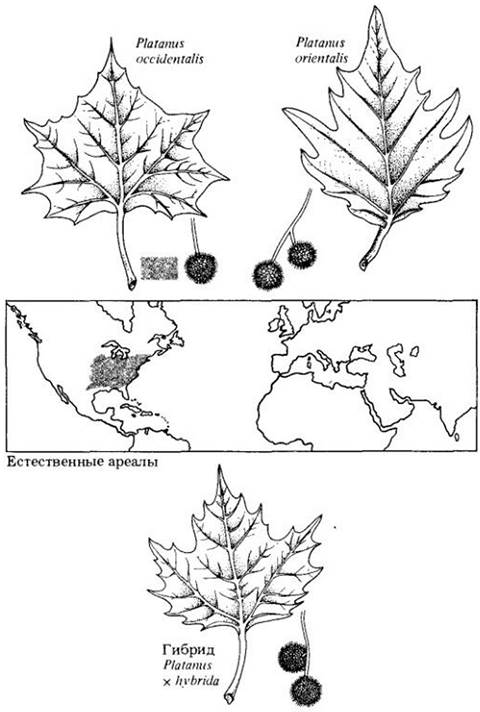
Даже если в природе межвидовые гибриды образуются редко, они могут быть ценны тем, как в них сочетаются родительские признаки. Окружающие условия могут быстро меняться, а особи гибридного происхождения благодаря новым комбинациям генов часто оказываются лучше адаптированными к новым условиям и способными заселить такие местообитания, в которых родительские организмы никогда не смогли бы существовать. В тех случаях, когда местообитания родительских видов расположены в одном ареале, но резко различаются (как и в случае американского шарлахового и бархатного дубов), гибридизация маловероятна. Напротив, там, где местообитания объединены или нарушены, ситуация может сложиться по-разному. Здесь рекомбинация генетического материала, первоначально характеризовавшего два разных вида, может привести к получению более адаптированного потомства, чем изменения, произошедшие внутри одной популяции (рис. 28-10). Признаки гибридов могут стабилизироваться, если они окажутся лучше приспособленными к новым местообитаниям, чем любой из родительских организмов.
Рис. 28-10. Платан (Platanus) образует хорошо различающиеся популяции, сохранившие способность к гибридизации.
Современные виды этого рода разделились около 50 млн. лет тому назад в отдаленных друг от друга местах произрастания. Один из них — восточный платан (P. orientalis) — распространен от восточного Средиземноморья до Гималаев. Это дерево широко разводится в Южной Европе со времен Древнего Рича, но не может расти в Северной Европе, вдалеке от смягчающего влияния моря. После открытия Нового Света один из североамериканских видов (P. occidentalis) был акклиматизирован в более холодных районах Северной Европы, где стал процветать. Около 1670 г. в Англии произошла спонтанная гибридизация этих сильно различающихся видов, где они были посажены вместе. В результате получились промежуточные по признакам и плодовитые платаны (Platanus х hybrida). Эти гибриды способны расти в районах с холодными зимами. Теперь их используют для озеленения улиц в городах умеренной климатической зоны
Межвидовая гибридизация — важный эволюционный механизм у многих групп растений. У некоторых родов рекомбинация генетического материала, по-видимому, представляет собой основной способ образования новых видов, которые могут освоить иные местообитания. Эти роды состоят в основном из деревьев или кустарников, как, например, Eucalyptus, Quercus, Arctostaphylos и Ceanothus (рис. 28- 11). Группы родственных видов более обычны на островах или в экологически разнообразных районах, например, в Калифорнии и соседних с ней штатах. Другой пример, на этот раз с участием опылителей, показан на рис. 28-12.
Рис. 28-11. Образование гибридных популяций — важный эволюционный механизм у многих групп древесных растений, в том числе у рода Ceanothus. А — карта центральной части Калифорнии (обратите внимание на залив Сан-Франциско внизу) — области, сложной в геологическом отношении. Два относительно широко распространенных и обособленных вида: прибрежный — С. gloriosus и населяющий внутренние области — С. cuneatus, границы распространения которого заходят далеко на восток за пределы ареала, изображенного на карте. Данные виды образуют три типа гибридных популяций; каждая изменчива, но стабилизирована и способна расти лучше, чем любой из родительских видов, там, где они произрастают совместно. Листья, характерные для представителей разных популяций, изображены на карте. Б — расщепление в потомстве при искусственном скрещивании родительских видов, внешне резко отличающихся друг от друга; некоторые промежуточные формы бывают похожи на упомянутые ebiuie промежуточные популяции, показанные на карте слева. В — цветущая ветвь С. gloriosus. Г — цветущая ветвь С. cuneatus
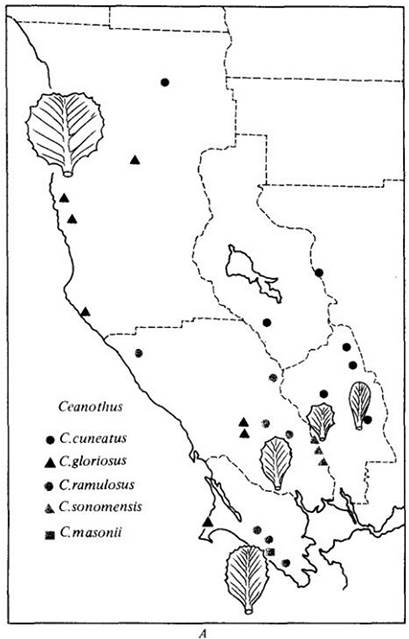
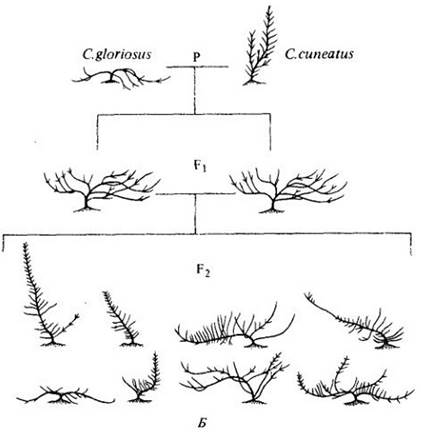
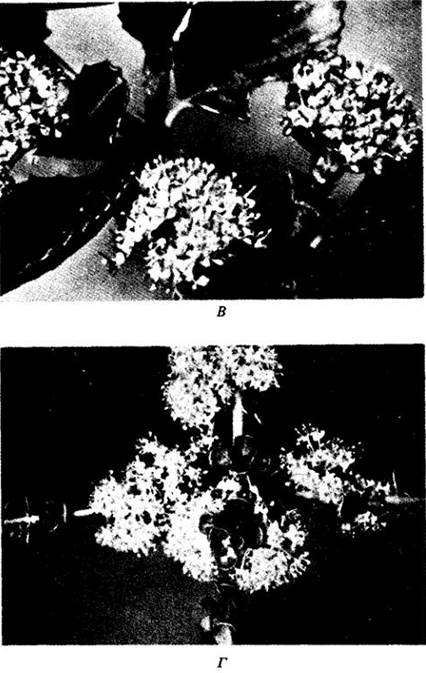
Рис. 28-12. Ричард Строу из Калифорнийского государственного университета в Лос-Анджелесе предположил, что вид Реnstemon spectabils, обнаруженный в горах южной Калифорнии, имеет гибридное происхождение. Для одного из родительских видов (Р. grinnellii) характерны большие двугубые бледно-голубые цветки, которые обычно опыляются крупными пчелами, например, пчелами-плотниками. Другой вид, Р. centranthifolius, имеет длинные тонкие красные цветки, посещаемые преимущественно колибри. Предполагаемый гибрид Р. spectabilis, показанный здесь, занимает промежуточное положение в морфологическом и экологическом отношении и имеет розово-красные цветки. Их опыляют осы из специализированного семейства, собирающие пыльцу. Эти осы не посещают цветки ни одного из родительских видов

Возникновение и стабилизация гибридных популяций зависят от плодовитости гибридов. Однако, даже если гибриды стерильны, они все же могут размножаться либо бесполым путем, либо восстановив свою плодовитость путем полиплоидизации.
Бесполое воспроизведение у гибридов
Стерильные гибриды могут размножаться вегетативным путем. Системы, в которых вегетативное воспроизведение преобладает, но ауткроссинг (т. е. перекрестное оплодотворение между различными линиями) иногда происходит, являются наиболее гибкими. В результате ауткроссинга может произойти гибридизация между обособившимися расами или видами, что в свою очередь, возможно, приведет к новым комбинациям аллелей, которые могут оказаться полезными в данных условиях.
Наглядным примером подобной системы может служить мятлик луговой (Роа pratensis), который в той или иной форме распространен по всему Северному полушарию. Случайная гибридизация в пределах всей группы родственных видов дала сотни апомиктических рас, хорошо приспособленных к экологическим особенностям конкретных мест произрастания. В таких гибких системах новые генотипы постоянно создаются за счет гибридизации, а лучшие из них сохраняются благодаря апомиксису. Апомиктические растения, в том числе Роа, наиболее приспособлены к условиям Арктики, поскольку могут обходиться без опыления насекомыми, которое затруднено из-за небольшой продолжительности вегетационного периода и суровых климатических условий. Кроме того, более стабильные генотипы могут иметь большее преимущество в трудных арктических условиях, чем генотипы, образующиеся в результате рекомбинации (т. е. полового процесса). Таким образом, популяции, возникающие в результате вегетативного размножения или самоопыления, могут иметь двойное преимущество.
Сотни видов боярышника (Crataegus) и ежевики (Rubus) на востоке США представляют собой апомиктические производные групп видов, среди которых случайно образовывались гибриды. Во всех этих случаях нарушение человеком естественных мест произрастания, приобретающее широкие масштабы, приводит к появлению большого разнообразия новых генотипов, которых никогда раньше не было в коренных лесах этих мест.
Полиплоидия
Клетки или особи, имеющие более двух наборов хромосом, называются полиплоидами. Полиплоидные клетки с небольшой частотой возникают в результате «ошибок» митоза, когда хромосомы делятся, а цитокинез нс происходит. Таким образом могут образоваться клетки с удвоенным числом хромосом. Если они, пройдя через интерфазу, будут делиться, то смогут дать начало (половым или бесполым путем) новым особям, клетки которых будут иметь вдвое больше хромосом, чем у родителей. Полиплоидные растения могут быть получены искусственно с помощью колхицина — алкалоида, подавляющего образование митотического веретена в результате нарушения образования микротрубочек.
У полиплоидов изменчивость часто бывает значительно уже, чем у родственных диплоидов, поскольку каждый ген представлен у них по меньшей мере в удвоенном числе. При расщеплении в потомстве гомозиготные по какому-нибудь рецессивному гену особи составят только 1/16 вместо 1/4 у диплоидов. (В обоих случаях принимается, что частота рецессивных аллелей равна 0,50.) Полиплоидам свойственно самоопыление, еще более снижающее их изменчивость, несмотря на то что родственные им диплоиды преимущественно опыляются перекрестно. Одни полиплоиды лучше приспособлены к сухим местам или более низким температурам, чем исходные диплоидные формы, в то время как другие лучше приспособлены к особым типам почв. Благодаря этому они могут заселять места с экстремальными условиями существования, в которых их диплоидные предки скорее всего бы погибли.
С небольшой частотой полиплоиды встречаются во многих естественных популяциях. Полиплоиды легче, чем соответствующие им диплоиды, вступают в неродственные скрещивания. При этом сразу же могут получаться плодовитые гибриды. Реже полиплоиды гибридного происхождения образуются путем удвоения числа хромосом у стерильных диплоидных гибридов; это один из путей восстановления плодовистости.
Именно таким, менее обычным, путем образовались полиплоидные гибриды между редькой (Raphanus sativus) и капустой (Brassica olerácea). Это был первый хорошо документированный случай полиплоидии1. Оба рода принадлежат к семейству крестоцветных (Brassicaceae) и находятся в близком родстве. В соматических клетках и того и другого вида находится 18 хромосом, и в первой метафазе мейоза всегда обнаруживается 9 пар хромосом. С некоторым трудом был получен гибрид между этими растениями. В мейозе он имел 18 непарных хромосом (9 от редьки и 9 от капусты) и был совершенно бесплоден. Среди этих, гибридных растении, спонтанно образовался полиплоид, у которого в соматических клетках было 36 хромосом и в процессе мейоза регулярно образовывались 18 пар. Иными словами, полиплоидный гибрид имел все 18 хромосом как редьки, так и капусты, и они функционировали нормально. Полиплоидный гибрид был довольно плодовитым.
1Плодовитый межродовой редечно-капустный гибрид был получен советским генетиком Г. Д. Карпеченко, пионером в преодолении бесплодия отдаленных гибридов посредством полиплоидизация и, таким образом, первооткрывателем неизвестного прежде пути видообразования. — Прим. перев.
Некоторые полиплоиды возникали как сорняки в местах, связанных с деятельностью человека, и иногда они достигали удивительного процветания. Один из хорошо известных примеров — обитатели соленых болот из рода Spartina. Один из видов, S. maritima, встречается на болотах вдоль берегов Европы и Африки. Другой вид, S. alterniflora, был завезен в Великобританию с востока Северной Америки около 1800 г. и впоследствии широко распространился, образовав крупные локальные колонии.
В Британии растения местного вида S. maritima низкорослы, тогда как S. alterniflora гораздо выше и нередко достигают в высоту 0,5 м, а иногда 1 м и более. Вблизи Саутгемптона оба вида (местный и интродуцированный) произрастали совместно на протяжении всего XIX в. В 1870 г. ботаники обнаружили стерильный гибрид между двумя видами, который хорошо размножался корневищами. Один из родительских видов, 5. maritima, имеет диплоидное число хромосом, равное 60 (2n = 60), другой, S. alterniflora, — равное 62 (2n = 62); гибрид тоже имеет 2n = 62, вероятно, из-за небольшого нарушения мейоза. Этот стерильный гибрид, названный Spartina х townsendii, до сих пор существует. Примерно в 1890 г. естественным путем из него образовался мощный плодовитый полиплоид. Он имеет диплоидное число хромосом 2n = 122 (одна пара хромосом, очевидно, потерялась) и быстро распространился по берегам Великобритании и северо-западной Франции. Его часто сажают для закрепления заболоченных берегов, что способствует еще большему распространению этого растения.
Одной из важнейших полиплоидных групп растений можно считать род Triticum (пшеницы). Самая распространенная в мире хлебная культура — мягкая пшеница (Т. aestivum) — имеет 2n = 42. Мягкая пшеница возникла по крайней мерс 8000 лет назад, вероятно, в Центральной Европе, в результате естественной гибридизации возделываемой пшеницы, имеющей 2п=28, с диким злаком того же рода, имеющим 2n = 14. Дикий злак, вероятно, рос как сорняк среди посевов пшеницы. Гибридизация, давшая начало мягкой пшенице, могла произойти между полиплоидами, появлявшимися время от времени в популяциях обоих родительских видов.
Вполне вероятно, что как только 42-хромосомная пшеница с ее полезными признаками появилась на полях первых земледельцев, они сразу ее заметили и отобрали для дальнейшего культивирования. Одна из ее родительских форм, 28- хромосомная возделываемая пшеница, в свою очередь произошла в результате гибридизации двух диких 14-хромосомных видов с Ближнего Востока. Виды пшеницы, имеющие 2n = 28, и теперь продолжают возделываться наряду с 42-хромосомными. Такие 28-хромосомные пшеницы представляют собой главный источник зерна для производства макарон благодаря высокой клейкости их белка.
Исследования последних лет показали, что новые линии, полученные с помощью гибридизации, могут улучшить сельскохозяйственное производство. Особенно многообещающих! является Triticosecale1 — группа созданных человеком гибридов между пшеницей (Triticum) и рожью (Secale). Некоторые из них, сочетающие урожайность пшеницы с неприхотливостью ржи, наиболее устойчивы к линейной ржавчине — болезни, наносящей большой ущерб сельскому хозяйству (см. рис. 13-37). Эти свойства особенно важны в высокогорных районах тропиков и субтропиков, где ржавчина — главный фактор, лимитирующий культивирование пшеницы. Triticosecale теперь выращивается в больших масштабах и получила широкую популярность во Франции и других странах (см. с. 242). Наибольшую известность имеет 42-хромосомная линия этой зерновой культуры. Она была получена путем удвоения числа хромосом после гибридизации 28- хромосомной пшеницы с 14-хромосомной рожью.
1 В советской литературе более принято иное название этого гибрида — тритикале (Triticale). — Прим. перев.
В природе полиплоиды отбираются под влиянием внешних условий, а не благодаря деятельности человека. Полиплоидия — один из важнейших эволюционных механизмов. Явления, упомянутые выше (см., например, происхождение пшениц), должны были произойти более 100 000 раз, чтобы можно было объяснить то количество полиплоидов, которое в наше время представлено в мировой флоре (более половины всех видов растений). Среди них многие из наиболее важных сельскохозяйственных культур — не только пшеница, но и хлопчатник, сахарный тростник, банан, картофель и подсолнечник. К этому перечню можно добавить большинство красивых садовых цветов — хризантемы, анютины глазки, георгины.
Рис. 28-13. Полиплоидия усиленно изучалась у злаков рода Spartina, растущих на соленых болотах у берегов Северной Америки и Европы. А. Такое болото у берегов Великобритании. Б. Гибрид Spartina. В. Spartina maritima, местный европейский вид соленых болот (2n = 60). Здесь показаны его хромосомы в анафазе I мейоза. Г. Spartina alterniflora, североамериканский вид, 2n = 62 (здесь показаны 30 бивалентов и 2 непарных хромосомы в метафазе I мейоза). В Европе это растение впервые найдено в водоемах у Саутгемптона в 1839 г., но настоящее время его интродукции неизвестно. Искусственные скрещивания двух указанных видов никогда не производились, но их стерильные гибриды — Spartina х townsendii— находили в этих местах в 1870 г. Д. Мощный полиплоид S. anglica произошел спонтанно от этого стерильного гибрида и был впервые найден в начале 1890-х гг. Полиплоид имеет 2n = 122; здесь показана его клетка в первой анафазе мейоза. Данный гибрид теперь распространился по всем соленым болотам Великобритании и другим страниц умеренного климата
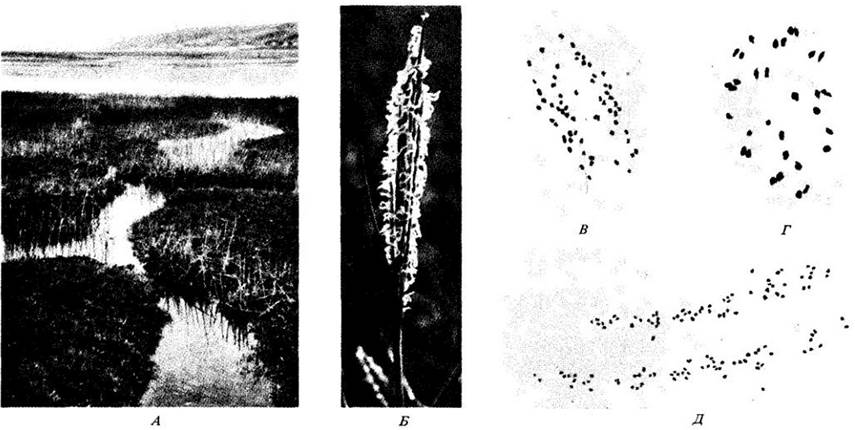
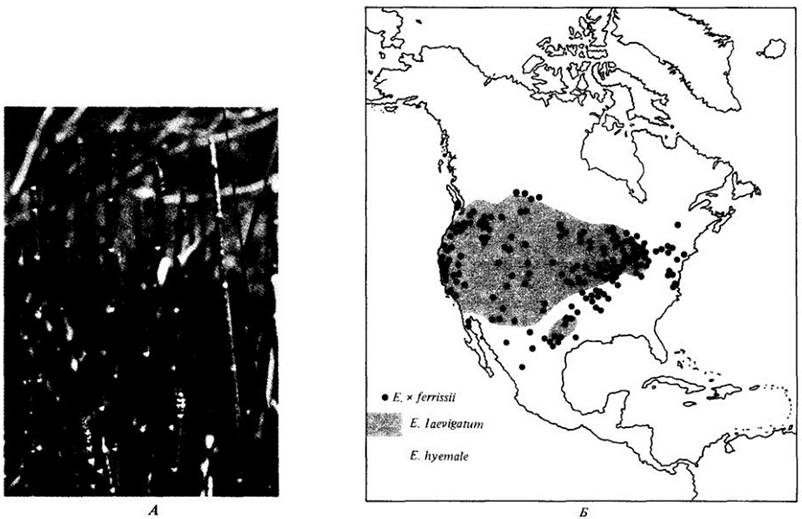
Рис. 28-14. Один из наиболее распространенных и крупных хвощей (см. рис. 26-3, Б) в Северной Америке — Equisetum х ferrissii — это полностью стерильный гибрид Е. hyemale и Е. laevigatum. Хвощи легко размножаются кусочками подземных стеблей, и гибрид поддерживает свое существование по всему обширному ареалу вегетативным размножением. А. Стебли Е. х ferrissii со стробилами. Б. Ареалы Е. х ferrissii и родительских видов
Происхождение таксонов высшего ранга
Как только знания о путях видообразования стали полнее, ученые обратились к вопросу о том, как возникают роды и другие таксоны высшего порядка. Приложение, посвященное флоре Гавайских островов, позволяет понять, что за образование родов ответственны те же процессы, что и за образование видов. Если какой-нибудь вид хорошо приспособлен к тем местообитаниям, которые сильно отличаются от местообитаний его предковой формы, он, вероятно, станет сильно отличаться от предка и может дать начало новым видам и сформировать новую эволюционную линию. Со временем эта линия может быть определена как новый род, семейство или даже класс организмов. Для возникновения таксонов надвидового уровня не требуется никаких иных механизмов — только прерывистость местообитаний и изменение образа жизни.
В последнее время много было написано о том, является ли эволюция всегда постепенным процессом, или она идет скачками, т. е. продолжительные периоды постепенных изменений (или отсутствия изменений) прерываются периодами быстрых изменений. Последняя из этих моделей эволюции называется прерывистым равновесием. Некоторые сторонники этой модели утверждают, что макроэволюция, т. е. процесс образования надвидовых таксонов, совершается на основе иных принципов, а не путем постепенных изменений, что характерно для микроэволюции, которой и была в основном посвящена эта глава. Однако таксоны высшего порядка различаются между собой такими же признаками, что и некоторые виды, поэтому процесс эволюции представляется единым.
Заключение
Естественный отбор — это процесс, благодаря которому организмы, обладающие признаками, наиболее благоприятными для жизни в данных условиях, оставляют больше жизнеспособных потомков. В его основе лежит генетически детерминированная изменчивость особей в естественных популяциях. Отбор вступает в действие, когда организмы начинают заселять новые местообитания. Репродуктивный успех (приспособленность) не может быть успехом вообще, а проявляется только в определенных конкретных условиях обитания популяции.
Основой генетики популяций является закон Харди— Вайнберга. Он гласит, что в больших популяциях, где происходит свободное скрещивание и отсутствуют факторы, влияющие на соотношение аллелей, соотношение доминантных и рецессивных аллелей остается постоянным из поколения в поколение.
Причиной изменения аллелей и отклонений от равновесия Харди—Вайнберга могут быть четыре главных фактора: величина популяции, миграция, мутационный процесс и отбор. Из них наиболее важным является отбор. Он определяется как неслучайное воспроизведение генотипов, когда одни получают преимущество перед другими в тех конкретных условиях, в которых они находятся.
Рецессивные аллели диплоидных организмов малодоступны отбору. Чем меньше становится их частота, тем большая их доля оказывается в гетерозиготном состоянии и фенотипически не проявляется (маскируется).
Реакция популяций на давление отбора по ряду причин сложна. Отбору доступны только фенотипы, а сходные фенотипы могут быть результатом очень разных сочетаний аллелей. Благодаря эпистазу и плейотропии аллели не могут отбираться по отдельности; отбор воздействует только на весь генотип в целом.
Популяции приспосабливаются к конкретным условиям и образуют четкие экотипы, если условия так же четко разграничены. Если же условия меняются постепенно, популяции растений могут образовывать клины по признакам, соответствующим этим условиям.
Изменения, происходящие в популяциях, влияют и на способность к успешному скрещиванию их между собой. После некоторого периода изоляции две популяции могут оказаться несовместимыми или давать при скрещивании только стерильные гибриды. Родственные виды относительно долгоживущих растений, например, деревьев и кустарников, менее подвержены репродуктивной изоляции, чем родственные виды однолетних или других короткоживущих растений.
Гибридные популяции, происшедшие от двух видов, представляют собой обычное явление, особенно среди деревьев и кустарников. Они наиболее характерны для областей, где число видов невелико и приспособление к различным локальным условиям особенно важно (например, для океанических островов), а также для районов, где климатические и другие условия имеют четкие границы, как, например, в Калифорнии.
Даже если гибриды между двумя видами стерильны, они могут размножаться апомиктически или вегетативно; восстановление плодовитости возможно в результате удвоения числа хромосом (полиплоидии).
Приложение 1. Вегетативное размножение: некоторые способы и перспективы
Способы вегетативного размножения растений многочисленны и разнообразны. Некоторые растения размножаются с помощью усов, или столонов, — длинных тонких побегов, которые стелятся по земле. У культурной земляники (Fragaria ananassa), например, листья, цветки и корни образуются на каждом узле такого побега. Сразу за вторым узлом верхушка его поднимается вверх и утолщается. На этой утолщенной части образуются первые придаточные корни, а затем и новый побег, от которого отходит новый ус.
Органами размножения во многих случаях могут служить корневища, особенно у злаков и осок. Корневища «захватывают» пространство около родительского растения, и каждый узел может дать новый цветоносный побег. Многие злостные сорняки размножаются таким способом, и поэтому от них так трудно избавиться, а некоторые садовые растения, например, ирисы, размножаются почти исключительно корневищами. Клубни и луковицы приспособлены для размножения и запасания веществ. Картофель искусственно размножают кусочками клубней с одним или несколькими «глазками». Именно глазок семенного картофеля дает начало новому растению.
Корни некоторых растений, например, вишни, яблони, малины и ежевики, формируют «корневые отпрыски», или корневую поросль, которые дают новые растения. Коммерческие сорта бананов не образуют семян и размножаются с помощью корневых отпрысков, образующихся из почек на подземных стеблях. Если разорвать корень одуванчика, что нередко случается при выпалывании, каждый кусочек корня может дать новое растение (см. конец гл. 22).
Некоторые виды растений способны размножаться с помощью листьев. Пример тому — комнатное растение Kalanchoe daigremontiana, называемое в некоторых странах «мать тысячи детей». Название этого растения основано на том, что из меристематической ткани в выемках краев листьев образуются многочисленные маленькие растеньица. Вид обычно и размножают с помощью этих «детей», которые, достигнув определенного возраста, падают на землю и укореняются. Другим примером может служить костенец корнелистный (Asplénium rhizophyllun), называемый также «странствующим папоротником», у которого молодые растения образуются в тех местах, где листья касаются земли.
У ряда растений, в том числе лимона, некоторых злаков (например, мятлика лугового, Гоа pratensis) и одуванчика, зародыши в семенах образуются бесполым путем. Данный способ вегетативного размножения называют апомиксисом. Из семян апомиктических растений развиваются особи, генетически идентичные материнским растениям1).
1 В настоящее время большинство исследователей не считает апомиксис вариантом вегетативного размножения, поскольку в нем участвуют те или иные половые структуры и элементы полового процесса, благодаря чему сохраняется некоторая возможность генетического разнообразия. — Прим. перев.
Гербарный образец костенца корнелистного (Asplénium rhizophyllum). Видно, как укореняются листья и образуются новые растения. Таким способом папоротник может образовать большие колонии генетически однородных растений

Kalanchoe daigremontiana с мокренькими растеньицами, выросшими в выемках по краям листа

Земляника (Fragaria ananassa) размножается бесполым путем с помощью усов. Ош может размножаться и половым путем, образуя цветки

В целом в результате бесполого размножения потомство в точности повторяет родительский организм, хорошо приспособленный к определенным условиям среды или местообитаниям. В основе адаптации могут лежать признаки, которые желательно придать культурным растениям или которые способствуют выживанию при определенном сочетании факторов внешней среды.
Приложение 2. Адаптивная радиация гавайских Мfdinae
На Гавайских островах встречаются группы интереснейших местных растений. Главные острова этого архипелага поднялись из моря независимо друг от друга миллионы лет назад. Местообитания растений и животных, заселивших острова, очень разнообразны. От ареалов на материке, из которых они некогда мигрировали, их отделяет большое расстояние. Организмы, достигшие Гавайских островов, зачастую сильно менялись, когда захватывали новые, подходящие для них местообитания. Процессы, с помощью которых происходят такие изменения, называются адаптивной радиацией.
Яркий пример адаптивной радиации — группа из 28 гавайских видов растений, принадлежащих к трем близким родам семейства Аsteraceae, а именно — к подтрибе Madiinae. Большинство ее представителей найдено в Калифорнии и прилегающих областях. Род Argyroxiphium очень близок к двум другим родам, также найденным только на Гавайских островах, — Dubautia и Wilkesia. Морфологическое разнообразие представителей их видов велико; от маленьких, образующих заросли кустиков и розеточных растений до крупных деревьев и лиан. Они растут в самых разнообразных местах: на застывшей лаве, в сухих кустарниках и редколесьях, в сырых лесах и на болотах. В этих местах количество выпадающих за год осадков колеблется от менее 40 см до более 1230 см. Некоторые из них входят в число самых влажных уголков суши.
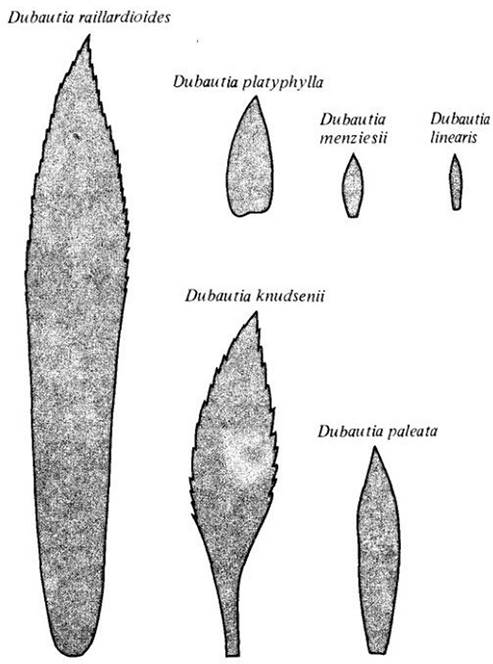
Столь разнообразные условия сопровождаются и значительной изменчивостью формы и размеров листьев. Например, виды Dubautia, растущие на хорошо освещенных сухих местах, обычно имеют очень мелкие листья, тогда как в затененном подлеске влажного леса — гораздо крупнее. Представитель другого рода Argyroxiphium sandwicense, растущий на сухих альпийских склонах кратера Халеакала на острове Мауи, имеет листья, покрытые множеством серебристых волосков. Эти волоски, по-видимому, защищают лист от интенсивной солнечной радиации и способствуют сохранению влаги. На листьях близкородственного растения А. grayanum, тоже встречающегося на Мауи, но в сырых лесах и на болотах, опушения нет, поэтому в засуху способны поддерживать более высокое тургорное давление (а соответственно и более высокие уровни метаболической активности).
Контуры листьев шести видов Dubautia из сухих и влажных мест, которые различаются по форме и величине
Argyroxiphium sandwicense — замечательное растение, растущее на открытых, покрытых пеплом склонах кратера Халеакала на острове Мауи в условиях интенсивной солнечной радиации и низкой влажности

Dubautia reticulata; виды рода Dubautia, охватывающего 21 из 28 встречающихся на Гавайях видов подтрибы Madiinae представлены, с одной стороны, деревьями и кустарниками, с другой — лианами и небольшими, едва одревесневшими растениями, образующей заросли. Dubautia reticulata растет во влажных лесах Мауи, может достигать восьми и более метров в высоту; диаметр ствола — около 0, 5 м

Wilkesia gymnoxiphium. Это причудливое растение, похожее на юкку, встречается только на острове Кауаи, где его распространение ограничено сухими заросшими кустарников по краю каньона Ваймеа. Кауаи — самый древний из главных острогов Гавайского архипелага. Роберт Робишо, сфотографированный рядом с этим растением, изучает физиологическую экологию этой интересной группы растений

Dubautia scabra. Низкие, образующие ковры травянистые представители этого вида встречаются на островах Гавайского архипелага в местообитаниях с высокой влажностью. Считается, что несколько других видов на более молодых островах архипелага произошли от этого вида

Гавайским Маdiinae, растущим в самых разнообразных условиях, свойственны и значительные физиологические различия. Роберт Робишо из Калифорнийского университета в Беркли показал, например, что виды Dubautia, встречающиеся на сухих местах, гораздо легче переносят недостаток воды, чем представители того же рода из сырых и переувлажненных мест. Дело оказалось в том, что в листьях устойчивых видов клетки имеют более эластичные оболочки и Гавайские острова. На самом древнем из главных островов, Кауаи, возраст некоторых пород достигает 6 млн. лет; самый молодой остров еще формируется. Гавайский архипелаг постепенно перемещается к северо-западу вместе с тихоокеанской тектонической плитой, причем самые старые из островов постепенно разрушаются в глубине моря, а самые молодые продолжают формироваться, очевидно, потому, что проходят над «горячей точкой» — участком земной коры, через который извергается лава. Это значит, что приблизительно там, где теперь находятся Гавайи, уже более 6 млн. лет назад находились острова
Несмотря на морфологическую неоднородность и разнообразие местообитаний, все 28 видов, относящиеся к трем родам, близко родственны друг другу. Два любых рода, насколько мы знаем, способны к гибридизации, и все гибриды, по крайней мере частично, плодовиты. Исследования этих взаимоотношений проводятся Джеральдом Карром из Гавайского университета и Дональдом Кайхосом из Калифорнийского университета в Дейвисе. Вся эта группа из трех родов эволюционировала в полной изоляции на Гавайских островах и произошла от единственного предкового растения, занесенного сюда с западного побережья Северной Америки.