СОВРЕМЕННАЯ БОТАНИКА - П. РЕЙВН - 1990
РАЗДЕЛ V. СТРОЕНИЕ И РАЗВИТИЕ ТЕЛА ПОКРЫТОСЕМЕННЫХ
ГЛАВА 22. ПОБЕГ: ПЕРВИЧНАЯ СТРУКТУРА И РАЗВИТИЕ
Побег, состоящий из стебля и листьев, закладывается в процессе развития зародыша, в котором он представлен почечкой, состоящей из стебелька (эпикотиля), одного или нескольких листовых примордиев (зачатков) и апикальной меристемы (почечка может считаться первой почкой растения). С возобновлением роста зародыша при прорастании семени из апикальной меристемы развиваются новые листья, а стебель удлиняется и дифференцируется на узлы и междоузлия. Постепенно в пазухах листьев формируются примордии почек (рис. 22-1 и 22-2), со временем проходящие последовательные этапы роста и дифференцировки, более или менее сходные с наблюдавшимися у первой почки. Эта схема развития в ходе образования побеговой системы растения многократно повторяется.
Рис. 22-1. Часть побега кротона (Croton). Листья этого двудольного растения пятнистые и расположены на стебле спирально. У апекса побега они настолько сближены, что выделить узлы и междоузлия как отдельные участки невозможно. Удлинение стебля между листьями, прикрепленными к нему в узлах, приводит к образованию междоузлий
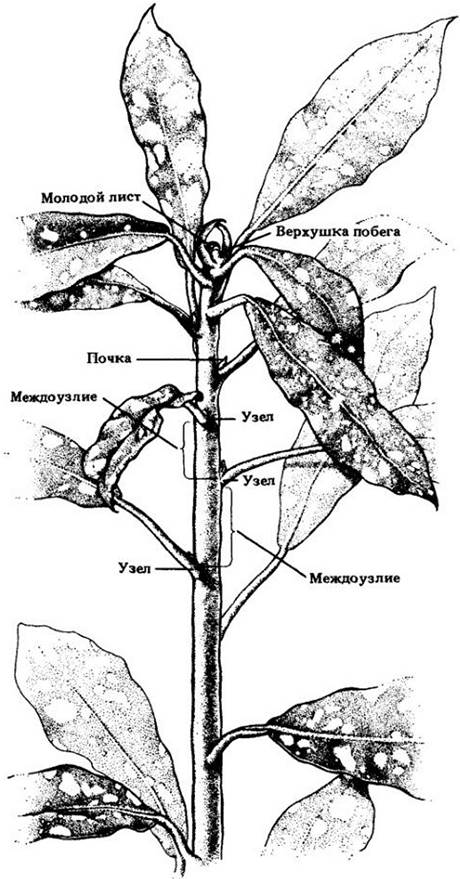
Рис. 22-2. Продольный срез верхушки побега распространенного комнатного растения Coleus blumei из двудольных. Листья здесь расположены в узлах супротивно, причем каждая последующая пара перпендикулярна предыдущей; таким образом, листья второго снизу узла находятся под прямым углом к плоскости среза
Растущая терминальная (верхушечная) почка побега обычно тормозит развитие латеральных (боковых). Это называется апикальным доминированием (см. гл. 24). Когда расстояние между верхушкой побега и латеральными почками возрастает, это тормозящее влияние уменьшается и последние могут начать свое развитие. На описанном явлении основана чеканка, т. е. удаление верхушек побега — обычный прием, которым пользуются в садоводстве для усиления кустистости растений.
Две главные функции стебля — проводящая и опорная. Вещества, синтезируемые в листьях, передвигаются по флоэме стебля к местам потребления, включая растущие листья, сами стебли, корни, развивающиеся цветки, семена и плоды. Много питательных соединений накапливается в паренхимных клетках корней, семян и плодов, но стебли также являются важными запасающими органами, а некоторые из них, например, подземные стебли картофеля, специально приспособлены для этого. Главные фотосинтезирующие органы растения — листья — поддерживаются стеблями, которые способствуют их благоприятному положению по отношению к свету. Кроме того, большая часть воды теряется растением через листья (гл. 27), поступая к ним по ксилеме от корней через стебель.
Происхождение и рост первичных тканей стебля
Строение апикальной меристемы побега сложнее, чем в корне. Здесь она не только образует клетки первичных тканей, но и участвует в формировании примордиев листьев и пазушных почек (рис. 22-2), развивающихся затем в боковые побеги. Эта меристема не защищена структурой, подобной корневому чехлику.
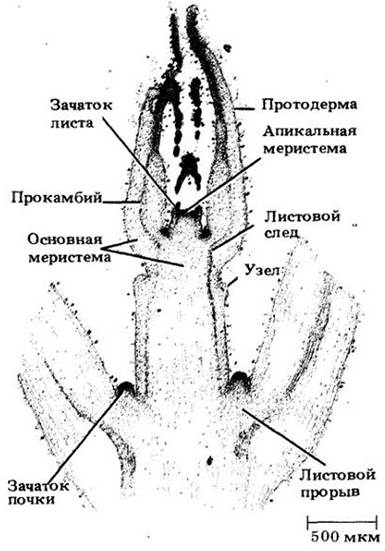
Вегетативный апекс побега большинства цветковых построен по так называемому типу туники-корпуса (рис. 22-3), двух зон, обычно различающихся плоскостями клеточных делений. Туника включает наружный слой (или слои) клеток, делящихся антиклинально и обеспечивающих главным образом увеличение поверхности. Корпус образован массой клеток, лежащих под туникой. Здесь они делятся в различных плоскостях, давая основной объем побега. Корпус и каждый слой туники имеют собственные инициали.
Рис. 22-3. Апекс побега Coleus blumei с организацией меристемы по типу туника — корпус. Зона центральных материнских клеток примерно соответствует корпусу
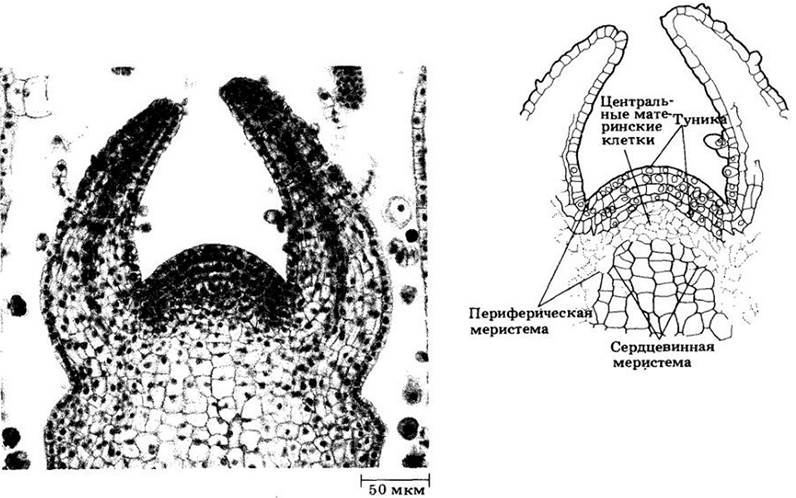
У многих покрытосеменных корпус соответствует области отчетливо вакуолизированных клеток, называемой зоной центральных материнских клеток (рис. 22-3). Она окружена периферической меристемой, происходящей частично из туники, частично из корпуса, а вглубь от нее располагается сердцевинная меристема. В зоне центральных материнских клеток клеточные деления относительно нечасты, зато периферическая зона митотически очень активна. Протодерма всегда происходит из наружного слоя туники, в то время как прокамбий и часть основной меристемы (кора и иногда часть сердцевины) из периферической меристемы. Остальная часть основной меристемы (вся или большая часть сердцевины) формируется сердцевинной меристемой.
Хотя у первичных тканей стебля периоды роста сходны с выделяемыми для корня, стебель не может быть разделен вдоль оси на зоны клеточного деления, растяжения и дифференцировки. В период активного роста апикальная меристема побега образует листовые примордии в такой быстрой последовательности, что узлы и междоузлия сначала не различимы. Постепенно участки между ярусами листьев начинают расти и, вытягиваясь, приобретают вид междоузлий; в то время как области прикрепления листьев оформляются в узлы (рис. 22-4). Таким образом, рост стебля в длину происходит в основном за счет удлинения междоузлий.
Рис. 22-4. Стадии развития верхушечной и двух боковых почек конского каштана (Aesculus hippocastanum). А. Молодые побеги плотно упакованы в почках и защищены почечными чешуями. Б. Почки вскрываются, обнажая самые старые рудиментарные листья. В. Удлинение междоузлий отделило друг от друга узлы. Верхушечная почка конского каштана смешанная, т. е. дает и листья, и цветки (последние на рисунке не видны). Боковые почки образуют только листья. Г. Нижние части молодых побегов; почечные чешуи разделены и отогнуты

Обычно меристематическая активность, приводящая к их удлинению, более интенсивна в основании этих развивающихся структур. Если удлинение междоузлия происходит относительно долго, такая зона у его основания может быть названа интеркалярной (т. е. находящейся между более дифференцированными областями) меристемой. Здесь дифференцируются некоторые элементы первичных флоэмы и ксилемы, в частности протофлоэма и протоксилема, которые связывают находящиеся выше и ниже более дифференцированные ткани.
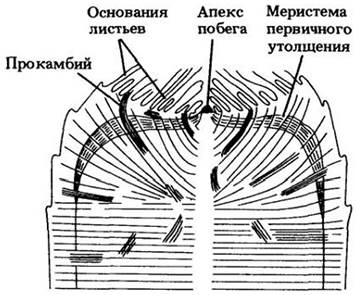
Первичный рост стебля в толщину осуществляется за счет как продольных делений, так и растяжения клеток первичной коры и сердцевины. У растений с вторичным ростом он умеренный. Однако у многих однодольных, например, пальм, происходит существенное первичное утолщение. Этот рост обычно идет вблизи апикальной меристемы (часто находящейся в углублении апекса) и приурочен к относительно узкой околопериферической зоне, называемой меристемой первичного утолщения (рис. 22-5).
Рис. 22-5. Схема анатомического строения верхушки, или розетки, толстостебельного однодольного без вторичного роста. Разрастание таких стеблей в толщину происходит благодаря активности меристемы первичного утолщения. Апикальная меристема и самые молодые листовые примордии как бы погружены в окружающие ткани стебля. У его верхушки ряды клеток расположены наподобие стопки блюдец
Как и в корне, апикальная меристема побега дает начало первичным меристемам — протодерме, основной меристеме и прокамбию (см. рис. 22-2). Они в свою очередь развиваются соответственно в эпидерму, основную и первичную проводящую ткани.
Первичное строение стебля
Это строение у семенных растений весьма разнообразно, но можно выделить три основных его типа. 1. У некоторых хвойных и двудольных узкие вытянутые прокамбиальные клетки и развивающиеся из них первичные проводящие ткани образуют более или менее сплошной полый цилиндр, погруженный в основную ткань (рис. 22-6). Ее наружная зона называется первичной корой, а внутренняя — сердцевиной. 2. У других хвойных и двудольных первичные проводящие ткани формируют цилиндр из отдельных тяжей, разделенных основной тканью (рис. 22-7). Эта ткань между прокамбиальными тяжами (а позже — зрелыми проводящими пучками), не прерываясь, переходит в первичную кору и сердцевину и называется межпучковой паренхимой. Ее межпучковые участки часто называют сердцевинными лучами. Такие узкие участки связывают кору с сердцевиной и при первом типе строения, но там они слабо выражены. 3. В стеблях большинства однодольных и некоторых травянистых двудольных расположение проводящих пучков более сложное. Они не располагаются одним кольцом между первичной корой и сердцевиной, а обычно образуют несколько колец или систему тяжей, рассеянных среди основной ткани, которая в этом случае часто не может быть разделена на первичную кору и сердцевину (рис. 22-8).
Стебель липы
Стебель липы (Tilia americana) построен по первому рассмотренному типу (рис. 22-6). Как и в большинстве случаев, его эпидерма представлена единственным слоем клеток, покрытых кутикулой. В ней здесь гораздо меньше устьиц, чем у листьев.
Рис. 22-6. А. Поперечный срез стебля липы (Tilia americana) на стадии первичного роста. Проводящие ткани образуют непрерывный полый цилиндр, делящий основную ткань на внутреннюю сердцевину и наружную первичную кору. Б. Фрагмент того же самого стебля при большем увеличении
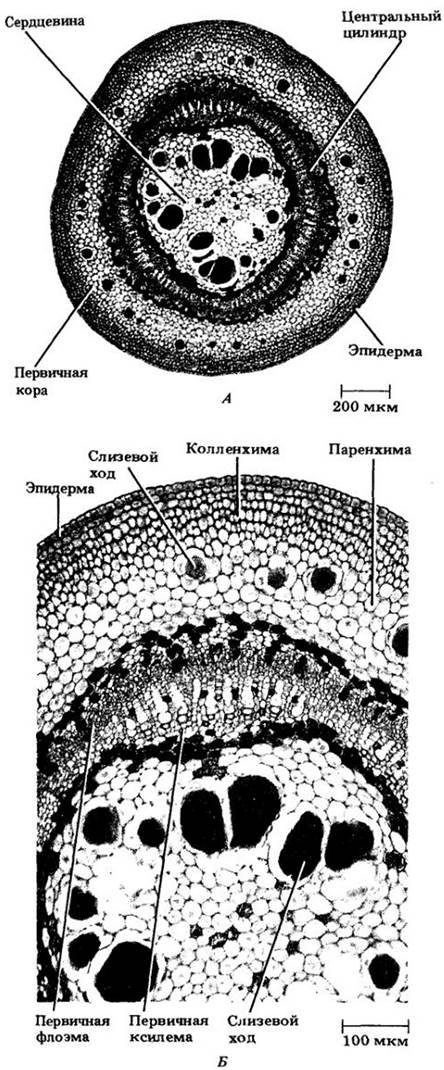
Первичная кора включает паренхимные и колленхимные клетки. Несколько слоев последних, обеспечивающих опору молодого стебля, образуют под эпидермой непрерывный цилиндр. Остальная часть первичной коры состоит из паренхимных клеток, содержащих в зрелом состоянии хлоропласты. Внутренний слой коры благодаря своей темной окраске резко отграничивает се от первичных проводящих тканей.
У подавляющего большинства стеблей, в том числе и у липы, первичная флоэма развивается из наружных клеток прокамбия, а первичная ксилема — из внутренних. Однако в эти ткани дифференцируются не все прокамбиальные клетки. Один их слой между первичными ксилемой и флоэмой остается меристематическим, становясь камбием. У липы стебель деревянистый, т. е. образующий много вторичной ксилемы (вторичный рост стеблей рассматривается в гл. 23). После того как удлинение междоузлий завершается, в первичной флоэме здесь образуются волокна, называемые первичными флоэмными волокнами (см. рис. 23-10).
Внутренняя граница первичной ксилемы у липы резко оконтурена одним или двумя слоями более темных клеток сердцевины, состоящей в основном из паренхимных клеток и содержащей многочисленные крупные ходы, или каналы, со слизью (особыми углеводами). Аналогичные ходы образуются в первичной коре. По мере увеличения размеров коровых и сердцевинных клеток между ними развиваются многочисленные межклетники. Эти заполненные воздухом пространства очень важны для газообмена с атмосферой. Паренхимные клетки сердцевины и коры запасают различные вещества.
Стебель бузины
В стебле бузины (Sambucus canadensis) прокамбий и первичные проводящие ткани расположены отдельными тяжами вокруг сердцевины. Последняя, а также эпидерма и первичная кора очень сходны здесь по своему строению с обсуждавшимися на примере липы, поэтому ниже будет детально рассмотрено только развитие первичных проводящих тканей стебля.
На рис. 22-9, А показаны три прокамбиальных тяжа, в которых первичные проводящие ткани только начинают дифференцироваться. Левый тяж несколько старше двух остальных и содержит по меньшей мере по одному зрелому ситовидному и трахеальному элементу. Можно видеть, что зрелый ситовидный элемент находится в наружной части тяжа (около коры), а первый зрелый трахеальный элемент — в его внутренней части (на границе с сердцевиной). Если сравнить рис. 22-9, А и 22-9, В, то заметно появление более молодых ситовидных элементов ближе к центру стебля, тогда как ксилема дифференцируется в противоположном направлении.
Первые элементы первичных ксилемы и флоэмы (протоксилема и протофлоэма соответственно) растягиваются в процессе удлинения междоузлия и часто разрушаются. Как и у липы, стебли бузины одревесневают. В первом случае почти весь камбий происходит из прокамбиальных клеток между первичными ксилемой и флоэмой, а межпучковые зоны очень узкие. У бузины они относительно широкие, вследствие чего существенная часть камбия развивается из межпучковой паренхимы.
Стебли люцерны и лютика
Стебли многих двудольных с небольшим вторичным ростом или вообще без него являются травянистыми, или неодревесневшими (см. гл. 23). Примерами здесь могут служить стебли люцерны (Medicago) и лютика (Ranunculus).
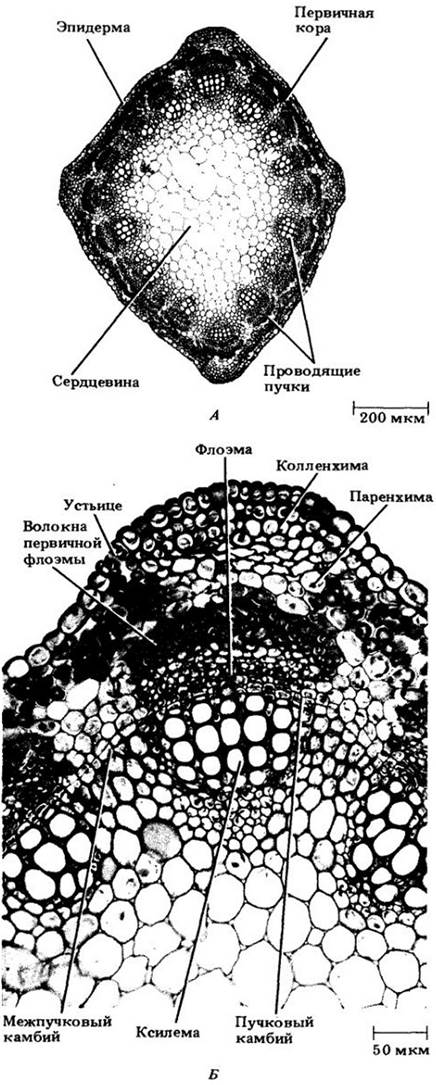
Люцерна — травянистое двудольное с некоторым вторичным ростом (см. рис. 22-7). Структура и развитие первичных тканей стебля у этого растения во многом сходны с известными для бузины и других древесных представителей его класса. Проводящие пучки разделены широкими межпучковыми зонами и окружают мощную сердцевину. Камбий по происхождению отчасти прокамбиальный, отчасти межпучковый, но вторичные проводящие ткани формируются главным образом внутри пучков. Межпучковый камбий откладывает преимущественно склеренхимные клетки в сторону ксилемы. В травянистом стебле лютика проводящие пучки сходны с известными у многих однодольных. После дифференцировки первичных проводящих тканей в них не остается прокамбия, вследствие чего никогда не образуется камбий и способность к дальнейшему росту утрачивается. Такие пучки (рис. 22-10) называют «закрытыми» в отличие от «открытых», в которых закладывается камбий. У большинства двудольных проводящие пучки второго типа и образуют хотя бы немного вторичных проводящих тканей.
Рис. 22-7. А. Поперечный срез стебля люцерны (Medicago sativa), двудольного растения с разделенными проводящими пучками. Б. Фрагмент того же самого стебля при большем увеличении
Стебель кукурузы
В травянистом стебле кукурузы (Zea mays) и многих других однодольных закрытые проводящие пучки образуют систему рассеянных по всей основной ткани тяжей (см. рис. 22-8).
Рис. 22-8. Стебель кукурузы (Zea mays). А. Поперечный срез междоузлия; видны многочисленные проводящие пучки, рассеянные среди основной ткани. Б. Поперечный срез узла молодого стебля кукурузы; видны горизонтальные прокамбиальные тяжи, связанные с вертикальными пучками. В. Расщепленный продольно зрелый стебель; основная ткань удалена, чтобы показать проводящую систему
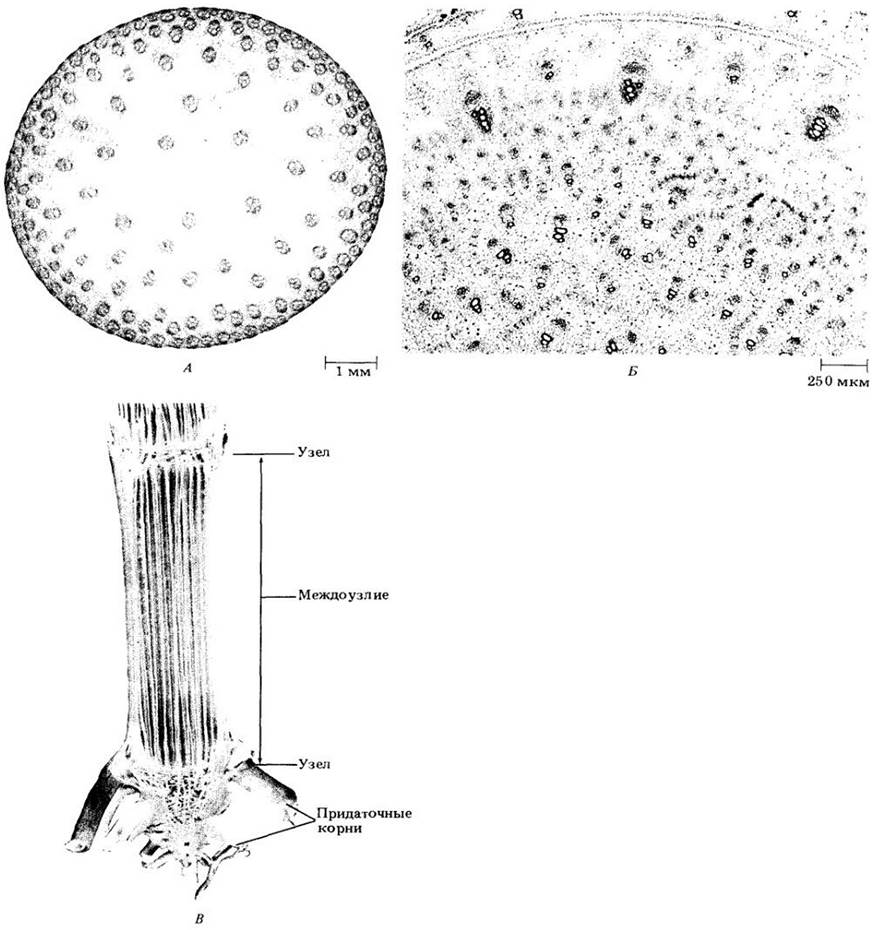
Рис. 22-9. Поперечные срезы стебля бузины (Sambucus canadensis) на стадии первичного роста. А. Очень молодой стебель с протодермой, основной меристемой и разделенными прокамбиальными тяжами. Тяж слева содержит по одному зрелому ситовидному (верхняя стрелка) и трахеальному (нижняя стрелка) элементу. Б. Более развитые первичные ткани. В. Стебель перед завершением первичного роста. Пучковый и межпучковый камбии еще не сформировались. Дальнейшие стадии роста стебля бузины представлены на рис. 23-8, 23-9 и 23-11
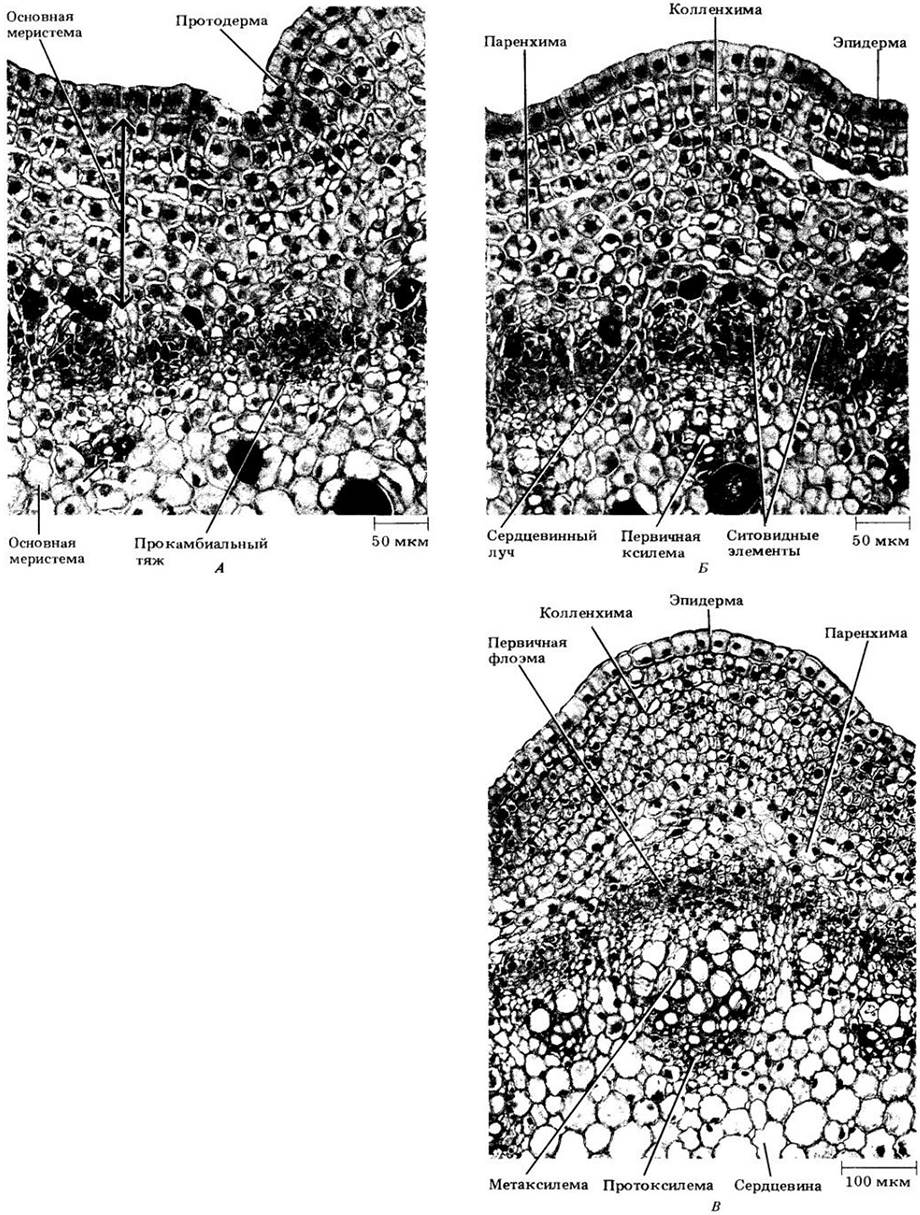
Рис. 22-10. Поперечный срез проводящего пучка лютика (Ranunculus), травянистого двудольного. Проводящие пучки этого растения закрытые, т. е. все прокамбиальные клетки в них дифференцированы и вторичных тканей не развивается. Первичные флоэма и ксилема окружены пучковым влагалищем из толстостенных склеренхимных клеток. Сравните этот пучок со зрелым проводящим пучком кукурузы, приведенным на рис. 22-11, В
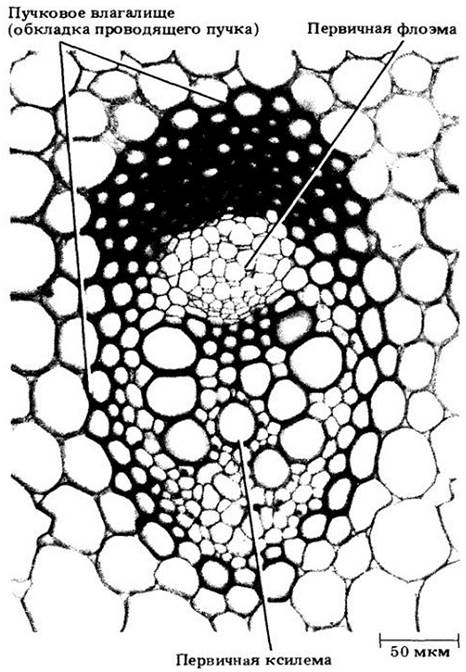
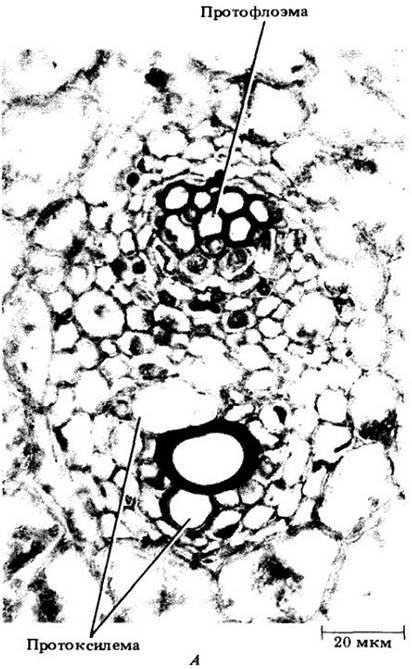
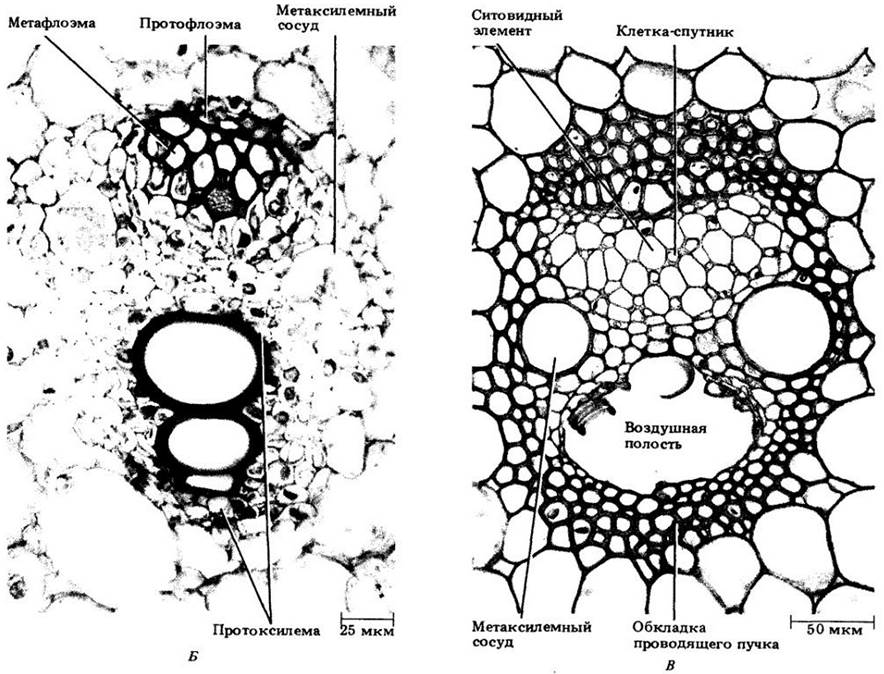
На рис. 22-11 показаны три стадии развития такого пучка. Как и в стебле двудольных, флоэма развивается из наружных клеток прокамбиального тяжа, а ксилема — из внутренних клеток, причем дифференцировка флоэмы и ксилемы идет в противоположных направлениях. На поперечных срезах видно, что первая дифференцируется центростремительно, а вторая — центробежно. Первые элементы флоэмы и ксилемы (протофлоэма и протоксилема) растягиваются и разрушаются в процессе удлинения междоузлий, в результате чего на ксилемной стороне пучка образуется крупная воздухоносная полость (рис. 22-11, В). Зрелый проводящий пучок, включающий два широких членика сосуда (сосуды метаксилемы) и флоэму (метафлоэму) из члеников ситовидной трубки и клеток-спутников, заключен в обкладку из склеренхимных клеток.
Рис. 22-11. Три стадии дифференцировки проводящих пучков кукурузы на поперечных срезах стебля. А. Зрелые элементы протофлоэмы и (два) протоксилемы. Б. Ситовидные элементы протофлоэмы разрушены; значительная часть метафлоэмы созрела. В протоксилеме три зрелых элемента, а два членика сосудов метаксилемы почти закончили растяжение. В. Зрелый проводящий пучок окружен влагалищем из толстостенных склеренхимных клеток. Метафлоэма состоит целиком из члеников ситовидных трубок и клеток- спутников. Часть проводящего пучка, занятая ранее протоксилемными элементами, теперь превратилась в крупную воздухоносную полость. Можно видеть по ее границе утолщения оболочек, разрушенных протоксилемных элементов
Взаимосвязь между проводящими тканями стебля и листа
Расположение проводящих пучков в стебле указывает на тесную связь в строении и развитии между ним и его боковыми придатками, т. е. листьями. Общий термин «побег» служит не только удобным обозначением для комплекса надземных вегетативных органов, но и отражает эту взаимосвязь.
Прокамбиальные тяжи стебля начинаются сразу же за апикальной меристемой под развивающимися листовыми примордиями, а иногда и под местом их будущего заложения еще до начала этого развития. По мере роста примордиев в длину тяжи дифференцируются по направлению к кончику листа. Таким образом, уже с самого начала прокамбиальная система последнего оказывается непрерывно связанной с прокамбием стебля.
В каждом узле один или более проводящих пучков идут от центрального цилиндра стебля через кору в лист (или листья) данного узла (рис. 22-12 и 22-13). В стебле пучок, направляющийся от продольного стеблевого пучка к основанию листа, где он связывается с проводящей системой последнего, называется листовым следом, а широкий промежуток, т. е. участок основной ткани, находящийся в центральном цилиндре над местом отхождения листового следа, называется листовым прорывом. В один лист может входить из стебля один или более листовых следов. Число междоузлий, которые они при этом пересекают, различно, т. с. длина листовых следов неодинакова.
Рис. 22-12. Схемы строения первичной проводящей системы стебля мари сизой (Chenopodium glaucum), обособленные пучки которой образуют цилиндр из взаимосвязанных тяжей вокруг сердцевины. А. Трехмерная схема расположения пучков в стебле. Б. Проводящая система развернута в плоскости. Можно видеть листовые следы, отходящие наружу от проводящего цилиндра, и их связь с проводящими пучками стебля. У этого вида в каждый лист входят три листовых следа
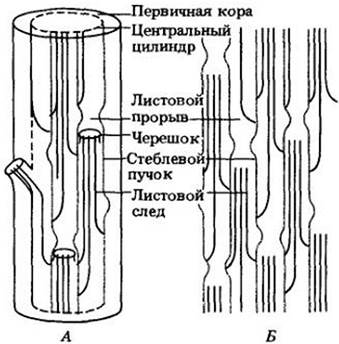
Рис. 22-13. Схемы продольного (А) и поперечных (Б) срезов побега табака (Nicotiana tabacum), показывающие взаимоотношения проводящих систем листа и стебля. «Непрерывный» центральный цилиндр здесь состоит из неодинаковых по ширине участков. В каждом узле в лист отходит единственный листовой след и присутствуют также следы ветвления, часто тесно связанные с листовыми. У табака от проводящей системы стебля к почке направляются два следа ветвления, а прорыв ветвления не отделен от листового
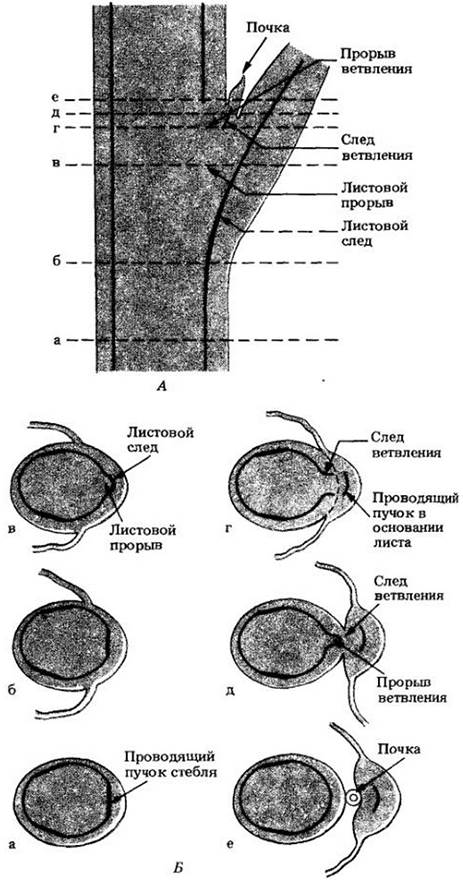
Если проследить ход проводящего пучка вверх и вниз по стеблю, выяснится, что он связан с несколькими листовыми следами, образуя так называемый симподий. В одних стеблях некоторые или все симподии взаимосвязаны, в то время как в других каждый из них представляет собой независимый проводящий комплекс. В любом случае структура проводящей системы стебля отражает расположение на нем листьев.
Морфология листа
Листья весьма разнообразны по форме и внутреннему строению. У двудольных они обычно состоят из плоской расширенной части, пластинки, и стеблевидного черешка (рис. 22-14). У основания некоторых листьев образуются мелкие чешуевидные или листовидные структуры, называемые прилистниками (рис. 22-15). Многие листья лишены черешков; их называют сидячими (рис. 22-16). У большинства однодольных и некоторых двудольных их основание расширено в охватывающее стебель влагалище (рис. 22-16, Б). У ряда злаков оно покрывает все междоузлие. Расположение листьев на стебле может быть спиральным (очередным), супротивным (попарным) или мутовчатым (по три или больше листьев в узле). Например, у шелковицы (Morus alba) и дуба (Quercus) листорасположение спиральное, у клена (Acer) — супротивное, у Veronicastrum virginicum — мутовчатое (рис. 22-14).
Рис. 22-14. Несколько примеров простых листьев. А. Шелковица белая (Morus alba). Б. Veronicastrum virginicum. В. Клен сахарный (Acer saccharum). Г. Клен серебристый (Acer saccharinum). Д. Дуб красный (Quercus rubra). Можно видеть спиральное, или очередное, листорасположение у шелковицы и мутовчатое у Veronicastrum. У клена оно супротивное, а у дуба спиральное, но здесь показаны только отдельные листья этих деревьев

Листья двудольных бывают простыми или сложными. В первом случае их пластинка не разделена на части, хотя может быть глубоколопастной (рис. 22-14), а во втором представлена отдельными листочками, каждый из которых обычно снабжен собственным мелким черешком (черешочком). Можно выделить перисто- и пальчато-сложные листья (рис. 22-17). У первых листочки располагаются двумя рядами по обе стороны оси — рахиса, представляющего собой разросшийся черешок. У пальчато-сложного листа они отходят от верхушки черешка и рахис отсутствует.
Поскольку листочки внешне напоминают простые листья, иногда эти структуры трудно различить. Удобно использовать два критерия: (1) в пазухе простого и сложного листа находится почка, а в пазухе листочка ее нет; (2) листья отходят от стебля в разных плоскостях, а листочки от рахиса — в одной.
Структура листа
Различия в структуре листьев покрытосеменных в значительной степени связаны с условиями существования растений; особенно важный фактор, влияющий на их форму и строение, — доступность воды. По потребностям в ней и соответствующим адаптациям виды обычно делят на мезофиты (нуждающиеся в высокой влажности почвы и относительно влажной атмосфере), гидрофиты (произрастающие в переувлажненных местообитаниях, часто полностью или частично погруженные в воду) и ксерофиты (приспособленные к аридным условиям). Эти экологические типы не имеют отчетливых границ, и часто листья имеют смешанные признаки. Независимо от формы листья покрытосеменных являются специализированными органами фотосинтеза и, как корни и стебли, состоят из систем покровных, основных и проводящих тканей.
Эпидерма
Основная масса эпидермальных клеток листа, как и у стебля, расположена плотно и покрыта кутикулой, снижающей потери воды (см. гл. 3 и с. 25). Устьица могут встречаться на обеих его сторонах, но обычно более многочисленны на нижней поверхности (рис. 22-18). У плавающих на поверхности воды листьев гидрофитов они могут встречаться только на верхней эпидерме (рис. 22-19), а на погруженных листьях обычно совсем отсутствуют. На листьях ксерофитов, как правило, больше устьиц, чем у других растений. Предполагают, что это обеспечивает более интенсивный газообмен в относительно редкие периоды благоприятного водоснабжения. У многих ксерофитов устьица погружены в углубления на нижней поверхности листа (рис. 22-20), которые могут быть выстланы многочисленными эпидермальными волосками. Две эти особенности, по-видимому, способствуют уменьшению потерь воды. Эпидермальные волоски, или трихомы, могут встречаться на любой поверхности листа или на обеих сразу. Их густой покров также снижает отдачу влаги.
У листьев двудольных устьица часто рассеяны по поверхности беспорядочно (рис. 22-21). Их развитие смешанное, т. с. формирующиеся и уже полностью дифференцированные устьица встречаются рядом на одном и том же развивающемся листе. У однодольных, устьица расположены рядами, параллельными оси листа (рис. 22-22), и развиваются от его верхушки к основанию.
Мезофилл
Именно мезофилл — основная ткань листа с крупными межклетниками и многочисленными хлоропластами — наиболее приспособлен для фотосинтеза. Межклеточные пространства связаны с атмосферным воздухом через устьица, обеспечивающие быстрый газообмен, необходимый для эффективного протекания этого процесса. У мезофитов мезофилл дифференцирован на палисадную и губчатую паренхиму. В первой клетки имеют столбчатую форму с длинными осями, ориентированными под прямыми углами к эпидерме. В губчатой паренхиме форма клеток неправильная (см. рис. 22-18, Б, Г). Хотя палисадная ткань кажется более плотной, чем губчатая, большая часть вертикальной поверхности ее клеток граничит с межклетниками, и в некоторых листьях общая площадь этих клеток может быть в два — четыре раза больше, чем в губчатой паренхиме. Хлоропласты также более многочисленны в палисадных клетках, в связи с чем здесь, по-видимому, сосредоточена основная фотосинтетическая активность.
Обычно палисадная паренхима располагается на верхней стороне листа, а губчатая — на нижней (см. рис. 22-18). У ксерофитов первая часто находится с обеих сторон листа, а у некоторых растений, например, кукурузы (см. рис. 7-19) и других злаков (см. рис. 22-25 — 22-27), клетки мезофилла более или менее одинаковые по форме и различия между двумя типами паренхимы отсутствуют.
Рис. 22-15. Перисто-сложный лист гороха (Pisum sativum). Обратите внимание на прилистники у его основания (у этого рода они часто крупнее, чем листочки) и тонкие усики на конце
Рис. 22-16. Сидячие (бесчерешковые) листья часто встречаются у двудольных, например, у Moricandia из семейства крестоцветных (А), но особенно характерны для злаков и других однодольных. Б. У кукурузы (однодольного растения) основание листа образует вокруг стебля влагалище, из которого выступает язычок — небольшой вырост листовой ткани
Рис. 22-17. Некоторые примеры сложных листьев. А. Пальчато-сложный у конского каштана павия (Aesculus pavia). Остальные листья — перисто-сложные. Б. Кария овальная (Caria ovala). В. Ясень пенсильванский (Fraxinus pennsylvanica var. subintegerrima). Г. Белая акация (Robinia pseudo-acacia). Д. Гледичия трехколючковая (Gleditsia tria- canthos). У этого вида каждый листочек подразделен на более мелкие листочки

Проводящие пучки
Мезофилл листа густо пронизан многочисленными проводящими пучками, или жилками, непосредственно связанными с проводящей системой стебля. У большинства двудольных они образуют разветвленную систему, в которой от более крупных жилок ответвляются все более мелкие. Такое жилкование называют сетчатым (рис. 22-23). Часто самая крупная (средняя) жилка тянется вдоль оси листа и вместе с прилегающей основной тканью выступает над его нижней поверхностью в виде гребня (рис. 22-18, А). В отличие от этого у большинства однодольных множество жилок примерно равного размера проходит вдоль листа параллельно друг другу. Такое жилкование называется параллельным (рис. 2-16, Б). При этом продольные жилки связаны между собой более мелкими, образуя сложную сеть (рис. 22-24).
Жилки содержат ксилему и флоэму, имеющие, как правило, первичное происхождение (средние, а иногда и другие крупные жилки у некоторых двудольных способны к вторичному росту). Окончания жилок у двудольных часто содержат только трахеальные элементы, хотя до их концов могут доходить как ксилемные, так и флоэмные элементы. Обычно ксилема находится на верхней стороне листа, флоэма — на нижней (рис. 22-18, А, Б).
Проводящие пучки, более или менее полностью погруженные в мезофилл, называют мелкими жилками, а сопровождающиеся выступами на нижней стороне листа — крупными жилками. Главную роль в отводе ассимилятов из клеток мезофилла играют первые. С увеличением размеров жилок их связь с мезофиллом ослабевает, и они все больше окружаются нефотосинтезирующими тканями гребней. Следовательно, функция сбора ассимилятов постепенно вытесняется главным образом транспортной.
Проводящие ткани жилок редко граничат с межклетниками мезофилла. Крупные жилки окружены паренхимой, содержащей мало хлоропластов, в то время как мелкие — одним и более слоями компактно расположенных клеток, образующих обкладку проводящего пучка (пучковое влагалище) (см. рис. 22-18 — 22-20) и часто напоминающих клетки мезофилла, в которые погружены эти жилки. Обкладка сопровождает пучок до самого окончания, так что проводящие ткани нигде не контактируют с межклетниками и все вещества, поступающие в проводящие ткани или выходящие из них, неизбежно проходят через ее клетки (см. рис. 22-18, Г). Таким образом, обкладка функционирует аналогично эндодерме корня.
Рис. 22-18. Срезы листа сирени (Syringa vulgaris). А. Поперечный через среднюю жилку. Б. Поперечный через участок пластинки. Можно видеть две. мелкие жилки. В. Парадермольный, т. е. примерно параллельный поверхности листа, срез. От верхней к нижней части этой микрофотографии срез все больше углубляется в ткани листа, в результате вверху можно видеть часть верхней эпидермы, а внизу — часть нижней. Хорошо заметно, что на последней больше устьиц. У сирени жилкование сетчатое. Г и Д. Увеличенные фрагменты рис. В. Г. Видна палисадная (вверху) и губчатая (внизу) паренхима. Вверху справа на этой же микрофотографии — окончание жилки; срез прошел через несколько трахеальных элементов, окруженных обкладкой пучка. Д. Участок нижней эпидермы с двумя трихомами (эпидермальными волосками), несколькими устьицами и многочисленными обычными эпидермальными клетками
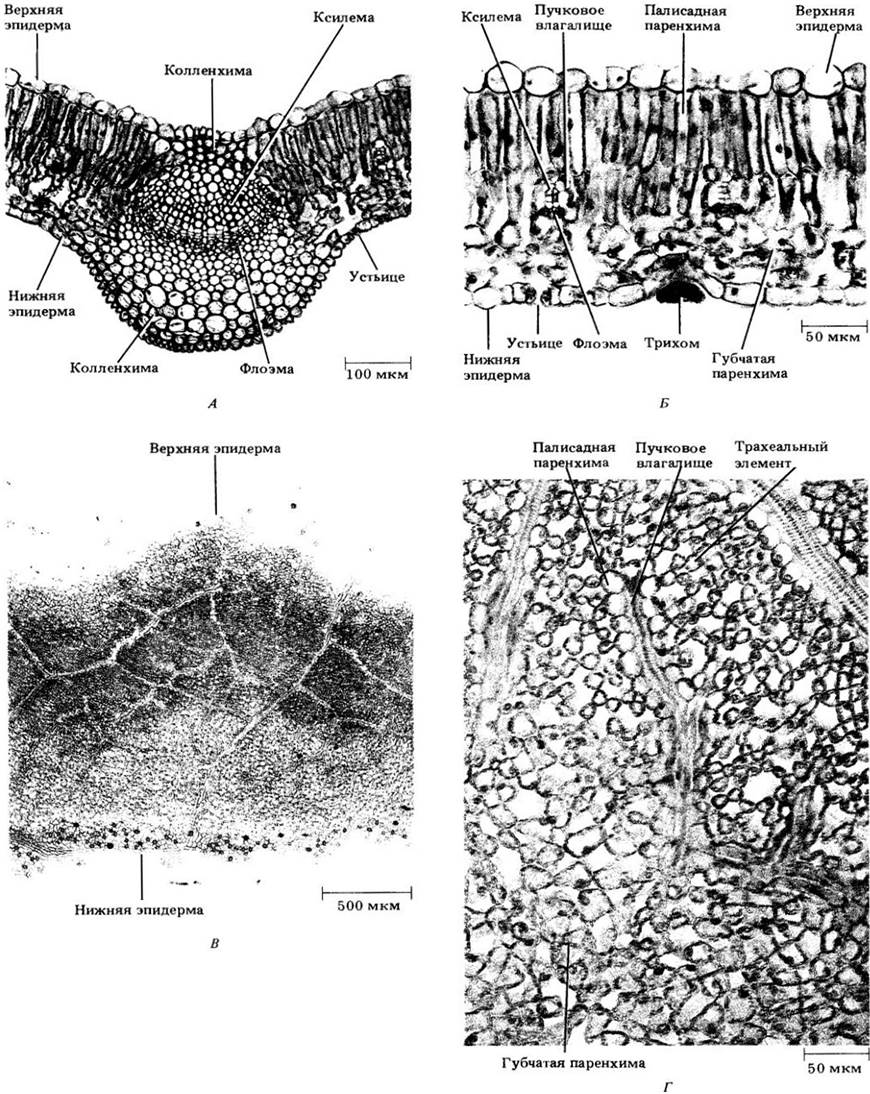
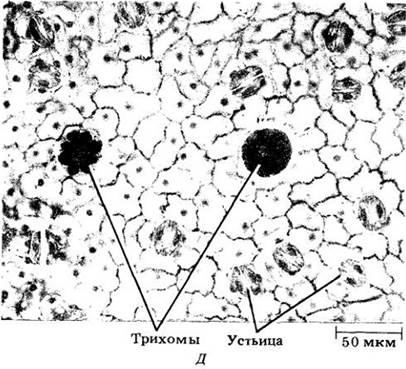
Рис. 22-19. Поперечный срез плавающего на поверхности воды листа кувшинки (Nymphaea odorata) с устьицами только на верхней эпидерме. Как это свойственно гидрофитам, проводящая ткань сильно редуцирована, особенно ксилема. Палисадная паренхима состоит из нескольких слоев клеток над губчатой паренхимой
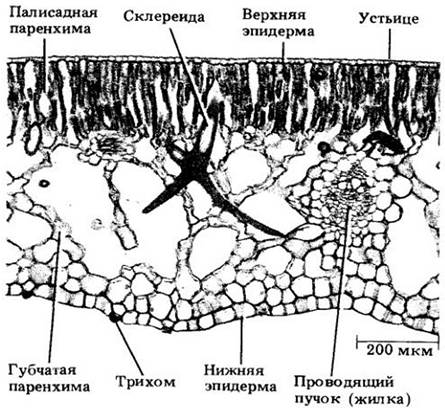
У многих листьев обкладки проводящих пучков связаны с верхней и (или) нижней эпидермой клетками, напоминающими клетки самой обкладки (рис. 22-20). Такие ответвления от нее называют продолжениями обкладки проводящего пучка. Они механически укрепляют лист, а, кроме того, у двудольных, по-видимому, подводят к эпидерме воду.
Рис. 22-20. Поперечный срез листа олеандра (Nerium oleander). Этот вид — ксерофит, что находит свое отражение в структуре листа. Можно видеть очень толстую кутикулу, покрывающую многослойную эпидерму на верхней и нижней его поверхностях. Устьица и трихомы приурочены к углублениям в нижней эпидерме, так называемым устьичным криптам
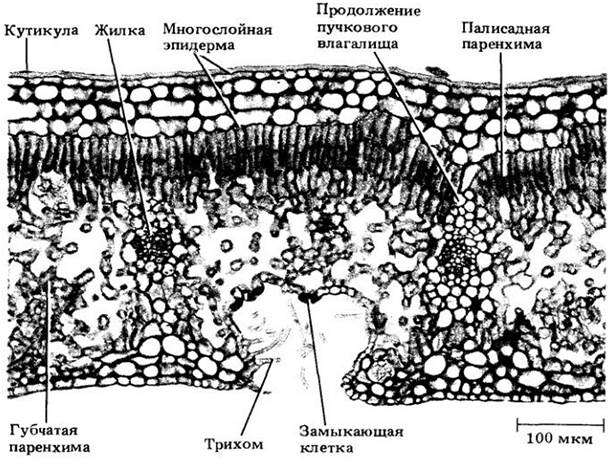
Сама эпидерма в значительной мере обеспечивает прочность листа благодаря плотному расположению своих клеток и наличию кутикулы. Крупные жилки у двудольных часто окружены колленхимными или склеренхимными клетками, также выполняющими опорную функцию. У однодольных, жилки могут быть окаймлены волокнами. Клетки колленхимы и волокна иногда располагаются также по краю листа двудольных и однодольных соответственно.
Листья злаков
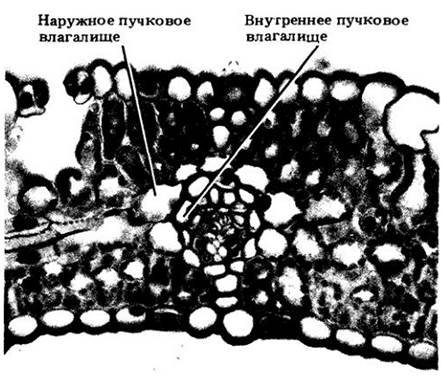
После открытия у сахарного тростника С4-фотосинтеза (рис. 7-18) множество работ было посвящено сравнительной анатомии листьев злаков и связи ее с различными типами этого процесса. Выяснено, что листья С3- и С4-злаков анатомически довольно четко различаются. Например, у С4-видов клетки мезофилла и обкладки пучков, как правило, образуют вокруг последних два концентрических слоя (см. поперечный срез, рис. 22-25). Плотно расположенные очень крупные паренхимные клетки обкладки С4-злаков содержат много объемистых хорошо заметных хлоропластов. Концентрическое расположение клеточных слоев мезофилла и обкладки называют кранц-анатомией (от немецкого слова Kranz, означающего «венок»).
Рис. 22-21. Вид листа картофеля (Solanum tuberosum) в сканирующем электронном микроскопе. Заметно неупорядоченное расположение устьиц, свойственное двудольным. Замыкающие клетки у картофеля почковидные и не связаны с побочными
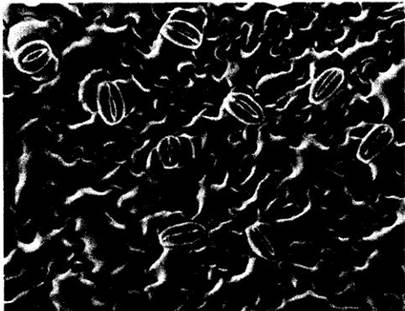
Рис. 22-22. Вид листа кукурузы в сканирующем электронном микроскопе. Параллельное расположение устьиц типично для однодольных. У кукурузы каждая пара узких замыкающих клеток связана с двумя побочными, по одной с каждой стороны устьица

Рис. 22-23. Просветленный лист (удален хлорофилл) тополя дельтовидного (Роpulus deltoides) при двух увеличениях. У этой породы характерное для двудольных, сетчатое жилкование. Небольшие участки мезофилла, ограниченные жилками, называются ареолами. Все метки мезофилла находятся вблизи жилок. Вода с растворенными минеральными веществами поступает сюда по ксилеме, а синтезированные органические молекулы отводятся по флоэме
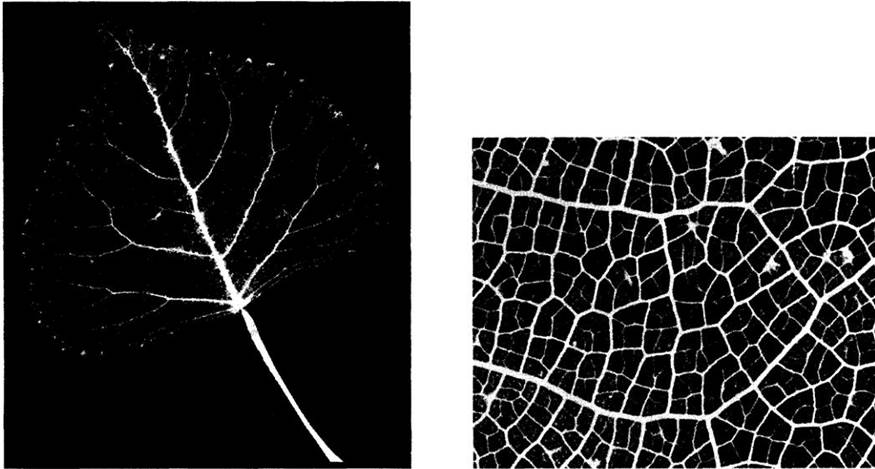
Рис. 22-24. Парадермальный срез листа кукурузы, демонстрирующий характерное для однодольных, параллельное жилкование. Заметны многочисленные поперечные жилки, связывающие воедино систему продольных (параллельных) жилок

Рис. 22-25. Поперечный срез листа сахарного тростника (Saccharum officinarum). Как это свойственно С4-злакам, клетки мезофилла расположены радиально вокруг обкладок проводящих пучков, состоящих из крупных клеток с хлорофиллом

У С3-растений клетки мезофилла и обкладки не имеют такого концентрического расположения. Более того, относительно мелкие клетки паренхиматозной обкладки содержат здесь некрупные хлоропласты и при небольшом увеличении кажутся пустыми и светлыми. Обычно у С3-видов присутствует еще и внутренняя так называемая местомная обкладка из более или менее толстостенных клеток (рис. 22- 26).
Рис. 22-26. Поперечный срез листа пшеницы (Triticum aestivum) — типичного С3-злака. Клетки мезофилла не располагаются вокруг обкладки проводящего пучка радиально. Сама она состоит из двух слоев паренхимных клеток: наружного — из относительно тонкостенных и внутреннего (местомного) — из толстостенных
Другое устойчивое структурное различие между листьями С3- и С4-злаков заключается в расстоянии между жилками, т. е. между обкладками соседних проводящих пучков. У С4- злаков оно соответствует только двум — четырем клеткам мезофилла, у С3-злаков всегда большему их числу (в одной работе указывается в среднем 12).
Из листьев С4-растений продукты фотосинтеза отводятся обычно быстрее и полнее, чем у С3-видов. Причины этого неизвестны, но высказано предположение, что расстояние между клетками мезофилла и флоэмой проводящих пучков может влиять на скорость поступления веществ в ситовидные трубки.
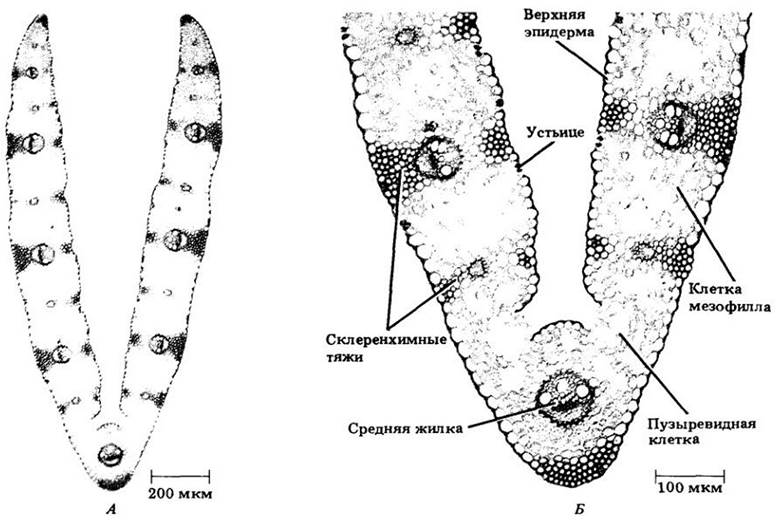
Эпидерма злаков состоит из клеток различного типа. Большинство их узкие и вытянутые. Некоторые, выделяющисся крупными размерами и называемые пузыревидными, располагаются продольными рядами и, как предполагают, участвуют в складывании (скручивании) и развертывании (раскручивании) листьев (рис. 22-27). Толстостенные замыкающие клетки устьиц связаны с побочными клетками (см. рис. 22-22, а также рис. 20-20).
Рис. 22-27. Поперечный срез листа С3- злака мятлика однолетнего (Роа annua). Б. Увеличенная часть среза со средней жилкой. В листе злака мезофилл не дифференцирован на палисадную и губчатую паренхиму. Выше и ниже жилок обычно находятся тяжи склеренхимных клеток. Эпидерма содержит крупные пузыревидные клетки, которые, как считают, участвуют в складывании и разворачивании листьев. У мятлика на рисунке они частично утратили тургор, поэтому лист сложен. По-видимому, возрастание тургора этих клеток заставило бы его развернуться
Развитие листа
Первый структурный признак закладки листа у большинства покрытосеменных — начало периклинальных делений под протодермой в периферической зоне апекса побега. Сочетание растяжения клеток и дальнейших их делений вскоре приводит к образованию выступа, или листового бугорка (рис. 22-28, A), в то время как на апексе в определенное время и в определенном месте по отношению к ранее заложившимся листьям возникают новые центры митотической активности. До или во время формирования бугорка под молодым листовым примордием появляется прокамбиальный тяж.
Рис. 22-28. Ранние стадии развития листа Coleus blumei на продольных срезах верхушки побега. Листья у этого вида располагаются в узлах попарно и супротивно друг другу (см. рис. 22-2). А. По краям апикальной меристемы можно видеть два мелких супротивных выступа — листовых бугорка. Ниже, в пазухах двух молодых листьев, можно видеть зачатки почек. Б. Два прямостоячих клиновидных листовых примордия, развившиеся из листовых бугорков. Заметны входящие в них про- камбиальные тяжи. Почечные примордии (ниже) более развиты, чем на рис. А. По мере роста листовых примордиев (В) в них продолжают развиваться про- камбиальные тяжи, отходящие от проводящих пучков стебля. Трихомы, или эпидермальные волоски, образуются из некоторых протодермальных клеток очень рано, задолго до окончания дифференцировки протодермы в эпидерму
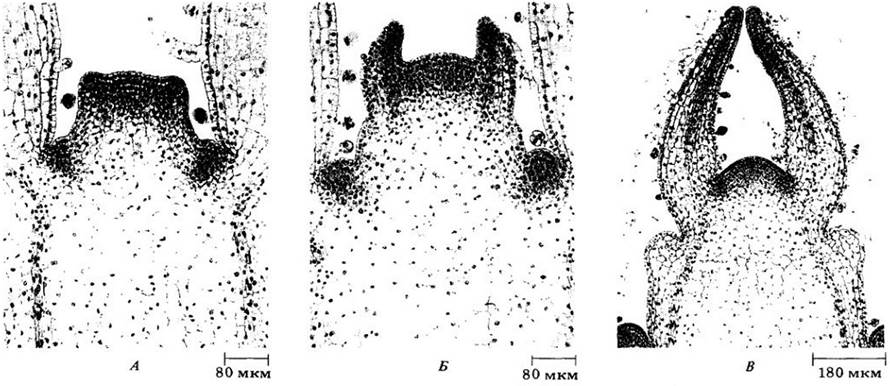
По мере роста в длину листовой бугорок развивается в прямостоячий клиновидный листовой примордий (рис. 22- 28, Б). У двудольных на нем примерно с противоположных сторон от оси вскоре появляются зоны меристематической активности. Эти области, начинающие формировать листовую пластинку, называются маргинальными меристемами (рис. 22-29 и 22-30).
Апикальный рост примордия непродолжителен. Лист увеличивается в длину главным образом за счет интеркалярного роста, т. е. делений и (в основном) растяжения клеток всей пластинки. В результате активности маргинальной меристемы довольно рано устанавливается определенное число клеточных слоев мезофилла пластинки, хотя в ходе дальнейшего развития оно может увеличиться. Различия в скорости делений и растяжения клеток разных слоев пластинки приводят к образованию многочисленных межклетников и типичной структуры мезофилла. Как правило, первой прекращает рост верхушка листа, а последним — его основание. По сравнению со стеблем большинство листьев растет недолго. Неограниченный или продолжительный рост вегетативной апикальной меристемы называют недетерминированным, а ограниченный (свойственный листу и апексам цветков) — детерминированным.
По мере удлинения клиновидного листового примордия про камбиальный тяж все дальше проникает в него (рис. 22- 28, В), образуя систему крупных жилок, отходящих от главной, или средней, жилки (рис. 22-29). У верхушки листа закладываются более мелкие жилки, которые развиваются по направлению к его основанию, без перерывов переходя в более крупные. Таким образом, верхушка листа цервой приобретает полную систему жилкования, что отражает общую последовательность развития этого органа «сверху вниз».
Рис. 22-29. Схемы продольных и поперечных срезов, иллюстрирующих ранние стадии развития листа табака (Nicotiana tabacum). А. Молодой клиновидный листовой примордий без пластинки. Б. На противоположных его сторонах началась активность маргинальной меристемы, ведущая к формированию пластинки. Присутствие такой меристемы в листе табака еще недавно ставилось под сомнение. Как и апикальный рост листового примордия, ее деятельность может продолжаться сравнительно недолго. В. По мере роста пластинки становятся заметными главные боковые жилки (узкие светлые участки), отходящие от средней жилки. Г. Примордий стал выше. С дальнейшим ростом пластинки вдоль крупных жилок появляются выросты паренхимной ткани (гребни). Мелкие жилки начинают развиваться у верхушки листа. Самые нижние рисунки справа изображают поперечные срезы «крылатого» черешка. Сравните эти схемы со срезами на рис. 22-30
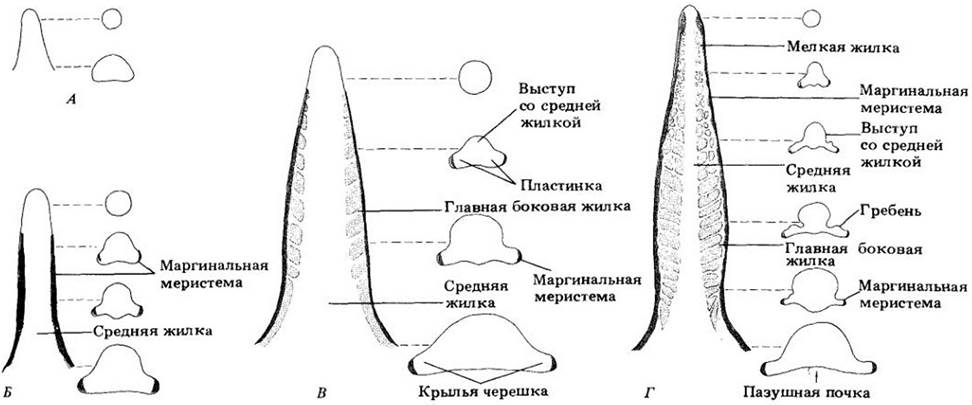
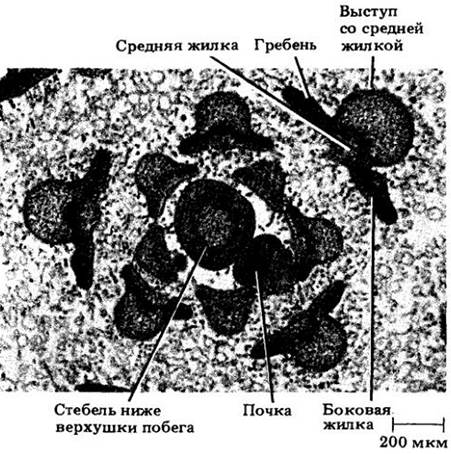
Рис. 22-30. Поперечный срез под апикальной меристемой развивающихся листьев табака, сгруппированных вокруг верхушки побега. Чем моложе листья, тем ближе они к оси. Примордий сначала не дифференцирован на среднюю жилку и пластинку. Можно видеть некоторые ранние стадии их развития (сравните эти срезы со схемами на рис. 22-29). Вокруг развивающихся листьев видны фрагменты многочисленных трихомов
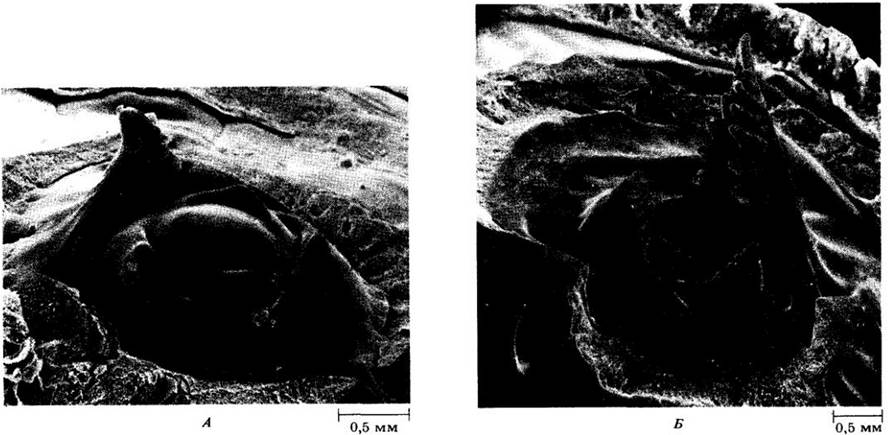
На рис. 22-31 представлено трехмерное изображение апекса побега и некоторых ранних стадий развития листа сельдерея (Apium graveolens). Листовые примордии и молодые листья расположены здесь спирально, причем более молодые листья находятся ближе к центру апекса. На рис. 22-31, А можно видеть два листовых бугорка справа и слева у его верхушки. Листовой примордий, развиваясь из бугорка, расширяется в виде раковины и благодаря быстрому относительно недолгому верхушечному росту образует черешково-рахисную часть будущего перисто-сложного листа. Ниже верхушки по обе стороны рахиса — сначала у его основания, а затем все ближе к верхушке — возникают выросты, из которых впоследствии формируются листочки. На первых выростах со временем появляются выросты второго порядка, становящиеся либо долями первичных листочков, либо отдельными вторичными листочками. Их можно видеть у относительно крупного зачаточного листа на рис. 22-31, Б. Каждый листочек развивается аналогично простому листу.
Рис. 22-31. Вид ранних стадий развития листа на апексе побега сельдерея (Apium graveolens) в сканирующем электронном микроскопе. А. Чем ближе развивающийся лист к центру апекса, тем он моложе. Б. Два листа (справа и слева) более развиты, чем любой из представленных на рис. А
Световые и теневые листья
Факторы окружающей среды, особенно свет, действуя на развивающиеся листья, могут оказывать существенное влияние на их окончательные размеры и толщину. У многих видов листья, выросшие при высокой освещенности (световые), мельче и толще, чем теневые, сформировавшиеся при меньшем количестве света. Увеличение толщины световых листьев связано главным образом с усиленным развитием палисадной паренхимы. Проводящая система в них также более протяженная, стенки эпидермальных клеток толще, а кроме того, гораздо выше отношение внутренней поверхности мезофилла к площади листовой пластинки. Влияние этих различий проявляется в том, что, хотя у обоих типов листьев интенсивность фотосинтеза одинакова при низкой освещенности, теневые листья не приспособлены к яркому свету и, следовательно, фотосинтезируют в таких условиях гораздо слабее световых.
Поскольку освещенность в различных частях кроны деревьев весьма неодинакова, здесь можно обнаружить крайние формы листьев обоих типов. Световые и теневые листья встречаются также у кустарников и травянистых растений. Образование того или иного типа можно стимулировать, выращивая растения при определенной освещенности.
Опадение листьев
У многих видов нормальному отделению листа от стебля, т. е. процессу опадения, предшествуют структурные и химические изменения вблизи основания черешка (см. с. 102), приводящие к образованию отделительной зоны (рис. 22- 32). У древесных двудольных здесь можно различить два слоя: отделительный и защитный. Первый состоит из относительно коротких клеток со слабо развитыми утолщениями клеточных оболочек, что делает его непрочным. Второй формируется за счет отложения суберина в клеточных оболочках и межклетниках под отделительным слоем. После опадения листа защитный слой заметен на стебле в виде листового рубца (см. рис. 23-18). Связанные с опадением гормональные факторы обсуждаются в гл. 24.
Рис. 22-32. Отделительная зона у листа клена (Acer). Продольный срез через основание черешка

Переход между проводящими системами корня и побега
Как указывалось, в предыдущей главе, различия между органами растений заключаются прежде всего в относительном расположении проводящих и основной тканей. Например, в корнях двудольных проводящие ткани образуют, как правило, сплошной цилиндр, окруженный первичной корой. Кроме того, тяжи первичной флоэмы чередуются с радиальными лучами первичной ксилемы. В стебле же проводящие ткани образуют цилиндр из отдельных пучков, расположенных вокруг сердцевины, с флоэмой на наружной их стороне и ксилемой на внутренней. Понятно, что где-то в первичном теле растения один тип структуры должен смениться другим. Это происходит постепенно в особой так называемой зоне перехода.
Как говорилось в гл. 19, побег и корень закладываются в эмбриогенезе как единая непрерывная структура. Следовательно, переориентация проводящих тканей происходит еще в оси молодого растения. Она начинается при закладке в зародыше прокамбия и завершается вместе с дифференцировкой различно расположенных его тканей в проростке. Непрерывная связь между проводящими системами побега и корня поддерживается на протяжении всей жизни растения.
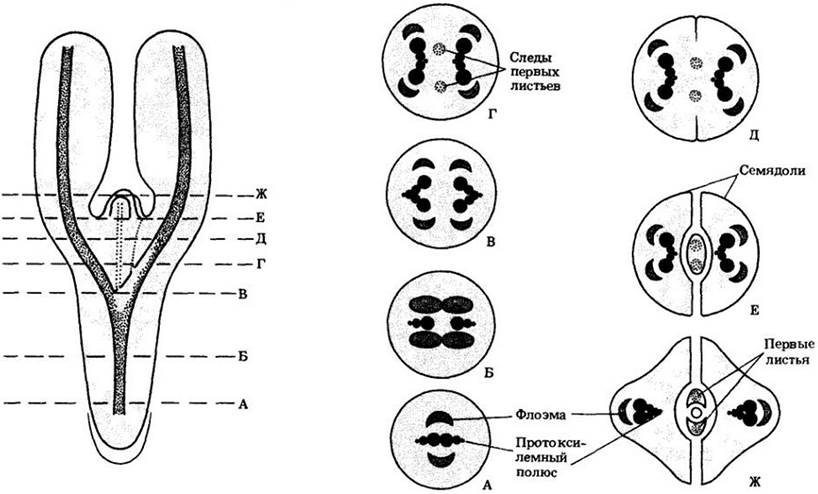
Структура зоны перехода может быть очень сложной и у разных видов сильно различается. У большинства голосеменных и двудольных перестройка проводящей системы наблюдается между корнем и семядолями. На рис. 22-33 изображена зона перехода, типичная для двудольных. Можно видеть диархную (с двумя протоксилемными полюсами) структуру корня, разветвление и переориентацию первичных ксилемы и флоэмы, приводящие в верхней части оси к образованию сердцевины, а также следы первых листьев эпикотиля.
Рис. 22-33. Зона перехода, связывающая корень и семядоли, у проростков двудольных с диархным корнем. Первичная проводящая система представлена в нем единственным цилиндром проводящей ткани. В гипокотиль-корневой оси эта система разветвляется, направляясь в семядоли, а ксилема и флоэма в пределах этой оси переориентируются
Развитие цветка
Меристематическая активность апекса вегетативного побега завершается развитием цветка или соцветия.
В процессе перехода к цветению в этом апексе происходят последовательные физиологические и структурные изменения, превращающие его в репродуктивный апекс. Следовательно, цветение можно рассматривать как стадию развития этой зоны побега и растения в целом. Поскольку репродуктивный апекс отличается детерминированным ростом, цветение однолетников означает приближение конца их жизненного цикла. У многолетников оно может происходить неоднократно. Известно, что в индукции цветения участвуют факторы окружающей среды, включая длину светового дня и температуру (см. гл. 25).
Переходу от вегетативного к цветковому апексу часто предшествуют удлинение междоузлий и раннее развитие под ним латеральных почек. Сам апекс заметно увеличивает митотическую активность, что сопровождается изменениями в его размерах и структуре: сначала относительно мелкий и организованный по типу туники—корпуса, он становится расширенным и куполообразным.
Закладка и ранние стадии развития чашелистиков, лепестков, тычинок и плодолистиков протекают почти так же, как у листьев. Обычно первыми закладываются чашелистики, затем лепестки, тычинки и наконец плодолистики (рис. 22-34). Этот типичный порядок появления частей цветка может быть в некоторых случаях изменен, но взаиморасположение их в пространстве всегда одинаково (рис. 22-35 и 22-36).
Рис. 22-34. Микрофотографии в сканирующем электронном микроскопе некоторых стадий развития обоеполого цветка Neptunia pubescens (бобовое) с радиальной симметрией и круговым расположением частей. Верхушки противолежащих брактей на рис. Б — И удалены. На рис. Е — И удалено также большинство чашелистиков и лепестков. А. Апекс цветка (а) в пазухе брактеи (б). Б. Пять примордиев чашелистиков (ч) вокруг апекса цветка. В. Пять примордиев лепестков (л) вокруг апекса цветка чередуются с чашелистиками (ч). Развиваясь. чашелистики образуют трубку чашечки. Г. Пять тычиночных примордиев (стрелки) вокруг апекса цветка чередуются с лепестками (л). Д. Вокруг апекса цветка, чередуясь с членами первого (внешнего) круга тычинок (т1), заложился второй (внутренний) их круг (т2) (стрелка). В центре цветочного апекса обособилась карпель (к). Теперь все части цветка налицо. Е. На корпели образуется продольная борозда, которая разовьется в гнездо завязи. Тычинки наружного круга (т1) начинают дифференцироваться на пыльники и тычиночные нити. Ж. Карпель начинает дифференцироваться на столбик и завязь (з). 3. Цветок на более поздней стадии с двумя кругами тычинок. И. Цветок на еще более поздней стадии; несколько тычинок удалено, чтобы показать карпель, дифференцированную на завязь (з), столбик и рыльце (стрелка)
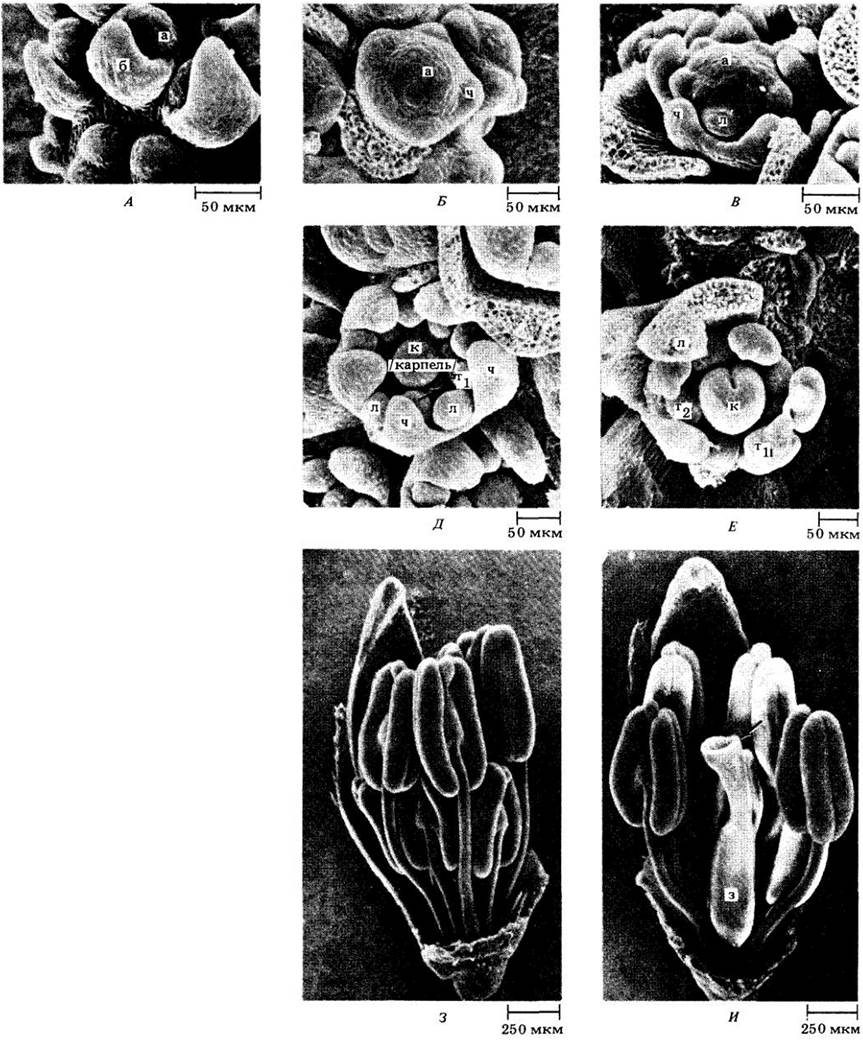
Рис. 22-35. Продольный срез молодой корзинки мелколепестника (Erigeron) из сложноцветных. Соцветие содержит многочисленные цветковые примордии. Более развитые из них находятся ближе к периферии соцветия, более молодые — в центре
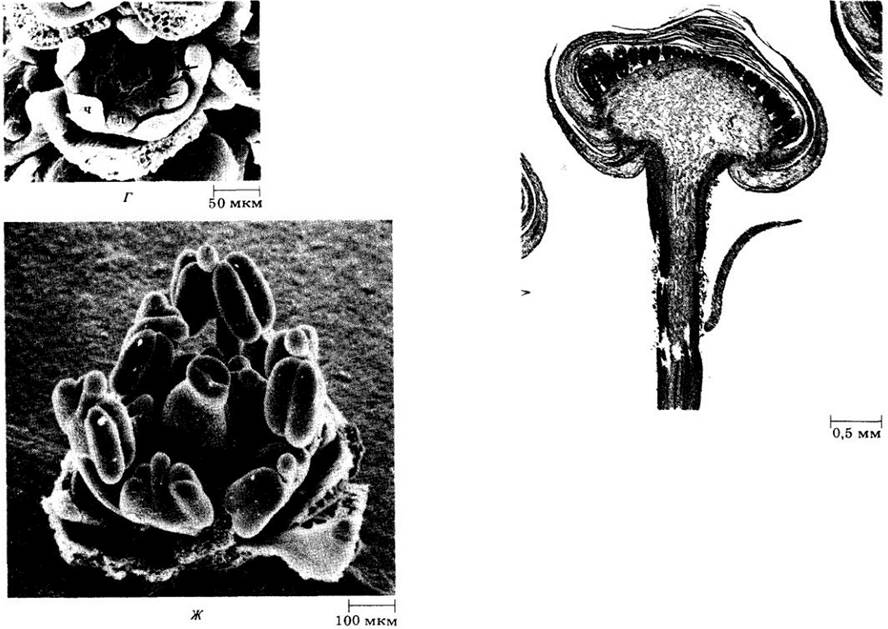
Рис. 22-36. Стадии развития цветка мелколепестника на продольных срезах. А. Цветковый примордий с развивающимся венчиком. Б. Закладка приросших к венчику тычинок. В. Цветок с развивающимся венчиком, тычинками и карпелями (продолжение рис. 22-36 см. с. 66)
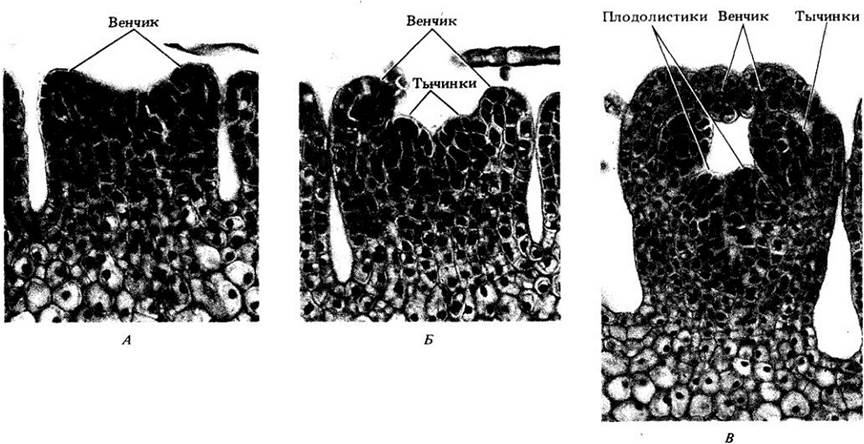
Рис. 22-36. Г, Д. Цветки с развивающимися семязачатками и столбиками. Две корпели образуют свод над полостью, заключающей семязачаток, а затем вытягиваются в сплошной столбик с двулопастным рыльцем. Чашечка, или nanny с, закладывается примерно в то же время, что и пыльники, но развивается медленно. Е. Цветки почти полностью развиты. В пыльниках можно различить пыльцевые зерна. Цветок мелколепестника имеет нижнюю завязь
Они могут оставаться раздельными в течение всего развития или объединяться внутри отдельных кругов (срастание) и между кругами (прирастание).
Типичная структура цветка и некоторые ее варианты обсуждались в гл. 18.
Модификации листа и стебля
Стебли и листья могут видоизменяться и выполнять нетипичные для себя функции. Одна из наиболее распространенных модификаций — образование усиков, служащих для закрепления. В некоторых случаях они представляют собой видоизмененные стебли. Например, у плюща (Hedera) усики образуют на своей верхушке расширенные чашевидные присоски. У винограда (Vitis) (рис. 22-37) и девичьего винограда (Parthenocissus quinquefolia) это также модифицированные стебли, обвивающиеся вокруг опоры. У винограда на усиках иногда развиваются мелкие листья и цветки.
Рис. 22-37. Усики винограда — видоизмененные побеги

Однако в большинстве случаев усики относятся к листовым модификациям. У бобовых, в частности гороха огородного (Pisum sativum), они соответствуют терминальной части перисто-сложного листа (см. рис. 22-15). Однако в этом семействе их образуют лишь немногие растения. У земляного ореха (Arachis hypogea), например, известно другое интересное приспособление. После оплодотворения тычинки и венчик опадают, а междоузлие между завязью и цветоложем начинает удлиняться, по мере вытягивания изгибается вниз и погружает на несколько сантиметров в землю развивающийся плод, где тот и созревает. Если завязь остается над поверхностью почвы, она завядает и спелых семян не образуется.
Ветви, принимающие форму листьев, называют кладофиллами, или кладодиями. Нитевидные листоподобные ветви спаржи (Asparagus officinalis) — типичный их пример (рис. 22-38). Ее толстые и мясистые надземные побеги («стрелки») съедобны. Чешуйки на них — настоящие листья. В ходе дальнейшего роста в пазухах этих крохотных невзрачных чешуек развиваются кладодии, функционирующие как фотосинтезирующие органы. У некоторых кактусов ветви напоминают листья (рис. 22-39).
Рис. 22-38. Нитевидные ветви обычной съедобной спаржи (Asparagus officinalis) напоминают листья. Такие модифицированные стебли называют кладодиями

Рис. 22-39. Ветви безыглого кактуса Epiphyllum напоминают листья, но на самом деле являются модифицированными стеблями, кладодиями

Иногда листья видоизменены в сухие и твердые иглы, не способные к фотосинтезу. Термины «игла» и «колючка» часто используются как взаимозаменяемые, но по происхождению вторые представляют собой модифицированные ветви, развивающиеся в пазухах листьев (рис. 22-40). Обычно синонимом этих терминов считают и «шип». Однако он не относится к модификациям стебля и листа, а представляет собой небольшой более или менее вытянутый острый вырост коры и эпидермы (пример — шипы на стебле розы). Колючки, иглы и шипы — защитные структуры, препятствующие поеданию растений животными-фитофагами. Высокоспециализированные взаимоотношения деревьев и растительноядных известны у так называемых «рогатых» акаций, иглы1 которых служат убежищем для муравьев, которые убивают других насекомых, пробующих питаться этими растениями (см. с. 256).
1 Предлагаемое здесь строгое различение терминов в зависимости от происхождения структуры в советской литературе не принято, поэтому «иглы» акаций (видоизмененные прилистники) или кактусов называют также колючками, а иногда и шипами. В морфологии уточняют «колючка листового происхождения» и т. д. — Прим. ред.
Рис. 22-40. A. Иглы этого кактуса (Ferocactus melocactiformis) — модифицированные листья. Б. Колючки — модифицированные ветви, образующиеся, как видно на этой фотографии боярышника (Crataegus sp.), в пазухах листьев
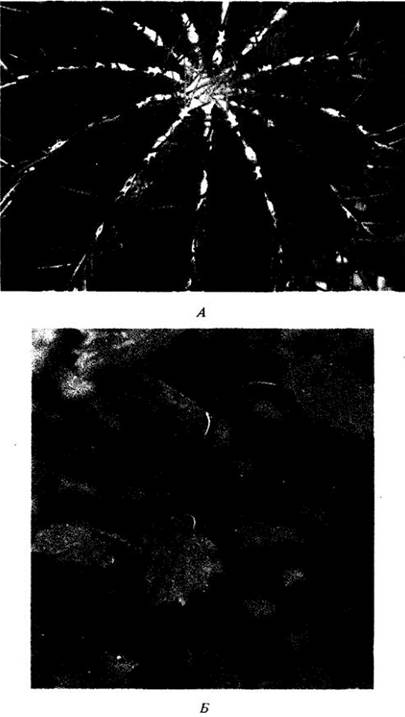
Очень эффектны модифицированные или специализированные листья плотоядных растений, например, саррацении, росянки и венериной мухоловки, служащие для ловли и переваривания насекомых. Образующиеся в результате питательные вещества поглощаются растением (см. гл. 26).
Запасание питательных веществ
Стебли, как и корни, служат для запасания питательных веществ. По-видимому, самым распространенным типом специализированных запасающих стеблей является клубень, например, у картофеля (Solarium tuberosum). У этого растения, если выращивать его из семян, они развиваются на концах столонов (тонких стеблей, растущих параллельно поверхности почвы). Однако если для размножения использовать разрезанные клубни, то новые такие же структуры формируются на концах длинных тонких корневищ, т. е. подземных стеблей (рис. 22-41). За исключением проводящих тканей, почти вся масса клубня под перидермой (кожицей) представлена запасающей паренхимой. Так называемые «глазки» картофеля — это углубления с группами почек, представляющие собой пазухи чешуевидных листьев.
Луковица — это по существу крупная почка, в которой находится небольшой конический стебель с многочисленными прикрепленными к нему модифицированными листьями. Они чешуевидные, с утолщенными основаниями, где откладываются питательные вещества. Придаточные корни отходят от основания стебля («донца»). Примеры растений с такими структурами — лук (рис. 22-42, Б) и лилия.
Рис. 22-41. Картофель (Solanum tuberosum) с клубнями, прикрепленными к корневищу, т. е. подземному стеблю
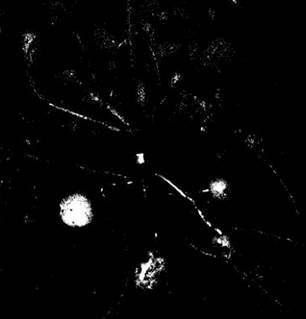
Рис. 22-42. Примеры модифицированных листьев и стеблей. А. Мясистый запасающий стебель кольраби (Brassica oleracea var. caulorapa). Б. Луковица лука репчатого (Allium сера) состоит из конического стебля и прикрепленных к нему чешуевидных листьев, содержащих запасные питательные вещества и составляющих съедобную часть этого видоизмененного побега. В. Клубнелуковица гладиолуса (Gladiolus grandiflorus) представляет собой мясистый стебель с мелкими пленчатыми листьями
Клубнелуковицы, хотя внешне и напоминают луковицу состоят преимущественно из тканей стебля. Их листья обычно тонкие и гораздо мельче, чем у луковиц, поэтому запасные вещества накапливаются здесь в мясистой стеблевой части. Клубнелуковицы образуют такие всем известные растения, как гладиолус (рис. 22-42, В), крокус, цикламен.
Кольраби (Brassica oleracea var. caulorapa) — пример съедобного растения с мясистым запасающим стеблем. Короткий и толстый, он поднимается над землей и несет несколько листьев с очень широкими основаниями (рис. 22-42, А). Обыкновенная, или кочанная, капуста (Brassica olerácea var. capitata) находится с кольраби в близком родстве. Ее так называемый «кочан» состоит из короткого стебля с многочисленными толстыми, перекрывающими друг друга листьями. Кроме верхушечной почки, здесь можно найти несколько хорошо развитых пазушных почек.
У некоторых растений толстыми и мясистыми становятся черешки листьев. Хорошо известные примеры — сельдерей (Apium graveolens) и ревень (Rheum rhaponticurn).
Накопление воды: суккулентность
Суккулентами называют растения с сочными тканями, специализированными для запасания воды. Большинство их, например, американские кактусы, внешне похожие на них африканские молочаи (Euphorbia) (см. Приложение 3 к этой главе), а также агавы (Agave), обычно произрастает в аридных регионах, где способность накапливать воду является жизненной необходимостью. Зеленые мясистые стебли кактусов служат одновременно фотосинтезирующими и запасающими органами. Вода накапливается в крупных тонкостенных паренхимных клетках без хлоропластов.
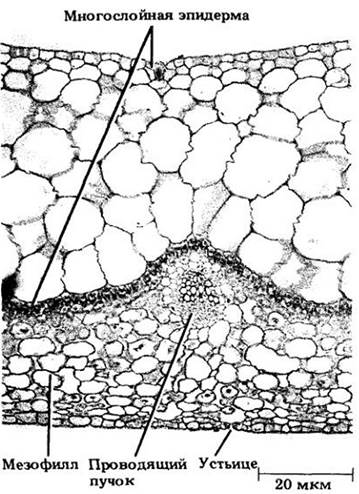
У агавы суккулентные части — листья. В них, как и в суккулентных стеблях, водозапасающая ткань представлена нефотосинтезирующими паренхимными клетками. Другие примеры растений с суккулентными листьями — хрустальная травка (Mesembryanthemum crystallinum), очиток (Sedum) и некоторые виды Peperomia. У хрустальной травки водозапасающую функцию выполняют крупные эпидермальные клетки с придатками (трихомами), называемые водяными пузырьками и удивительно похожие на хрустальные бусины (см. рис. 20-22, В). Водозапасающие клетки листа Ререromia являются частью многослойной эпидермы (рис. 22-43).
Рис. 22-43. Поперечный срез листовой пластинки Peperomia. Очень толстая многослойная эпидерма на верхней стороне листа, предположительно, является водозапасающей тканью
Заключение
Вегетативные апексы побегов большинства цветковых растений организованы по типу туники—корпуса, где туника включает один или более периферических слоев клеток, а корпус представлен массой ткани под ней. Хотя первичные ткани стебля проходят те же стадии развития, что и в корне, стебель нельзя аналогичным образом разбить на зоны деления, растяжения и дифференцировки (созревания). Он растет в длину главным образом за счет удлинения междоузлий.
Как и в корне, апикальная меристема побега дает начало протодерме, основной меристеме и прокамбию, развивающимся в первичные ткани. Существуют три типа распределения здесь основной и первичной проводящей тканей: (1) в виде более или менее непрерывного полого цилиндра, (2) в виде цилиндра из отдельных тяжей и (3) в виде системы тяжей, рассеянных по всей основной ткани. Независимо от типа строения флоэма обычно располагается снаружи от ксилемы.
У большинства двудольных листья состоят из пластинки и черешка. Иногда пластинки разделены на листочки. Устьица обычно более многочисленны на нижней стороне листа. Основная ткань, или мезофилл, листа является специализированной фотосинтезирующей и у мезофитов дифференцирована на палисадную и губчатую паренхиму. Она обильно пронизана межклетниками и жилками, т. е. проводящими пучками, состоящими из флоэмы и ксилемы и окруженными паренхиматозными обкладками. Ксилема обычно расположена на верхней стороне жилки, а флоэма на нижней.
У большинства однодольных, включая злаки, лист состоит из пластинки и влагалища, охватывающего стебель. Листья С3- и С4-злаков имеют существенные анатомические различия. Наиболее значительным из них является присутствие у С4- и отсутствие у Сз-видов кранц-анатомии, т. е. такого строения, при котором клетки мезофилла и обкладки располагаются вокруг проводящих пучков двумя концентрическими слоями.
Листья образуются в периферической зоне апекса побега, и их положение на стебле отражается в особенностях расположения в нем проводящей системы. Их рост детерминированный, т. е. относительно непродолжителен, тогда как у вегетативных апексов побега он может быть неограниченным, или недетерминированным. У многих видов листья, выросшие при высокой освещенности, мельче и толще развившихся при относительном затенении. Первые называются световыми, вторые — теневыми.
У многих растений опадению листьев предшествует формирование у основания черешка отделительной зоны.
Изменение типа структуры, свойственного корню, на характерный для побега происходит в определенной области оси зародыша и молодого проростка, называемой зоной перехода.
При цветении вегетативный апекс непосредственно превращается в репродуктивный.
Побеги, как и корни, могут запасать питательные вещества. Примеры специально приспособленных для этого мясистых побегов — клубни, луковицы и клубнелуковицы. Растения, запасающие воду, называются суккулентами. Водозапасающая ткань их состоит из крупных паренхимных клеток. Суккулентными могут быть стебли, листья или и те, и другие вместе.
Приложение 1. Растения, загрязнение воздуха и кислотные дожди
Лист растения, подобно легкому человека, может функционировать только тогда, когда способен к газообмену с окружающим воздухом. Следовательно, он, как и легкое, чрезвычайно чувствителен к атмосферному загрязнению.
Это загрязнение выступает в разных формах. Иногда оно создается твердыми частицами. Они могут быть органическими (например, в дыме, образующемся при сгорании ископаемого топлива и мусора) или неорганическими (в частности, пыль цементных и металлургических предприятий, соединения свинца, выделяющиеся при сжигании этилированного бензина). Будучи главным компонентом «смога», эти частицы снижают количество солнечного света, достигающего земной поверхности, а также оказывают прямое повреждающее воздействие на растения. Они могут засорять устьица, препятствуя их функционированию, или (особенно металлические) выступать в качестве ядов.
Фтористые соединения, попадающие в воздух в виде отходов фосфатного, сталелитейного, алюминиевого и других производств, действуют как кумулятивные яды, проникая в лист через устьица и разрушая ткани листа, по-видимому, за счет ингибирования ферментов, участвующих в синтезе целлюлозы. Тысячи гектаров цитрусовых рощ Флориды пострадали от выбросов фторидов с предприятий фосфатных удобрений.
При обработке руды, содержащей серу, образуется сернистый газ:
2СuS + 3O2 —> 2СuO + 2SO2.
Это соединение имеет неприятный остросладкий вкус (необычным его свойством как загрязнителя воздуха является то, что вкус ощущается при меньших концентрациях в атмосфере, чем запах). Оксиды серы образуются также при сгорании ископаемого топлива, содержащего этот элемент. Во влажном воздухе они реагируют с водой, образуя капельки серной кислоты, обладающей сильным коррозионным действием и входящей в состав кислотного дождя. В некоторых районах США местность буквально превратилась в пустыню под влиянием выбросов в атмосферу SO2 и металлов. Еще в 1905 г. была начата борьба с загрязнением воздуха в окрестностях медеплавильных предприятий в шт. Теннесси, но и сейчас роскошные леса, когда-то распространенные здесь, полностью отсутствуют. Погибла вся растительность, а питательные вещества из почвы вымыты кислотой. Вокруг медеплавильных заводов долины Сакраменто в Калифорнии растительность уничтожена на площади 260 км2, а еще на 320 км2 ее рост сильно подавлен.
Наиболее распространенная форма атмосферного загрязнения в Калифорнии — фотохимический смог, образующийся при действии солнечного света на автомобильные выхлопы. Район Лос-Анджелеса — «идеальное» место для его возникновения, так как жизнь здесь тесно связана с использованием автомобиля, а горы к северу и востоку образуют барьер, препятствующий циркуляции воздуха. Многие виды растений не могут выживать не только в самом городе (это типично и для многих других крупных городов), но и на расстоянии 160 км от него, где смог, распространяющийся из этого бассейна, повреждает сельскохозяйственные культуры и губит в горах сосновые леса.
A. Повреждение листа ежевики (Rubus) сернистым газом; пораженные участки окружены здоровой тканью. Б. Повреждение озоном листа табака (Nicotiana tabacum) проявляется в виде штрихов или крапин отмершей ткани на его верхней поверхности. При тяжелом поражении озоном такие крапины сливаются в более обширные пятна, заметные на обеих поверхностях. В. Когда оксиды серы и азота реагируют в атмосфере с водой, образуются серная и азотная кислоты, выпадающие на землю в составе кислотных дождей

Один из главных компонентов фотохимического смога — двуокись азота (NO2), возникающая при любых процессах горения в воздухе (в сухом виде он содержит 77% азота), поэтому присутствующая и в автомобильных выхлопах. Под влиянием света NO2 расщепляется на NO и атомарный кислород. Последний чрезвычайно реакционноспособен и образует с молекулярным кислородом озон.
Сходные реакции под влиянием ультрафиолетового излучения происходят в верхних слоях атмосферы, образуя озоновый экран, описанный на с. 13, т. 1. Озон может также возникать при электрических разрядах, давая «свежий» запах после грозы. Это вещество высокотоксично. У растений оно повреждает тонкостенные палисадные клетки, изменяя, по- видимому, проницаемость мембран — клеток и хлоропластов. Другой компонент фотохимического смога — ПАН (пероксиацетилнитрат — С2Н3O5). Он в несколько раз токсичнее озона, но обычно присутствует в воздухе в гораздо меньших концентрациях. Наличие в атмосфере 0,25 млн-1 фотохимического смога снижает фотосинтез на 66%.
В настоящее время большую озабоченность вызывает вредное влияние на окружающую среду кислотного дождя. «Нормальный» дождь, выпадающий в незагрязненных районах, обычно имеет pH 5,6. Сжигание ископаемого топлива и обработка сульфидных руд приводит к выделению в атмосферу больших количеств оксидов серы и азота, которые, реагируя здесь с водой, образуют сильные кислоты (серную и азотную). Дождь и снег в этих условиях имеют pH меньше
5,6, т. е. по определению являются кислотными. Такие осадки сейчас широко распространены, особенно в Западной Европе, на востоке США и юго-востоке Канады, где среднегодовое значение их pH составляет А — 4,5. Кроме того, отдельные ливни бывают здесь гораздо более кислотными. В Шотландии, Норвегии и Исландии зарегистрированы дожди с pH 2,4, 2,7 и 3,5 соответственно. Попытка уменьшить локальное загрязнение путем увеличения высоты труб промышленных предприятий создала региональные проблемы. Загрязнители, выбрасываемые высокими трубами, распространяются по воздуху на большие расстояния. Например, считается, что более 75% серы в дожде, выпадающем в Скандинавии, образуется на Британских островах и в Центральной Европе. Влияние кислотного дождя на растения понятно не до конца, но исследуется повсеместно в Северном полушарии. Показано, что он замедляет рост лесных пород в Швеции. Экспериментальный кислотный дождь повреждает листья и тормозит прорастание семян. В последние несколько лет площадь пострадавших лесов в ФРГ увеличилась с нескольких процентов до более 50%.
Особенно заметно влияние кислотного дождя на популяции рыб, фактически погибшие в подкисленных озерах некоторых районов земного шара. Предполагается, что это обусловлено главным образом не прямым понижением pH воды, а повышением концентрации в ней алюминия. Этот металл, составляющий около 5% земной коры, почти нерастворим в нейтральной или щелочной среде и вследствие этого биологически неактивен. Однако в результате кислотных дождей концентрация растворенного алюминия в некоторых озерах может возрасти до уровня, токсичного для рыбы и других водных организмов. Растворимость других токсичных металлов, в частности свинца, кадмия и ртути, также резко увеличивается с понижением pH.
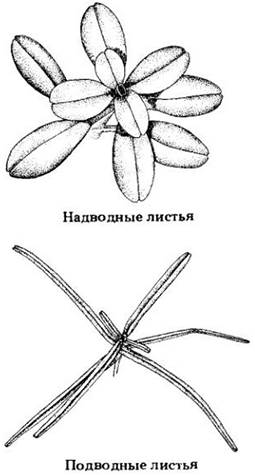
Приложение 2. Диморфизм листьев у водных растений
В естественной среде у водных цветковых растений иногда развиваются две разные формы листьев: под водой — узкие и часто сильно рассеченные (подводные), а над се поверхностью — обычного облика (надводные). Различными обработками можно стимулировать незавершившие формирование листья развиваться в атипичную для данных условий существования форму.
При недавних исследованиях водного растения Callitriche heterophylla было обнаружено, что растительный гормон, гибберелловая кислота, побуждает его воздушные, т. е. выступающие из воды, побеги образовывать подводные листья. Другой растительный гормон, абсцизовая кислота (см. гл. 24), приводит к возникновению надводных листьев на погруженных побегах. К такому же результату приводят повышенные температуры или добавки к воде спирта маннитола.
В природе клеточное тургорное давление погруженных (подводных) листьев относительно высокое, а у надводных относительно низкое. Последнее может отчасти объясняться транспирационной потерей воды через многочисленные устьица на листовой поверхности. С высоким тургором в развивающихся подводных листьях связано появление у них в зрелом состоянии длинных эпидермальных клеток.
Гибберелловая кислота заставляет клетки надводных листьев вытягиваться, увеличивая поглощение воды, а, следовательно, и тургорное давление. Полностью сформировавшись, такие листья обладают всеми признаками типично подводных, включая длинные эпидермальные клетки. Ограниченное растяжение клеток погруженных побегов под воздействием абсцизовой кислоты или высокой температуры, по-видимому, не является результатом снижения тургора; обработанные клетки становятся менее растяжимыми, поэтому высокий тургор не способствует увеличению их размеров. Выращивание погруженных побегов в растворе маннитола приводит к тургорному давлению, сходному с отмечаемым у контрольных надводных побегов, и формированию листьев с короткими эпидермальными клетками.
Результаты этих экспериментов показывают, что относительная величина клеточного тургорного давления определяет окончательные размеры и форму листа у Callitriche heterophylla.Таким образом, простое присутствие или отсутствие вокруг воды во время его развития приводит к соответствующей адаптации к над- или подводным условиям.
Приложение 3. Конвергентная эволюция
Сходные факторы отбора, действующие на растения в близких условиях, но в разных частях света, часто способствуют тому, что совсем неродственные виды приобретают сходный облик. Процесс, приводящий к этому, называют конвергентной эволюцией.
Рассмотрим некоторые адаптивные признаки пустынных растений: мясистые колоннообразные стебли (обеспечивающие запасание воды), защитные иглы и редуцированные листья. В трех весьма далеких друг от друга семействах цветковых — молочайных (Еuphorbiасеае), кактусовых (Саctaсеае) и ластовневых (Аsclepiadасеае) — известны представители со всеми этими признаками. Кактусоподобные молочайные и ластовневые, показанные на рисунках, произошли от облиственных растений, совершенно непохожих друг на друга.
Естественный ареал кактусов (за единственным исключением) охватывает лишь Новый Свет. Сравнительно мясистые представители двух других названных семейств встречаются главным образом в пустынных областях Азии и особенно Африки, где занимают такую же экологическую нишу, что и кактусы в Америке.
Хотя растения, показанные здесь (А — Euphorbia из молочайных; Б — кактус Echinocereus; В — Hoodia, суккулент из ластовневых), имеют фотосинтез типа САМ, все они родственны С3-растениям и происходят от них. Отсюда можно сделать вывод, что физиологические адаптации, связанные с CAM-фотосинтезом, также возникли в результате конвергентной эволюции (см. гл. 7).

Проводящие пучки, более или менее полностью погруженные в мезофилл, называют мелкими жилками, а сопровождающиеся выступами на нижней стороне листа — крупными жилками. Главную роль в отводе ассимилятов из клеток мезофилла играют первые. С увеличением размеров жилок их связь с мезофиллом ослабевает, и они все больше окружаются нефотосинтезирующими тканями гребней. Следовательно, функция сбора ассимилятов постепенно вытесняется главным образом транспортной.
Рис. 22-16. Сидячие (бесчерешковые) листья часто встречаются у двудольных, например, у Moricandia из семейства крестоцветных (А), но особенно характерны для злаков и других однодольных. Б. У кукурузы (однодольного растения) основание листа образует вокруг стебля влагалище, из которого выступает язычок — небольшой вырост листовой ткани
