СОВРЕМЕННАЯ БОТАНИКА - П. РЕЙВН - 1990
РАЗДЕЛ V. СТРОЕНИЕ И РАЗВИТИЕ ТЕЛА ПОКРЫТОСЕМЕННЫХ
ГЛАВА 21. КОРЕНЬ: ПЕРВИЧНОЕ СТРОЕНИЕ И РАЗВИТИЕ
У большинства сосудистых растений корни составляют подземную часть спорофита и служат прежде всего для закрепления его в почве и поглощения веществ (рис. 21-1). Известны и другие их функции, среди которых важнейшие — запасание и проведение. Большинство корней накапливает много запасов, а некоторые, например, у моркови, сахарной свеклы и батата, специально приспособлены для этого. Питательные вещества, синтезированные в надземных фотосинтезирующих частях растений, передвигаются по флоэме в запасающие ткани корня. Они могут быть частично использованы им самим, но чаще расщепляются и возвращаются по флоэме в надземные органы. У двулетников (растений, проходящих свой жизненный цикл в течение двух лет), в частности сахарной свеклы и моркови, в первый год запасающие ткани корня накапливают много питательных веществ, расходующихся на второй год при формировании цветков, плодов и семян. Вода и минеральные соли (т. е. неорганические ионы), поглощенные корнями, переносятся через ксилему к надземным органам. Туда же по ксилеме движутся гормоны (особенно цитокинины и гиббереллины), синтезированные в меристематических зонах корней и необходимые для роста и развития надземных частей растений (см. гл. 24).
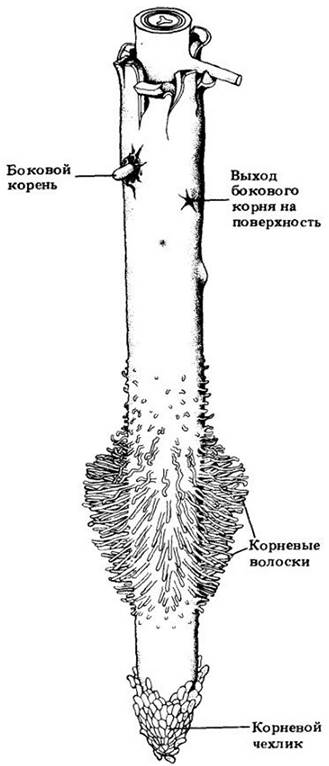
Рис. 21-1. Часть корня двудольного, где видны пространственные отношения между корневым чехликом, зоной корневых волосков и местами выхода на поверхность боковых корней, закладывающихся в глубине родительского корня. Новые корневые волоски образуются сразу же за зоной растяжения примерно с той же скоростью, с какой отмирают старые
Корневые системы
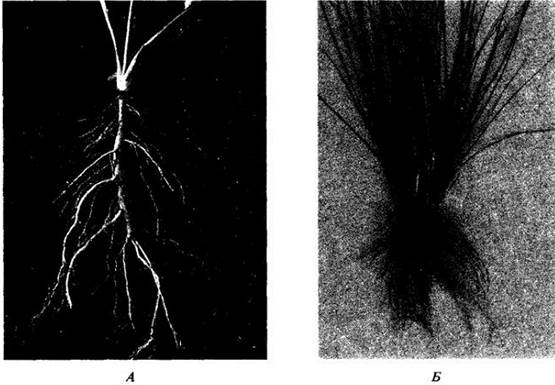
Первый корень растения, закладывающийся у зародыша, называется обычно первичным. У голосеменных и двудольных он становится главным и растет прямо вниз, давая боковые корни, самые старые из которых находятся около корневой шейки (на стыке корня и стебля), а самые молодые — ближе всего к кончику корня. Такая корневая система, включающая главный и боковые корни, называется стержневой (рис. 21-2, А).
У однодольных, первичный корень, как правило, недолговечен и корневая система развивается из придаточных корней, отходящих от стебля. Вместе с возникающими на них боковыми корнями они составляют мочковатую корневую систему, все элементы которой развиты более или менее одинаково (рис. 21-2, Б). Стержневая система проникает в почву обычно глубже, чем мочковатая, однако поверхностное заложение последней и цепкость, с которой она оплетает прилегающие частицы грунта, делает растения с такими корнями особенно ценными для создания дернового покрова, предупреждающего эрозию почв.
Рис. 21-2. Два типа корневых систем: А — стержневая у одуванчика (Taraxacum officinale): Б — мочковатая у злака
Насколько глубоко и насколько далеко в стороны распространяются корневые системы, зависит от нескольких факторов, включая влажность, температуру и состав почвы. Основная масса так называемых «питающих корней», активно участвующих в поглощении воды и минеральных солей, располагается в верхнем ее слое мощностью 1 м, причем у большинства деревьев — в верхних 15 см, т. е. в горизонте, обычно наиболее богатом органикой. Некоторые породы, например, ели, буки и осины, редко образуют глубокие стержневые корни, в то время как у других, в частности у дубов и многих сосен, они обычно очень развиты, вследствие чего эти виды плохо переносят пересадку. Рекордная глубина проникновения корней в почву отмечена у пустынного мескитового кустарника (Prosopis juliflora). Их находили на глубине 53,3 м при рытье котлована в 30 км к юго- востоку от Тусона (шт. Аризона) в 1960 г. Корни тамариска и акации обнаруживались в Египте во время сооружения Суэцкого канала на глубине 30 м. Корни деревьев распространяются в стороны обычно за пределы радиуса крон. Корневая система кукурузы часто заходит на глубину около 1,5 и примерно на 1 м во все стороны от растения. Корни люцерны (Medicago sativa)могут заглубляться на 6 м и более.
Живое растение нуждается в поддержании баланса между общими площадями фотосинтезирующей и поглощающей воду и минеральные вещества поверхностей. У молодого, только что прижившегося растения вторая обычно намного превышает первую, однако это отношение имеет тенденцию изменяться с возрастом на противоположное, что должны учитывать садоводы. Даже при очень осторожной пересадке баланс между побегом и корнем неизменно нарушается. Большинство тонких питающих корней при извлечении растения из почвы обрывается. Восстановить равновесие помогает обрезка побегов.
Одно из наиболее детальных исследований поверхности побега и корня было проведено на четырехмесячном растении ржи (Sесаlе сеrеаlе). У корневой системы, включая корневые волоски, она составила 639 м2, т. е. в 130 раз больше, чем у побега. Особенно поразительно то, что корни были распределены в объеме почвы, равном всего около 6 л.
Происхождение и рост первичных тканей
Рост многих корней представляет собой, по-видимому, непрерывный процесс, который прекращается только при таких неблагоприятных условиях, как засуха и низкая температура. В почве корни распространяются по пути наименьшего сопротивления и часто заполняют пространства, оставшиеся после их отмерших и сгнивших предшественников.
Кончик корня покрыт в виде наперстка корневым чехликом (рис. 21-1, 21-3 и 21-4) — массой клеток, защищающих апикальную меристему и способствующих продвижению корня в почве. По мере роста корня и проталкивания корневого чехлика вперед клетки на периферии последнего слущиваются, давая слизь, которая обволакивает корень, облегчая его скольжение между частицами почвы. Параллельно этому слущиванию апикальная меристема образует новые клетки корневого чехлика. Продолжительность их жизни (от возникновения до слущивания) составляет четыре—девять дней в зависимости от длины чехлика и вида растения.
Упомянутое слизистое вещество представляет собой сильно гидратированный полисахарид, вероятно пектиновой природы, секретируемый наружными клетками чехлика. Он накапливается в пузырьках диктиосом, которые, сливаясь с плазматической мембраной, высвобождают его в пространство между плазмалеммой и клеточной оболочкой. В конечном счете слизь выходит на поверхность последней, где образует мелкие капли.
Рис. 21-3. Продольные срезы конники корня лука (Allium сера). А. Непосредственно за апикальной можно различить первичные меристемы. Б. Фрагмент апикальной меристемы. Сравните ее строение с наблюдаемым в корне кукурузы (рис. 21-4)
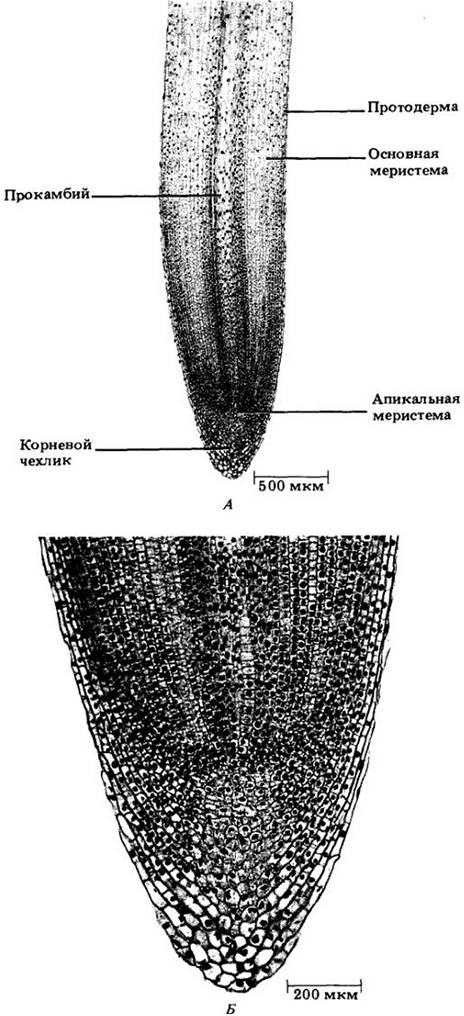
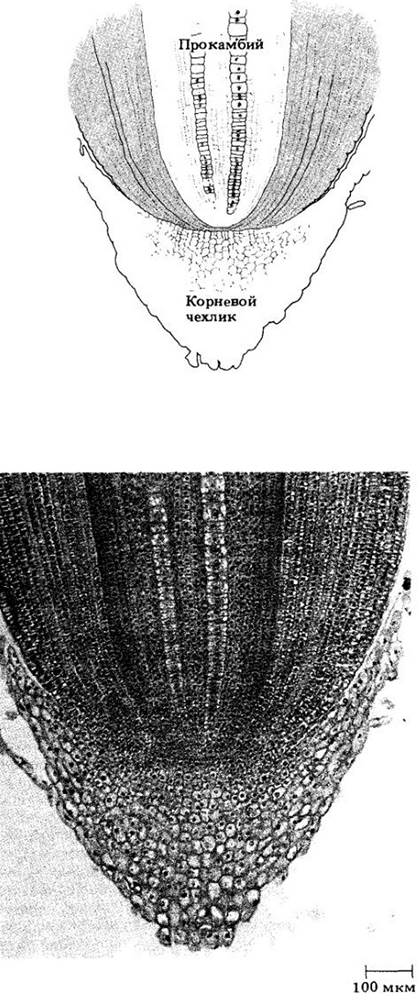
Рис. 21-4. Апикальная меристема кончика корня кукурузы (Zea mays). Обратите внимание на три отдельных слоя инициалей. Нижний дает начало корневому чехлику, средний — протодерме и основной меристеме (первичной коре), а верхний — прокамбию, т. е. центральному цилиндру
Корневой чехлик выполняет и другие важные функции, контролируя, в частности, реакцию корня на гравитацию (геотропизм, или гравитропизм, см. гл. 25).
Зоны роста корня
Наряду с корневым чехликом наиболее характерной структурной особенностью кончика корня является расположение продольных рядов клеток, берущих начало в апикальной меристеме. Она состоит из относительно мелких (от 10 до 20 мкм в диаметре) многогранных клеток — инициал ей и их непосредственных производных (с. 16), отличающихся густой цитоплазмой и крупными ядрами (рис. 21-3 и 21-4). Размещение и число инициалей заметно варьируют.
В корнях семенных растений описано два главных типа апикальной организации. В одном из них корневой чехлик, проводящий цилиндр (ксилема и флоэма) и кора возникают из общей группы клеток апикальной меристемы (рис. 21-3); в другом каждая эта область может быть выведена из независимого клеточного слоя (рис. 21-4), причем эпидерма имеет общее происхождение либо с корневым чехликом, либо с корой.
Хотя зона инициалей апикальной меристемы корня считалась когда-то местом активных клеточных делений, сейчас показано, что это не так и большинство делений происходит на некотором расстоянии от инициалей, образующих относительно неактивную область, называемую покоящимся центром (рис. 21-5).
Рис. 21-5. Апикальная меристема кончика корня кукурузы с покоящимся центром (обведен штриховой линией). Чтобы получить этот радиоавтограф, корень в течение суток заставляли поглощать радиоактивный тимин, меченный тритием (3Н). В активно делящихся клетках вокруг покоящегося центра это основание быстро встраивалось в ядерную ДНК, что можно проследить по участкам с темной зернистостью на радиоавтографе
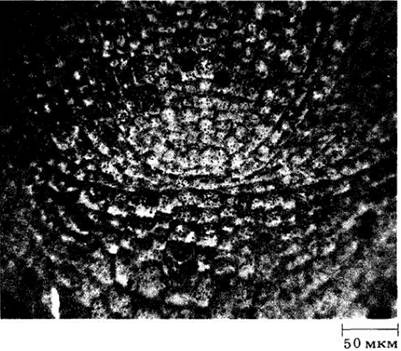
Слово «относительно» указывает, что в обычных условиях деления в нем все-таки происходят. Более того, этот центр способен восстанавливать периферические зоны меристемы при их повреждении. Недавно показано, например, что изолированные покоящиеся центры кукурузы, выращенные в стерильной культуре, способны вырастать в целые корни, минуя стадию каллуса, или раневой ткани. В другой работе поэтому же виду обнаружена четкая корреляция между размерами покоящегося центра и сложностью строения первичной проводящей системы корня. Эти и другие исследования говорят о существенной роли покоящегося центра в формировании структуры и развитии корня.
Расстояние от апикальной меристемы, на котором происходит большинство клеточных делений, неодинаково у разных видов, а в пределах вида — у корней разного возраста. Апикальная меристема вместе с этой прилегающей к ней частью корня называется зоной клеточных делений (рис. 21-6).
Рис. 21-6. Схема ранних стадий первичного роста кончика корня (ср. с рис. 21-1)
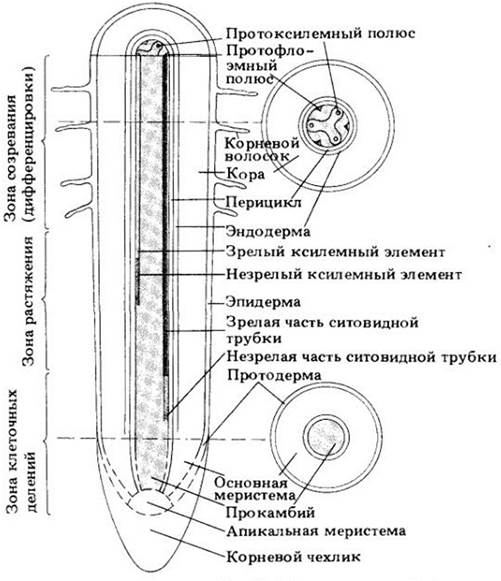
За ней без резкой границы следует зона растяжения длиной обычно всего несколько миллиметров (рис. 21-6). Именно рост клеток в этой зоне обусловливает основное удлинение органа. Дальше от апекса оно уже прекращается. Поскольку рост в длину происходит только около кончика корня, лишь очень ограниченная его часть постоянно продвигается в почве.
За зоной растяжения находится зона созревания (дифференцировки) большинства клеток первичных тканей (рис. 21-6). Здесь развиваются также корневые волоски, вследствие чего эту часть корня называют иногда зоной корневых волосков (см. рис. 21-1).
Важно отметить, что переход от одной зоны к другой происходит постепенно, без резких границ. Некоторые клетки начинают удлиняться и дифференцироваться еще в зоне клеточных делений, в то время как другие достигают зрелости в зоне растяжения, например, первые элементы флоэмы и ксилемы, которые из-за продолжающегося растяжения часто разрушаются в процессе удлинения корня.
Вблизи апикальной меристемы уже можно различить протодерму, прокамбий и основную меристему (см. рис. 21-3 и 22-6), т. е. первичные меристемы, дифференцирующиеся соответственно в эпидерму, первичные проводящие ткани и первичную кору (гл. 19).
Первичная структура
По сравнению с внутренней структурой стебля у корня она относительно проста. Это связано прежде всего с отсутствием здесь листьев и соответственно узлов и междоузлий, за счет чего в расположении тканей на разных уровнях наблюдаются очень небольшие различия.
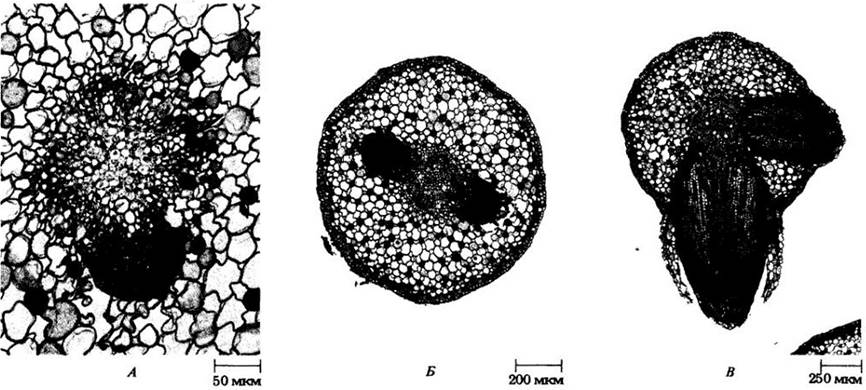
На первичной стадии роста корня поперечные (рис. 21-7 и 21-8) и продольные (см. рис. 21-3) срезы легко позволяют выделить три системы тканей: эпидерму (систему покровной ткани), первичную кору (систему основной ткани) и систему проводящих тканей. У большинства корней последняя образует сплошной цилиндр (рис. 21-7), а у некоторых — полый цилиндр вокруг сердцевины (рис. 21-8).
Рис. 21-7. Поперечные срезы корня лютика (Ranunculus). А. Общий вид для зрелого корня. Б. Деталь незрелого центрального цилиндра. Обратите внимание на межклетники о первичной коре. В. Деталь зрелого центрального цилиндра. В коровых клетках заметны многочисленные крахмальные зерна
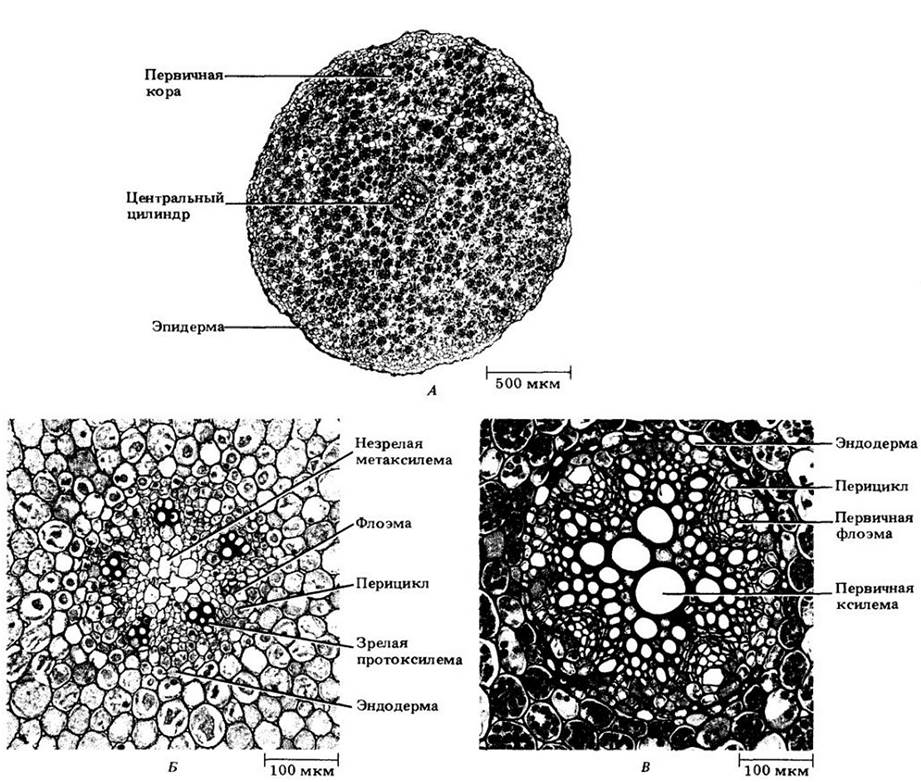
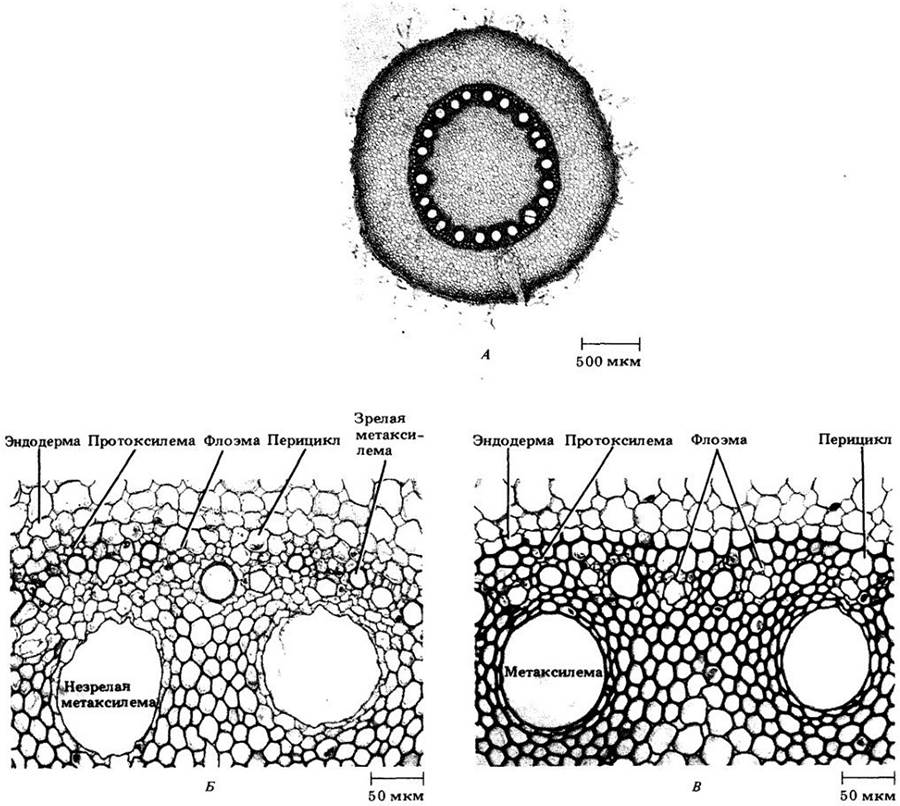
Рис. 21-8. Поперечные срезы корня кукурузы. А. Общий вид для зрелого корня. Внизу справа можно видеть часть бокового корня. Четко выделяется центральный цилиндр с сердцевиной. Б. Деталь незрелого центрального цилиндра. В. Деталь зрелого центрального цилиндра
Эпидерма
Эпидерма молодых корней абсорбирует воду и минеральные вещества; выполнению этой функции способствуют корневые волоски — трубчатые выросты эпидермальных клеток, существенно увеличивающие поглощающую поверхность органа (рис. 21-9, см. также раздел «Микориза» в гл. 13). В ранее упоминавшейся работе с четырехмесячными растениями ржи установлено, что у одной особи примерно 14 млрд корневых волосков с площадью поглощения 401 м2 и суммарной длиной более 10 000 км.
Рис. 21-9. Корень проростка полевицы тонкой (Agrostis tenuis) с корневыми волосками. Они могут достигать окончательной длины 1,3 см за несколько часов. Каждый волосок живет сравнительно недолго, но образование новых и отмирание старых продолжаются все время, пока корень растет
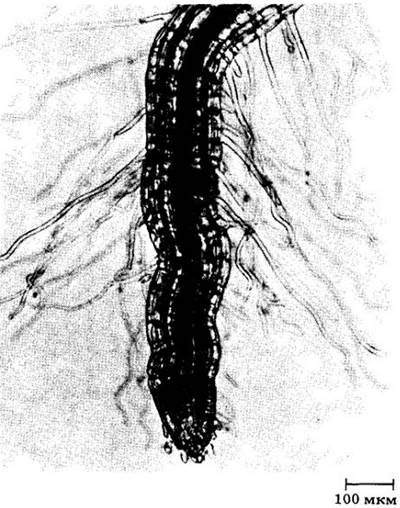
Эти волоски относительно недолговечны, приурочены главным образом к зоне созревания и образуются сразу же за зоной растяжения (см. рис. 21-1 и 21-6) примерно с такой же скоростью, с какой отмирают старые на верхнем конце покрытой ими зоны. По мере проникновения кончика корня в почву новые корневые волоски развиваются непосредственно за ним, давая поверхность, способную поглощать новые достигнутые запасы воды и минеральных ионов (см. гл. 27). Очевидно, именно молодые и растущие корни являются питающими, т. е. прежде всего участвуют в поглощении воды и неорганических веществ. В связи с этим при пересадке садоводы должны уделять особое внимание перенесению на новое место корневой системы с как можно большим количеством почвы. Если растение просто вырвать, основная масса питающих корней останется в земле и этот «саженец» скорее всего уже не приживется.
Эпидермальные клетки корня, включая те, что несут корневые волоски, являются паренхимными, плотно упакованными. Молодая эпидерма большей частью несет тонкую кутикулу, вследствие чего клеточная оболочка оказывает некоторое сопротивление прохождению сквозь нее воды и минеральных веществ. Кроме того, поверхность многих корней покрыта слизистым чехлом, способствующим установлению более тесного контакта с частицами почвы. Происхождение этого чехла еще точно не установлено, но предполагается, что он, по крайней мере частично, образуется корневым чехликом. Показано, что слизь создает благоприятные условия для поселения полезных бактерий. Она может также влиять на доступность почвенных ионов и обеспечивать кратковременную защиту корня от иссушения.
Первичная кора
Как показывает поперечный срез (см. рис. 21-7, А), на первичную кору приходится основная масса первичных тканей корня. Ее клетки накапливают крахмал и другие вещества, но обычно не содержат хлоропласгов. Корни с интенсивным вторичным ростом, например, у голосеменных и большинства двудольных, сбрасывают первичную кору рано. У них ее клетки остаются паренхиматозными. В противоположность этому первичная кора однодольных сохраняется в течение всей жизни корня, и многие ее клетки, лигнифицируясь, образуют вторичные оболочки.
Независимо от степени дифференцировки эта ткань содержит многочисленные межклетники — воздушные полости, имеющие большое значение для аэрации клеток корня (см. рис. 21-7 и 21-8). Клетки здесь образуют друг с другом многочисленные контакты, и их протопласты связаны плазмодесмами. В результате вещества в коре могут переходить из клетки в клетку по протопластам и плазмодесмам или по клеточным оболочкам.
В отличие от остальной части первичной коры в самом внутреннем ее слое клетки расположены плотно, без межклетников. Эта область, эндодерма (см. рис. 21-7 и 21-8), характеризуется присутствием на антиклинальных (т. е. перпендикулярных поверхности корня) клеточных стенках поясков Каспари. Так называют экваториальную часть первичной оболочки, пропитанную жироподобным веществом суберином, а иногда и лигнифицированную. Протопласт эндодермальных клеток плотно к ней прилегает (рис. 21-10 и 21-11). Поскольку в эндодерме клетки располагаются плотно, а пояски Каспари не проницаемы для воды, все вещества, поступая в центральный проводящий цилиндр и покидая его, должны проходить через протопласты эндодермальных клеток, либо пересекая их плазматическую мембрану, либо передаваясь через многочисленные плазмодесмы, образующие связь с протопластами соседних клеток первичной коры и центрального цилиндра.
Рис. 21-10. Часть среза незрелого корня лютика (Ranunculus) при большом увеличении с поясками Каспари в клетках эндодермы. Заметно, что плазмолизированные протопласты этих клеток прилегают к пояскам
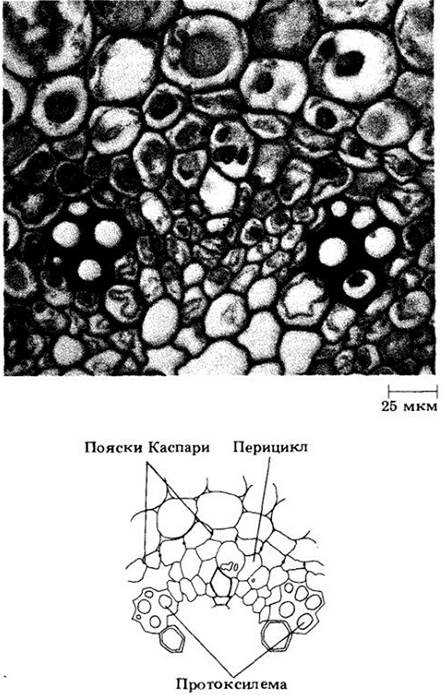
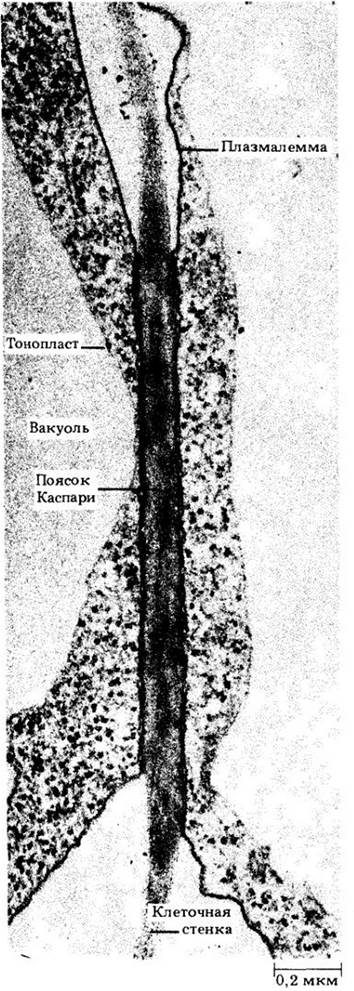
Рис. 21-11. Поперечный срез радиальной стенки между двумя эндодермалъными метками. Участок пояска Каспари окрашен интенсивнее остальных частей стенки. Видно, как плотно прилегает к стенке плазмалема каждой из этих плазмолизированных клеток в зоне пояска Каспари
Эффективность пояска Каспари как барьера для передвижения веществ через клеточные оболочки эндодермы показана на корнях кукурузы, поглощавших положительно заряженный ион лантана, не способный проникать через клеточные мембраны. Под электронным микроскопом этот элемент обнаружен только в клеточных оболочках коры, т. е. его транспорт в глубь корня резко и полностью остановлен поясками Каспари (см. гл. 27, в которой более подробно рассматривается роль эндодермы в передвижении воды и растворенных веществ по корню).
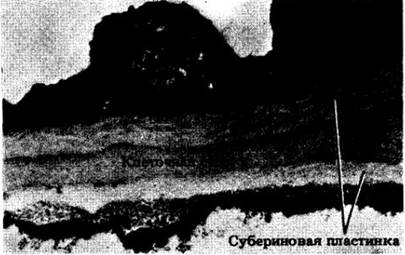
Как упоминалось выше, у корней с вторичным ростом первичная кора с эндодермой рано сбрасывается. В тех случаях, когда она сохраняется, по всей внутренней поверхности клеточных оболочек эндодермы со временем откладывается субериновая пластинка из чередующихся слоев суберина и воска, а после этого — целлюлоза, которая может лигнифицироваться (рис. 21-12). Такие изменения в эндодерме начинаются против флоэмных тяжей и распространяются к протоксилеме (см. рис. 21-7, А), причем против последней некоторые эндодермальные клетки могут оставаться тонкостенными, долгое время сохраняя свои пояски Каспари. Их называют пропускными; у большинства видов они в конце концов суберинизируются. У хвойных изменение клеточных оболочек в эндодерме оканчивается отложением субериновой пластинки (рис. 21-13).
Рис. 21-12. Трехмерная схема, показывающая три стадии развития эндодермальной клетки в первичном теле корня. А. Сначала эндодермальная клетка характеризуется присутствием на антиклинальных стенках пояска Каспари. Б. Затем изнутри по всей поверхности ее оболочки откладывается субериновая пластинка. В. Наконец, субериновая пластинка дополняется толстым, часто лигнифицированным слоем целлюлозы
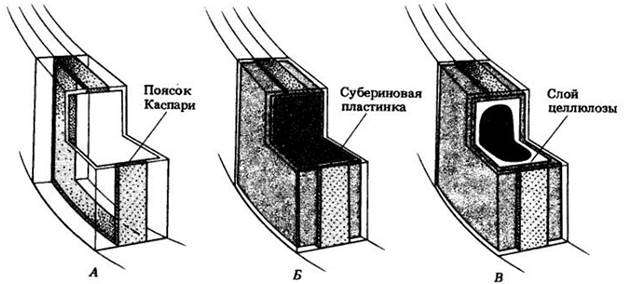
Рис. 21-13. Электронная микрофотография субериновых пластинок в стенках двух соседних эндодермальных клеток корня сосны смолистой (Pinus resinosa). Заметны чередующиеся светлые и темные слои, образованные, как считается, воском и суберином соответственно
Центральный цилиндр
Центральный цилиндр корня состоит из проводящей ткани, окруженной одним или несколькими слоями клеток, перициклом (см. рис. 21-7 и 21-8). В молодом корне перицикл состоит из паренхимных клеток с первичными оболочками, которые по мере старения растения могут дополняться вторичными (см. рис. 21-8).
Перицикл выполняет несколько важных функций. У большинства семенных растений в нем закладываются боковые корни. У видов с вторичным ростом он участвует в формировании камбия и обычно дает начало первому слою феллогена. В перицикле часто происходит образование новых клеток, входящих затем в его состав.
Внутреннюю часть центрального цилиндра у большинства корней занимает сплошной тяж первичной ксилемы, дающий к перициклу выступы в виде ребер (см. рис. 21-7). Между ними размещаются тяжи первичной флоэмы. Таким образом, центральный цилиндр корня является протостелой.
Число выступов первичной ксилемы неодинаково у разных видов, а иногда изменяется даже вдоль оси одного корня. Если их два, корень называют диархным, если три — триархным, четыре — тетрархным (см. рис. 21-7, А, В), а если много — полиархным (см. рис. 21-8). Созревание (прото-)ксилемных элементов начинается под перициклом; вершины ребер обычно называют протоксилемными полюсами (см. рис. 21-7 и 21-8). Метаксилема (meta- по-гречески означает «после»), занимающая внутренние части ребер и середину центрального цилиндра, созревает после протоксилемы. Корни некоторых однодольных (кукурузы, например) имеют сердцевину (см. рис. 21-8), которая рассматривается некоторыми ботаниками как потенциальная сосудистая ткань.
Образование боковых корней
У большинства семенных растений боковые корни берут начало в перицикле. В связи с тем, что они закладываются глубоко в материнском корне, их называют эндогенными, что в переводе с греческого означает «зарождающиеся внутри» (рис. 21-1 и 21-14).
Рис. 21-14. Три стадии развития боковых корней ивы (Salix). А. Один зачаток уже сформировался (внизу); в перицикле закладываются два других (стрелки). Центральный цилиндр еще очень молодой. Б. Два корневых зачатка пробиваются через кору. В. Один боковой корень вышел на поверхность, другой приближается к ней
Деления в перицикле, приводящие к образованию боковых корней, происходят на некотором расстоянии от зоны растяжения в частично или полностью дифференцированных тканях корня. У покрытосеменных в появлении здесь корневого примордия участвуют клетки как перицикла, так и эндодермы, хотя во многих случаях производные последней недолговечны. По мере увеличения размеров молодого бокового корня он проходит через первичную кору (рис. 21-14, В), возможно секретируя при этом ферменты, разрушающие коровые клетки на своем пути. Уже на ранних стадиях развития этот зачаток формирует корневой чехлик, апикальную и первичные меристемы. Сначала центральные цилиндры бокового и родительского корней не связаны друг с другом. Они соединяются позднее за счет дифференцировки лежащих между ними паренхимных клеток в элементы ксилемы и флоэмы.
Воздушные корни
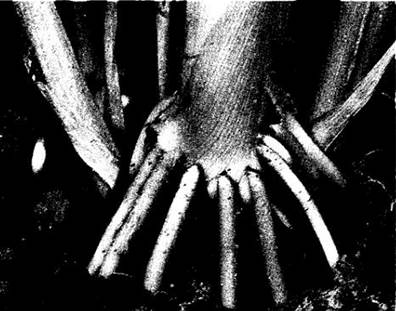
Воздушными называют придаточные корни, развивающиеся на надземных органах. У некоторых растений они служат опорой и называются в разных случаях столбовидными, ходульными или корнями-подпорками (рис. 21-15). Вступая в контакт с почвой, они разветвляются и также начинают поглощать воду и минеральные вещества. Такие корни развиваются на стволах и ветвях многих тропических деревьев, в частности у мангровых видов (Rhizophora mangle), баньяна (Ficus benghalensis) и некоторых пальм. Воздушные корни другого типа, например, у плюща (Hederá helix), внедряются в поверхность стен и других предметов, закрепляя на них лазающий стебель.
Рис. 21-15. Опорные корни кукурузы — один из типов придаточных корней
Корни нуждаются в кислороде для дыхания, и по этой причине большинство растений не может жить в недостаточно дренированной почве, где отсутствуют воздушные полости. Некоторые деревья болотистых местообитаний развивают корни, поднимающиеся из воды и служащие не только для заякоривания в субстрате, но и для снабжения кислородом. Например, корневая система авиценнии (Avicennia germinalis) образует отрицательно-геотропные выросты, называемые пневматофорами (дыхательными корнями), которые, торча из ила вверх, обеспечивают необходимую аэрацию (рис. 21-16). Сходную функцию приписывали «коленам» болотного кипариса (Taxodium distichum; см. рис. 18-27), но это представление теперь оспаривается.
Рис. 21-16. Пневматофоры (дыхательные корни) мангрового дерева Lagunculariа racemosa, поднимающиеся из ила у основания растения

Специальные приспособлении
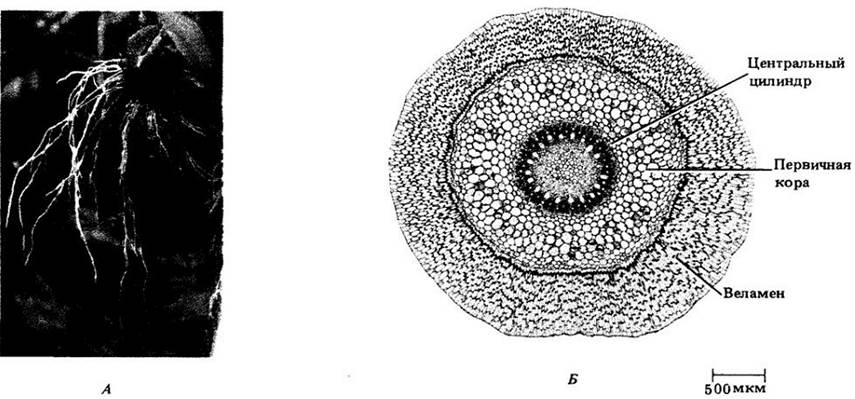
Многие специальные приспособления (адаптации) корней встречаются у эпифитов — растений, произрастающих на других растениях, не паразитируя на них. Например, многослойная эпидерма корня некоторых орхидных (рис. 21-17) иногда является единственным фотосинтезирующим органом растения. Такая ткань, называемая веламеном,обеспечивает также механическую защиту коры, сокращает потери воды и может участвовать в ее поглощении.
Рис. 21-17. А. Воздушные корни орхидного Oncidium sphacelatum. Б. Поперечный срез корня орхидного с многослойной эпидермой (веламеном)
Эпифит Dischidia rafflesiana, называемый иногда «цветочным горшком», имеет весьма своеобразное строение. Некоторые его листья уплощенные суккулентные, а другие образуют полые сосуды («цветочные горшки»), собирающие мелкие частицы и дождевую воду (рис. 21-18). Здесь поселяются колонии муравьев, что увеличивает азотное питание растений. Корни, образовавшиеся в узле над таким модифицированиым листом, проникают в «горшок», из которого поглощают воду и минеральные вещества.
Рис. 21-18. Эпифит Dischidia rafflesiana («цветочный горшок»). A. Видоизмененный в «горшок» лист, накапливающий мелкие твердые частицы и дождевую воду. Б. Тот же лист вскрыт, чтобы показать заходящие в него корни
Приспособления к накоплению питательных веществ
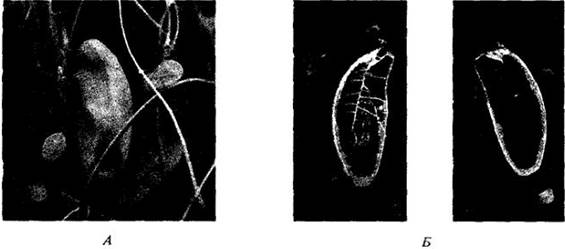
Многие корни являются запасающими органами, а у некоторых растений они специально приспособлены для выполнения этой функции, становясь мясистыми вследствие сильного разрастания паренхимы, пронизанной проводящей тканью. Развитие таких корней (например, у моркови, Daucus carota) происходит в основном так же, как и в случае немясистых, но с преобладанием во вторичных ксилеме и флоэме паренхимных клеток. Корень батата (Ipomoea batatas) примерно такой же, как у моркови, но у него вокруг отдельных сосудов или групп сосудов вторичной ксилемы закладываются добавочные камбиальные клетки (рис. 21-19). Отложив по нескольку трахеальных элементов в сторону сосудов и ситовидных трубок в противоположном направлении, они образуют с обеих сторон преимущественно клетки запасающей паренхимы. У сахарной свеклы (Beta vulgaris) утолщение корня происходит главным образом за счет деятельности дополнительных камбиев, закладывающихся концентрическими слоями вокруг первичного (рис. 21-20). Эти слои, внешне напоминающие годичные кольца корней и стеблей деревьев, откладывают внутрь и наружу от себя соответственно ксилему и флоэму с преобладающей в них паренхимой. Верхняя часть большинства мясистых корней в действительности развивается из гипокотиля.
Рис. 21-19. Поперечные срезы корня батата (Iроmоеа batatas). А. Общий вид. Б. Деталь ксилемы с камбием вокруг сосудов
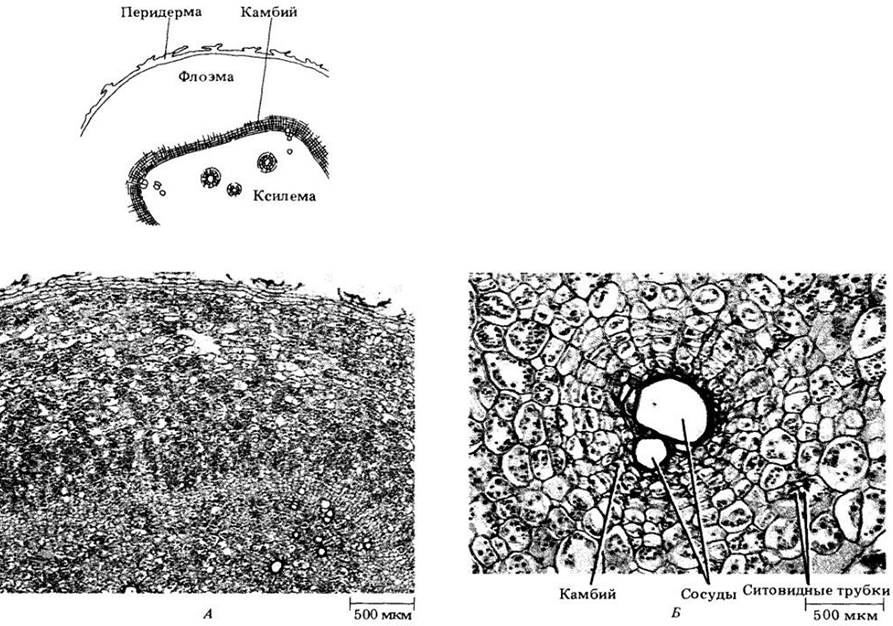
Рис. 21-20. Поперечный срез корня сахарной свеклы (Beta vulgaris) с дополнительными слоями камбия (обозначены стрелками). Исходный камбий (в центре корня) образует относительно мало ксилемы и флоэмы
Заключение
Корни представляют собой органы, специализированные для закрепления растения в субстрате, поглощения, накопления и проведения веществ. Голосеменные и двудольные обычно образуют стержневые, а однодольные — мочковатые корневые системы. Протяженность их зависит от разных факторов, но основная масса питающих корней находится в верхнем метре почвы.
Апикальные меристемы большинства корней содержат покоящийся центр; максимальная меристематическая активность, т. е. наиболее интенсивное деление клеток, наблюдается на некотором расстоянии от апикальных инициалей. В процессе первичного роста апикальная меристема дает начало трем первичным меристемам — протодерме, основной меристеме и прокамбию, дифференцирующимся соответственно в эпидерму, первичную кору и центральный цилиндр. Кроме того, апикальная меристема образует корневой чехлик, защищающий ее и способствующий внедрению корня в почву.
Многие эпидермальные клетки корня образуют корневые волоски, значительно увеличивающие его поглощающую поверхность. За исключением эндодермы, повсюду в коре имеются многочисленные межклетники. Плотно расположенные клетки эндодермы несут на своих антиклинальных стенках пояски Каспари. Вследствие этого все вещества, переносимые между первичной корой и центральным цилиндром, должны пройти через протопласты эндодермальных клеток.
Центральный цилиндр состоит из первичных проводящих тканей и полностью их окружающего перицикла. Среднюю часть цилиндра обычно занимает первичная ксилема с выростами, лучами, расходящимися к периферии и чередующимися с тяжами первичной флоэмы. Боковые корни закладываются в перицикле и пробиваются наружу сквозь первичную кору и эпидерму.