СОВРЕМЕННАЯ БОТАНИКА - П. РЕЙВН - 1990
РАЗДЕЛ V. СТРОЕНИЕ И РАЗВИТИЕ ТЕЛА ПОКРЫТОСЕМЕННЫХ
ГЛАВА 23. ВТОРИЧНЫЙ РОСТ
У многих растений (например, у большинства однодольных и некоторых травянистых двудольных, в частности, у лютика) с завершением дифференцировки первичных тканей рост данной части тела прекращается. Другой крайний случай представляют собой голосеменные и древесные двудольные, корни и стебли которых продолжают увеличиваться в диаметре даже в участках, завершивших удлинение (рис. 23-1). Это нарастание в толщину, или в обхват, называемое вторичным ростом, — результат деятельности двух латеральных меристем — камбия и пробкового каvбия.
Рис. 23-1. Одиноко растущее дерево карии овальной (Сагуa ovata) зимой. Растения могут достигать таких крупных размеров благодаря способности корней и стеблей разрастаться в поперечнике, т. е. их вторичному росту. Большая часть образующихся при этом тканей представлена вторичной ксилемой, или древесиной, не только проводящей воду и минеральные вещества, но и придающей большую прочность корням и стеблям

У трав, т. е. травянистых растений, вторичный рост побегов слабый или вообще отсутствует. В умеренных зонах такой побег или все растение в зависимости от вида живет в течение только одного сезона. Древесные растения, т. е. деревья и кустарники, — многолетние. В начале каждого вегетационного периода возобновляется их первичный рост, а к более старым частям тела латеральные меристемы, восстанавливая свою активность, добавляют новые ткани. Хотя у большинства однодольных вторичный рост отсутствует, некоторые из них (например, пальмы) образуют толстые стебли за счет одного первичного роста (см. с. 45).
Растения часто подразделяют в соответствии с сезонными циклами роста на одно-, дву- и многолетники. У однолетников, к которым относятся многие сорняки, дикорастущие и садовые «цветы» и овощи, все развитие от семени через вегетативную фазу до цветения и нового образования семян завершается в течение одного вегетационного периода продолжительностью иногда всего несколько недель. Между этими периодами растение представлено покоящимся семенем.
Двулетники развиваются от прорастания семени до формирования новых семян в течение двух вегетационных периодов. Первый из них завершается образованием корня, короткого стебля и розетки листьев у поверхности почвы. В течение второго растение цветет, плодоносит, дает семена и, закончив жизненный цикл, отмирает. В умеренных зонах однолетники и двулетники редко одревесневают, хотя у их стеблей и корней может наблюдаться ограниченный вторичный рост.
Многолетники — это растения с вегетативными частями, живущими и нарастающими в течение многих лет. Травянистые многолетники переживают неблагоприятные периоды в виде покоящихся подземных органов — корней, корневищ, луковиц, клубней. Древесные многолетники, к которым относятся лианы, кустарники и деревья, сохраняют в таких условиях надземные части, но обычно прекращают рост. Они цветут, только достигнув зрелого состояния, для чего может потребоваться много лет. Например, конский каштан (Aesculus hippocastanum) не зацветает до 25-летнего возраста, a Puya raimondii, очень крупное, родственное ананасу растение (семейство бромелиевых) из Анд высотой до 10 м — до 150-летнего. Многие древесные виды листопадные, т. е. теряют все листья одновременно и развивают их из почек при новом наступлении благоприятного сезона. У вечнозеленых деревьев и кустарников листья также опадают и сменяются, но не все сразу.
Камбий
В отличие от многогранных инициалей апикальных меристем, содержащих плотную цитоплазму и крупные ядра, меристематические клетки камбия сильно вакуолизированы. Известны две их формы: удлиненные в вертикальном направлении веретеновидные инициали и вытянутые в горизонтальном направлении или отчасти квадратные лучевые инициали. Длина первых намного превышает ширину, и на поперечном срезе они уплощены наподобие кирпичей. У веймутовой сосны (Pinus strobus) их длина в среднем 3,2 мм, у яблони (Malus sylvestris) — 0,53 мм (рис. 23-2), а у белой акации (Robinia pseudo-acacia) — 0,17 мм (рис. 23-3).
Рис. 23-2. Танге шпальный срез камбия яблони (Malus sylvestris). Такие срезы делаются под прямыми углами к лунам, которые видны ни них в поперечном сечении. Камбий, подобный изображенному здесь, с веретеновидными инициалями, не расположенными на тангентальных срезах горизонтальными рядами, называют неярусным

Рис. 23-3. Тангентальный срез камбия белой акации (Robinia pseudo-acacia). В этом камбии веретеновидные инициали расположены на тангентальных срезах горизонтальными рядами. Его называют ярусным

Вторичные ксилема и флоэма формируются в результате периклинальных делений камбиальных инициалей и их производных. Другими словами, возникающие между ними клеточные пластинки проходят параллельно поверхности корня или стебля (рис. 23-4, А). При делении камбиальной инициали клетка, откладывающаяся в сторону этой поверхности, со временем становится флоэмной, а оказывающаяся ближе к сердцевине — ксилемной. Таким образом возникает длинный непрерывный ряд клеток, тянущийся радиально от камбия — наружу к флоэме и внутрь к ксилеме (рис. 23-5).
Рис. 23-4. Периклинальные и антиклинальные деления веретеновидных инициален. А. Периклинальные деления дают клетки вторичных ксилемы и флоэмы, расположенные радиальными рядами (см. рис. 23-5). При таких делениях одна из дочерних клеток возникает позади (или впереди) другой. Б. Антиклинальные деления увеличивают число веретеновидных инициален. При этом на месте одной клетки появляются две расположенные бок о бок дочерние
Рис. 23-5. Схема связи камбия с происходящими из него тканями — вторичными ксилемой и флоэмой. Камбий состоит из клеток двух типов, веретеновидных и лучевых инициалей, дающих начало соответственно осевой и радиальной системам проводящих тканей. Делясь периклинально, камбиальные инициали образуют вторичные ксилему и флоэму. После такого деления одна дочерняя клетка (инициале) остается меристематической, а другая (производная инициали) со временем развивается в одну или несколько клеток проводящей ткани: с внутренней поверхности камбия — в ксилемные элементы, а с наружной — во флоэмные. Лучевые инициали, делясь, образуют лучи, проходящие перпендикулярно к производным веретеновидных инициалей. Прирост вторичной ксилемы смещает камбий и вторичную флоэму наружу. Схемы (слева направо) показывают последовательные стадии созревания
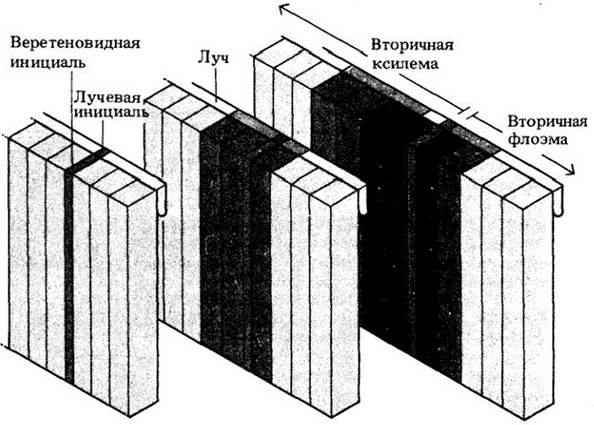
Образованные веретеновидными инициалями клетки ксилемы и флоэмы с их вертикально ориентированными продольными осями составляют осевую систему вторичных проводящих тканей. Лучевые инициали дают ориентированные горизонтально лучевые клетки радиальных лучей, или радиальной системы (рис. 23-5). Эти лучи состоят главным образом из паренхимных клеток и не одинаковы по длине. Питательные вещества переносятся от протопласта к протопласту по плазмодесмам (симпластный транспорт), проходя от вторичной флоэмы через камбий и далее по радиальным лучам к живым клеткам вторичной ксилемы. В то же время вода проходит от вторичной ксилемы к камбию и вторичной флоэме главным образом по оболочкам (апопластный транспорт) клеток лучей и осевой системы. Лучи служат также местом запасания крахмала и липидов.
В узком смысле термин «камбий» относится только к камбиальным инициалям, которых в каждом радиальном ряду всего по одной. Однако часто трудно или вообще невозможно отличить эти клетки от их непосредственных производных, иногда в течение длительного времени остающихся меристематическими (рис. 23-5). Даже зимой, когда камбий неактивен, между ксилемой и флоэмой можно видеть несколько слоев сходных по виду недифференцированных клеток. Поэтому некоторые ботаники применяют термин «камбий» в широком смысле для обозначения инициалей и их ближайших производных. Другие называют соответствующую им область камбиальной зоной.
По мере откладывания камбием клеток вторичной ксилемы и нарастания ее в толщину, сам он двигается наружу, параллельно компенсируя это увеличение своей окружности за счет антиклинальных делений инициалей (рис. 23-4, Б). При этом число веретеновидных и лучевых инициалей (и новых лучей) увеличивается так, что соотношение между ними во вторичных проводящих тканях поддерживается примерно на одном уровне. Очевидно, что изменения, происходящие в камбии при его развитии, чрезвычайно сложны.
В умеренных зонах камбий зимой находится в состоянии покоя, а весной возобновляет свою деятельность. За вегетационный период откладываются новые слои вторичных флоэмы и ксилемы. Сигналом к реактивации камбия служит распускание почек и возобновление их роста. По-видимому, она стимулируется гормоном ауксином, образующимся в развивающихся побегах и передвигающимся вниз по стеблю. Реактивации и поддержанию деятельности камбия способствуют и другие факторы (гл. 24).
Влияние вторичного роста на первичное тело растения
Корень
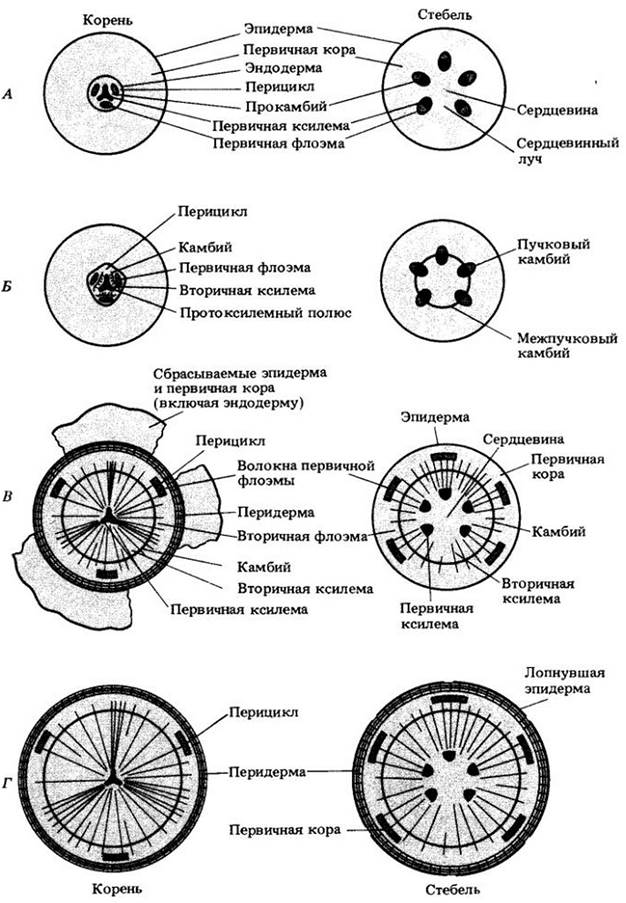
В корнях камбий возникает из меристематических прокамбиальных клеток, остающихся между первичными ксилемой и флоэмой. В зависимости от числа флоэмных тяжей более или менее одновременно закладываются две или более зоны камбиальной активности (рис. 23-6). Вскоре после этого клетки перицикла против протоксилемных полюсов делятся периклинально, и к камбию прибавляются образующиеся при этом внутренние сестринские клетки. Теперь он полностью окружает ксилему.
Против флоэмных тяжей камбий сразу же начинает формировать вторичную ксилему, в результате чего эти тяжи смещаются со своих мест между гребнями ксилемы наружу. Ко времени активного деления камбия против протоксилемных полюсов весь он уже располагается по окружности, и первичная флоэма оказывается отделенной от первичной ксилемы (рис. 23-6).
Рис. 23-6. Сравнение первичной и вторичной структур корня и стебля древесных двудольных. А. Корень и стебель в конце первичного роста. В представленном здесь триархном корне камбий закладывается в трех независимых зонах прокамбия, между тремя тяжами первичной флоэмы и первичной ксилемой. Б. Происхождение камбия. Клетки перицикла против трех протоксилемных полюсов также участвуют в его формировании. Немного вторичной ксилемы уже образовалось за счет камбия прокамбиального происхождения. В. В корне и стебле присутствуют вторичные ксилема и флоэма, а в корне также перидерма. Г. Результат изменения первичного тела растения под влиянием вторичного роста (включая образование перидермы) к концу первого вегетационного периода. Радиальные линии на рис. В и Г соответствуют лучам
В ходе неоднократных делений внутрь и наружу от камбия откладываются вторичные ксилема и флоэма корня (рис. 23-6 и 23-7). В некоторых случаях камбий, закладывающийся в пери цикле, образует широкие лучи, в то время как в других частях вторичных проводящих тканей возникающие лучи более узкие.
Рис. 23-7. Поперечные срезы одревесневающего корня ивы (Salix). А. Стадия, близкая к завершению первичного роста. К. Фрагмент первичного центрального цилиндра. В. Конец первого вегетационного периода; можно видеть влияние вторичного роста на первичное тело растения
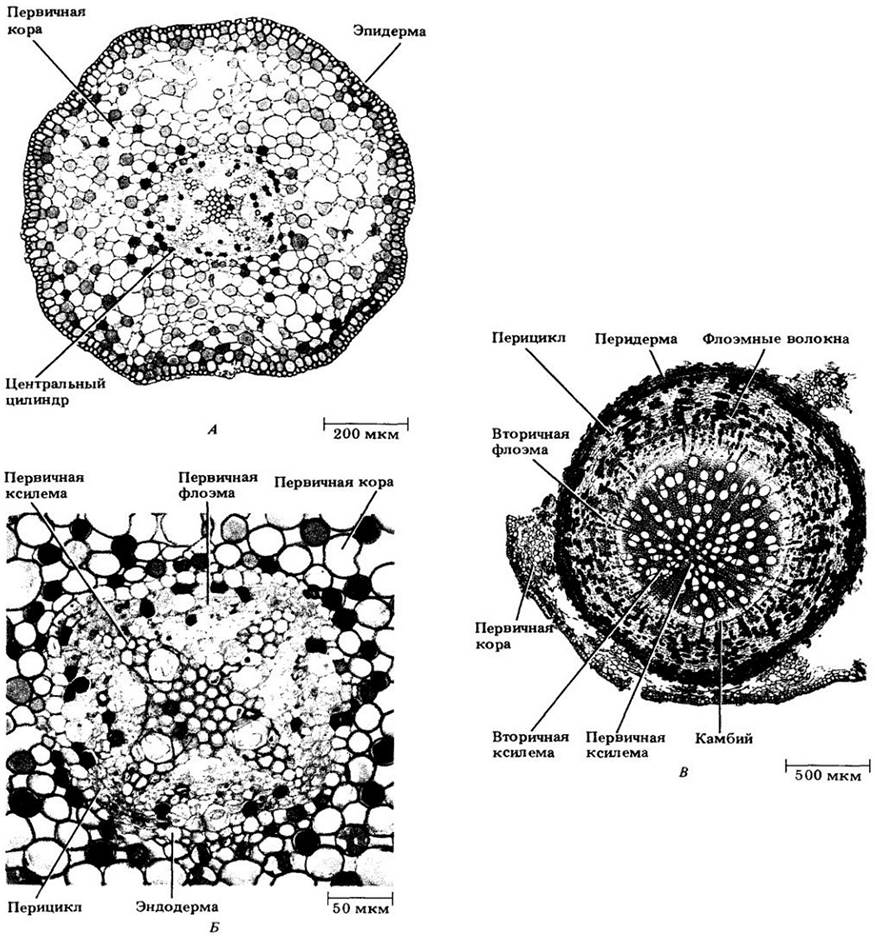
С разрастанием вторичных ксилемы и флоэмы в ширину большая часть первичной флоэмы сдавливается, или облитерируется. Иногда от нее остаются заметными только волокна.
Стебель
Как упоминалось ранее, камбий стебля возникает из недифференцированного прокамбия между первичными ксилемой и флоэмой, а также из паренхимы межпучковых зон. Ту его часть, которая закладывается в проводящих пучках, называют пучковым камбием, а ту, что образуется в межпучковых зонах, или сердцевинных лучах, — межпучковым камбием. В стебле в отличие от корня он образует кольцо непосредственно с момента возникновения (см. рис. 23-6).
В деревянистых стеблях вторичные ксилема и флоэма образуют цилиндр из проводящих тканей, пересекающийся в радиальном направлении лучами (рис. 23-6). Обычно ежегодно откладывается гораздо больше вторичной ксилемы, чем вторичной флоэмы (это же происходит и в корне). Как и в корне, по мере вторичного роста первичная флоэма, сдвигается наружу и ее тонкостенные клетки облитерируются, так что от этой ткани сохраняются только толстостенные волокна (рис. 23-9).
На рис. 23-8 и 23-9 показаны две стадии вторичного роста стебля бузины (Sarnbucus canadensis) (см. описание его первичного строения на с. 46 и 49). На первой из них (рис. 23-8) образовалось лишь небольшое количество вторичных ксилемы и флоэмы. Стебель к концу первого вегетационного периода изображен на рис. 23-9. Можно видеть, что вторичной ксилемы отложено значительно больше, чем вторичной флоэмы. Толстостенные клетки снаружи от последней представляют собой волокна первичной флоэмы.
Рис. 23-8. Поперечный срез стебля бузины (Sambucus canadensis), начавшего вторичный рост. Пробковый камбий еще не сформировался
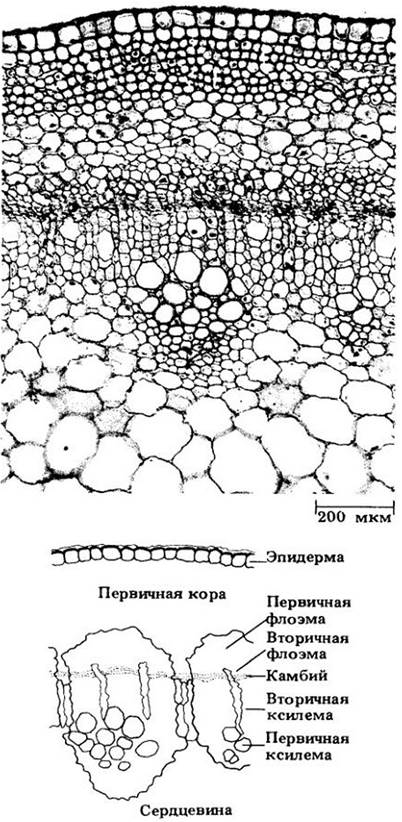
Рис. 23-9. Поперечный срез стебля бузины (Sambucus canadensis) в конце первого вегетационного периода
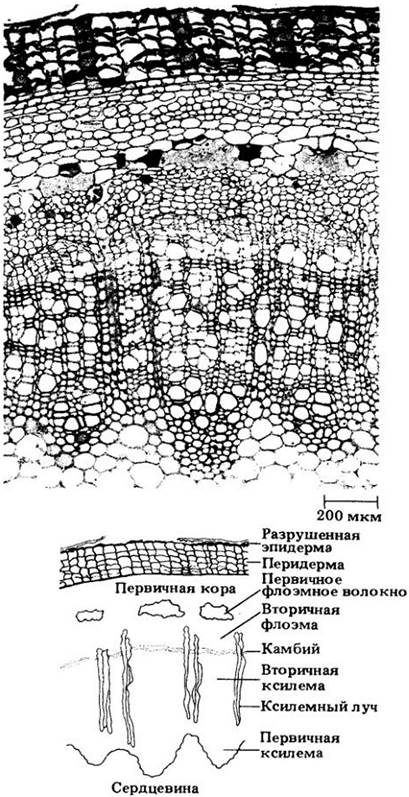
На рис. 23-10 представлено строение одно-, дву- и трехлетнего стебля липы (Tilia americana). В гл. 22 говорилось, что первичные ткани образуют в нем почти непрерывный полый цилиндр. Таким образом, большая часть камбия здесь пучковая по происхождению. Некоторые лучи во вторичной флоэме липы при разрастании стебля в толщину становятся очень широкими. Это один из способов сохранить ткани, расположенные снаружи от камбия, при увеличении диаметра ксилемы.
Рис. 23-10. Поперечные срезы стебля липы (Tilia americana). А. Однолетний стебель. Б. Двулетний стебель. В. Трехлетний стебель. Числа означают номера годичных колец вторичной ксилемы
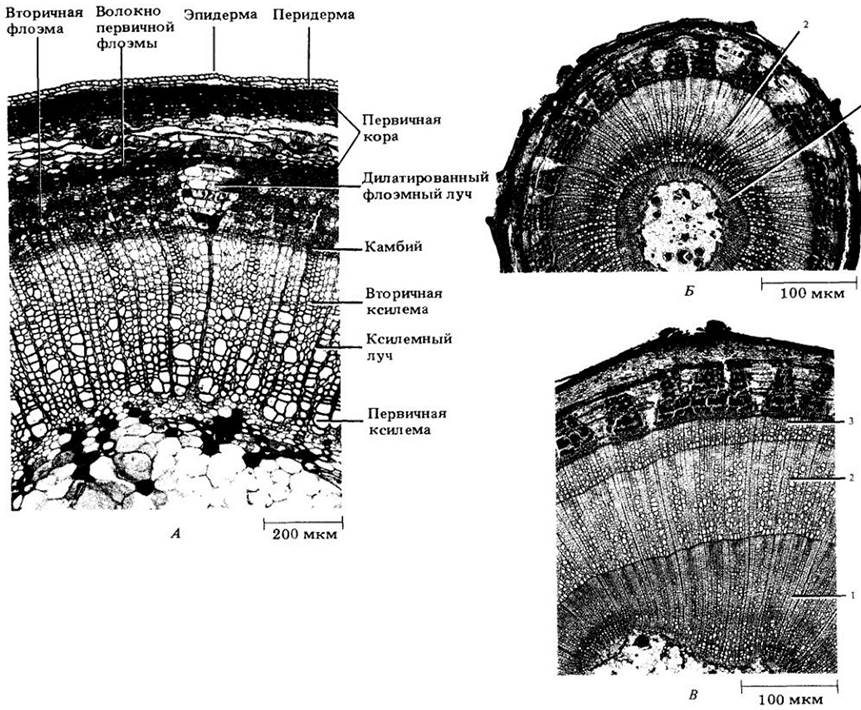
Камбий и вторичные ткани корня и стебля без перерывов переходят друг в друга. В отличие от первичного тела растения во вторичной зоне перехода нет (см. с. 63).
Перидерма
В большинстве деревянистых корней и стеблей за началом формирования вторичных ксилемы и флоэмы обычно следует образование пробки, и пробковая ткань замещает эпидерму на этих частях растения в качестве защитного покрова. Пробка, или феллема, откладывается пробковым камбием, или феллогеном, который может также давать феллодерму («пробковую кожицу»). Пробка образуется снаружи от него, а феллодерма — изнутри (рис. 23-11 и 23-12). Вместе эти три ткани — пробка, феллоген и феллодерма — составляют перидерму.
Рис. 23-11. Несколько стадий развития перидермы и чечевички у бузины (Sambucus canadensis) на поперечных срезах. А. Только что сформированная перидерма под эпидермой; колленхима и паренхима первичной коры. Б. Перидерма на более продвинутой стадии развития. В. Начало формирования чечевички; под ней колленхима первичной коры. Г. Хорошо развитая чечевичка. Феллодерма бузины обычно состоит из одного слоя клеток

У большинства двудольных и голосеменных первая перидерма обычно появляется в течение первого вегетационного периода в тех частях корня или стебля, которые прекратили рост в длину. В стеблях первый пробковый камбий чаще всего закладывается в слое коры, находящемся непосредственно под эпидермой (рис. 23-6 и 23-11), хотя у многих видов и непосредственное эпидерме. В корнях первый феллоген образуется за счет периклинальных делений клеток перицикла, причем наружные сестринские клетки смыкаются в сплошной цилиндр пробкового камбия. После этого оставшиеся клетки перицикла могут делиться под перидермой, давая начало ткани, сходной с первичной корой (см. рис. 23-6 и 23-7).
Многократные деления пробкового камбия приводят к формированию радиальных рядов плотно расположенных в большинстве случаев пробковых клеток (рис. 23-11 и 23-12). В процессе дифференцировки этих клеток их внутренние стенки выстилаются толстым слоем жироподобного вещества — суберина, сильно снижающего проницаемость ткани для воды и газов. Их стенки могут также лигнифицироваться. В зрелом состоянии пробковые клетки мертвые.
Клетки феллодермы остаются живыми, не суберинизируются и напоминают паренхимные клетки первичной коры. Они могут отличаться от них своим внутренним положением в общих для всех клеток перидермы радиальных рядах (рис. 23-12).
Рис. 23-12. Чечевичка стебля кирказона (Aristolochia) на поперечном срезе. В отличие от бузины феллодерма здесь состоит из нескольких слоев клеток
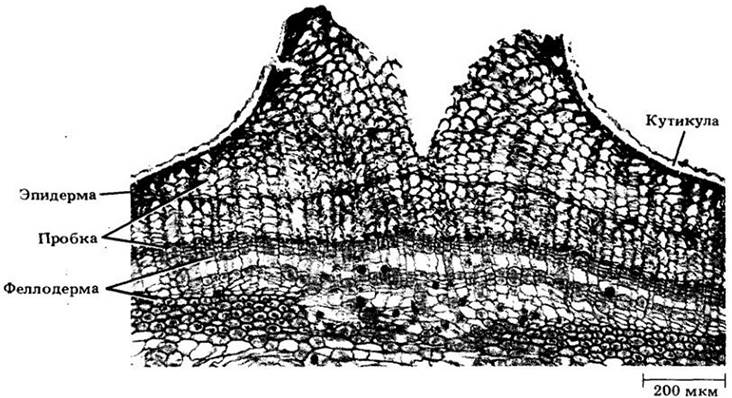
С образованием первой перидермы корня первичная кора (включая эндодерму) и эпидерма изолируются от остальной его части. Подстилаемые непроницаемым для воды и минеральных веществ пробковым слоем, они со временем отмирают и сбрасываются. Поскольку первая перидерма стебля обычно закладывается непосредственно под эпидермой, первичная кора в первый год не сбрасывается (см. рис. 23-6 и 23-9), хотя эпидерма все же высыхает и слущивается.
К концу первого вегетационного периода в деревянистом корне присутствуют следующие ткани (снаружи внутрь): остатки эпидермы и первичной коры, перидерма, перицикл, первичная флоэма (волокна и сдавленные тонкостенные клетки), вторичная флоэма, камбий, вторичная и первичная ксилема. В стебле порядок такой: остатки эпидермы, перидерма, первичная кора, первичная флоэма (волокна и сдавленные тонкостенные клетки), вторичная флоэма, камбий, вторичная и первичная ксилема, сердцевина (см. рис. 23-6).
Чечевички
Выше отмечалось, что суберинсодержащие пробковые клетки образуют плотную ткань, представляющую собой непроницаемый барьер для воды и газов. Однако внутренние части стебля и корня, как и все метаболически активные ткани, нуждаются в газообмене с окружающим воздухом (в последнем случае — с воздушными полостями между частицами почвы). При наличии перидермы этот газообмен осуществляется через чечевички (см. рис. 23-11 и 23-12), т. е. ее участки, в которых более активный, чем в других местах, феллоген образует ткань с многочисленными межклетниками и сам содержит их.
Чечевички начинают формироваться в ходе развития первой перидермы (см. рис. 21-11) и в стебле появляются главным образом под устьицем или группой устьиц. На поверхности стебля или корня они выглядят круглыми, овальными или вытянутыми бугорками (см. рис. 23-18). Чечевички образуются также на некоторых плодах. Они, например, заметны в виде мелких крапин на яблоках и грушах. Когда корни и стебли становятся старше, чечевички продолжают развиваться в новой перидерме на дне возникающих в коре трещин.
Кора
Термины «перидерма», «пробка» и «кора» часто смешивают. Как отмечалось выше, пробка — это одна из трех частей перидермы, вторичная ткань, замещающая эпидерму у большинства одревесневших корней и стеблей. Термин кора означает все ткани снаружи от камбия, включая перидерму, если она присутствует (рис. 23-13 и 23-14). Когда камбий только появляется, а вторичная флоэма еще не сформировалась, кора целиком состоит из первичных тканей. В конце первого вегетационного периода она включает еще существующие первичные ткани, вторичную флоэму, перидерму и все отмершие ткани снаружи от последней.
Каждый вегетационный период за счет деятельности камбия к коре добавляются новые порции вторичной флоэмы, а к центральной части стебля или корня — вторичной ксилемы. Обычно вторичной флоэмы образуется меньше, чем вторичной ксилемы. Кроме того, в ее старых частях тонкостенные клетки (ситовидные и различного рода паренхимные элементы) обычно сдавливаются (см. рис. 23-15 — 23- 17). Со временем старая вторичная флоэма изолируется от более молодой, новыми слоями перидермы. В результате в стебле и корне вторичной флоэмы накапливается значительно меньше, чем вторичной ксилемы, объем которой год от года возрастает.
По мере увеличения диаметра стебля или корня давление на более старые ткани коры возрастает. У некоторых растений в них появляются разрывы и образуются крупные воздушные полости. У многих видов паренхимные клетки осевой системы и лучей делятся и растягиваются, поэтому при увеличении окружности данной части растения старая вторичная флоэма некоторое время не распадается. Выше отмечалось, что некоторые лучи в стволе липы при его утолщении сильно расширяются. Их называют дилатированными.
При нарастании стебля или корня в толщину первая перидерма может сохраняться в течение нескольких лет, причем периодическая активность пробкового камбия чередуется с неактивными периодами (синхронно или не синхронно с работой камбия). В стволах яблони (Malus sylvestris) и груши (Pyrus communis) первый пробковый камбий может оставаться активным до 20 лет. В большинстве деревянистых корней и стеблей по мере увеличения поперечника осевых систем все глубже и глубже в коре возникают добавочные перидермы (см. рис. 23-13 и 23-14) из паренхимных клеток флоэмы, выключающихся из активного транспорта питательных веществ. Эти клетки становятся меристематическими и дают новые пробковые камбии.
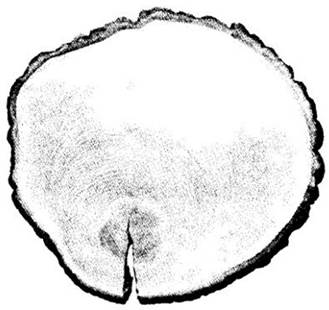
Рис. 23-13. Схема части ствола дуба красного (Quercus rubra) с поперечной, тангентальной и радиальной поверхностей. Темная зона в центре — ядро. Светлая часть древесины — заболонь
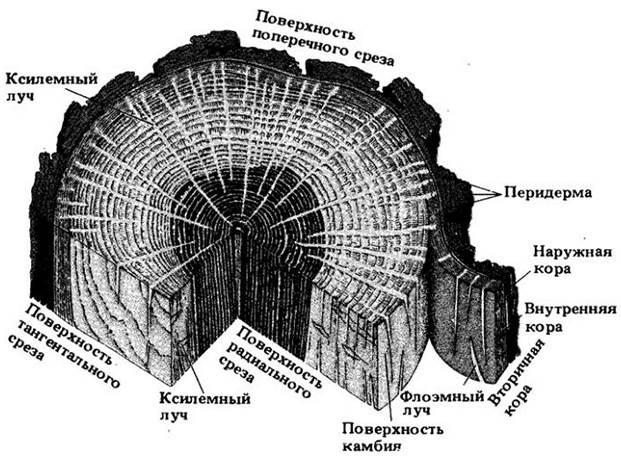
Рис. 23-14. Поперечный срез коры и части вторичной ксилемы старого ствола липы (Tilia americana). Несколько слоев перидермы пересекают в основном буроватую наружную кору в верхней трети среза. Под ней расположена внутренняя кора, хорошо отличающаяся внешне от более светлой ксилемы в нижней трети среза

Все ткани снаружи от самого внутреннего пробкового камбия, т. е. все перидермы с остатками коры и флоэмы между ними, составляют наружную кору (см. рис. 23-13 и 23-14). С созреванием суберинсодержащих пробковых клеток эти ткани перестают получать воду и минеральные вещества. Живая часть коры между камбием и самым внутренним слоем феллогена называется внутренней корой (см. рис. 23-13 и 23-14).
Способ формирования новых перидерм и характер изолируемых ими тканей оказывают существенное влияние на внешний вид коры (см. рис. 23-19). У некоторых растений новые перидермы развиваются прерывистыми, перекрывающими друг друга слоями, в результате чего возникает так называемая чешуйчатая кора (см. рис. 23-13 и 23-14), например, на относительно молодых стволах сосны (Pinus) и груши (Pyrus communis). В других случаях они образуются более или менее непрерывно и концентрически вокруг оси стебля, что приводит к формированию кольцевой коры, например, у винограда (Vitis) и жимолости (Lonicera). Она менее распространена, чем чешуйчатая. У многих растений кора промежуточного типа.
Коммерческую пробку получают из коры пробкового дуба (Quercus súber), родина которого — Средиземноморье. Первый феллоген у этого дерева закладывается в эпидерме, и пробка, формируемая им, имеет небольшую ценность. Когда дуб достигает примерно 20-летнего возраста, ранняя перидерма удаляется и в первичной коре в нескольких миллиметрах под первым закладывается новый пробковый камбий. Образованная им пробка нарастает очень быстро и примерно через 10 лет становится такой толстой, что ее можно снимать с дерева на продажу. Опять несколько глубже предыдущего закладывается новый феллоген, и процедура приблизительно с десятилетним интервалом повторяется, пока дерево не достигнет 150 лет и более. Пятнышки и продолговатые темные штрихи, заметные на поверхности продажной пробки, — чечевички.
У большинства деревянистых корней и стеблей в проведении питательных веществ действительно участвует лишь небольшая часть вторичной флоэмы. Как правило, дальний их транспорт обеспечивает только ее прирост текущего года. Причина этого — недолговечность ситовидных элементов (гл. 20), большинство которых отмирает к концу того же года, в котором они образуются из камбия. У некоторых растений, например, у белой акации (Robinia pseudoacacia), эти элементы спадаются и разрушаются вскоре после отмирания их содержимого (см. рис. 23-15 — 23-17).
Рис. 23-15. Поперечный срез коры стебля белой акации (Robinia pseudo-acacia), состоящей главным образом из нефункциональной флоэмы
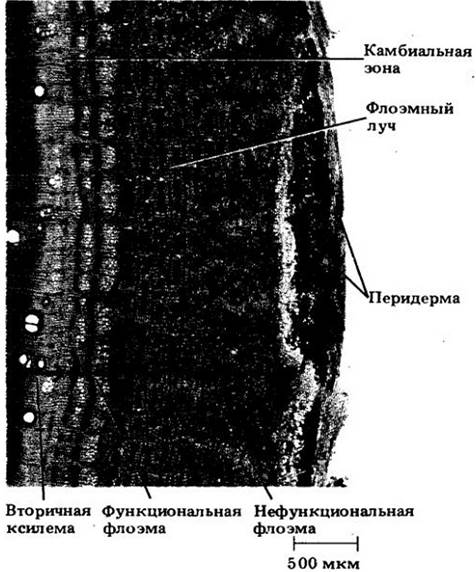
Рис. 23-16. Поперечный срез вторичной флоэмы белой акации; видна главным образом ее функциональная часть. Ситовидные элементы (показаны стрелками) нефункциональной флоэмы сдавлены
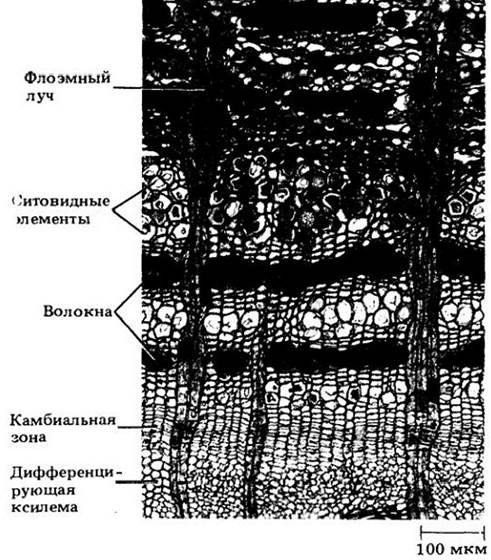
Рис. 23-17. Радиальный срез коры белой акации. Видна в основном нефункциональная флоэма со сдавленными ситовидными элементами (стрелки). У этого дерева функциональна только флоэма, возникшая в текущий вегетационный период. Она становится нефункциональной поздней осенью, когда ситовидные элементы отмирают и сдавливаются
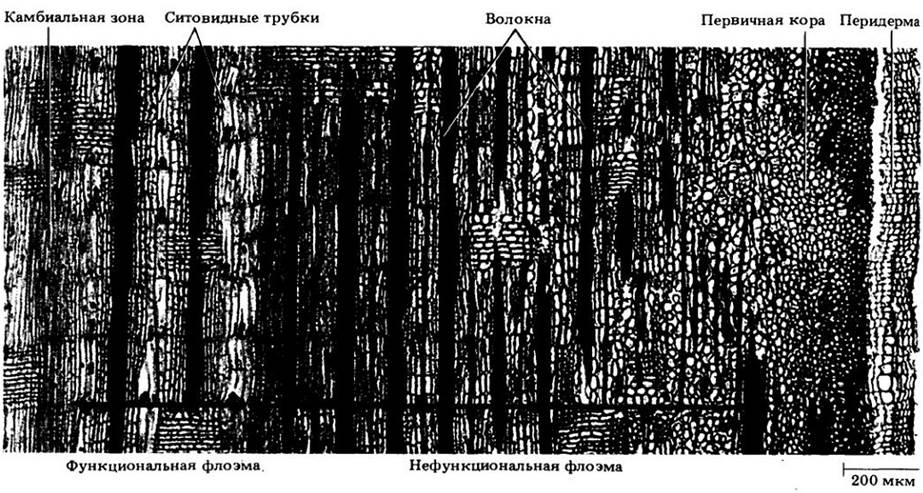
Рис. 23-18. Внешнее строение древесных стеблей. Осмотр ветвей листопадных пород позволяет обнаружить много важных черт строения и развития стебля. Самые заметные структуры — почки, образующиеся на верхушках (терминальные) и в пазухах листьев (боковые, или пазушные). Кроме того, у некоторых видов встречаются придаточные почки — по одной с каждой сто роны пазушной. Иногда при нормальном развитии связанных с ними пазушных почек они не развиваются. У других видов придаточные почки дают начало цветкам, а пазушные — облиственному побегу. После опадения листьев под пазушными почками остаются листовые рубцы со своими пучковыми рубцами. Листовой рубец образуется защитным слоем отделительной зоны. Пучковые рубцы — обломанные концы проводящих пучков, следовавших из листовых следов в черешок листа до его опадения. Группы терминальных рубцов почечных чешуй соответствуют местоположению прежних терминальных почек; пока вторичный рост не сделает их неразличимыми, эти рубцы можно использовать для определения возраста участков стебля. Его часть между двумя группами рубцов соответствует годичному приросту. Чечевички выглядят на стебле мелкими бугорками. А. Ясень пенсильванский (Fraxintcs реnnsylvanica var. subintegerrima). Б. Дуб белый (Quercus alba). В. Липа американская (Tilia americana). Г. Клен ясенелистный (Acer negundo). Д. Ильм американский (Ulmus americana). Е. Конский каштан (Aesculus hippocastanum). Ж. Орех серый (Juglans cinerea). 3. Белая акация (Robinia pseudoacacia)
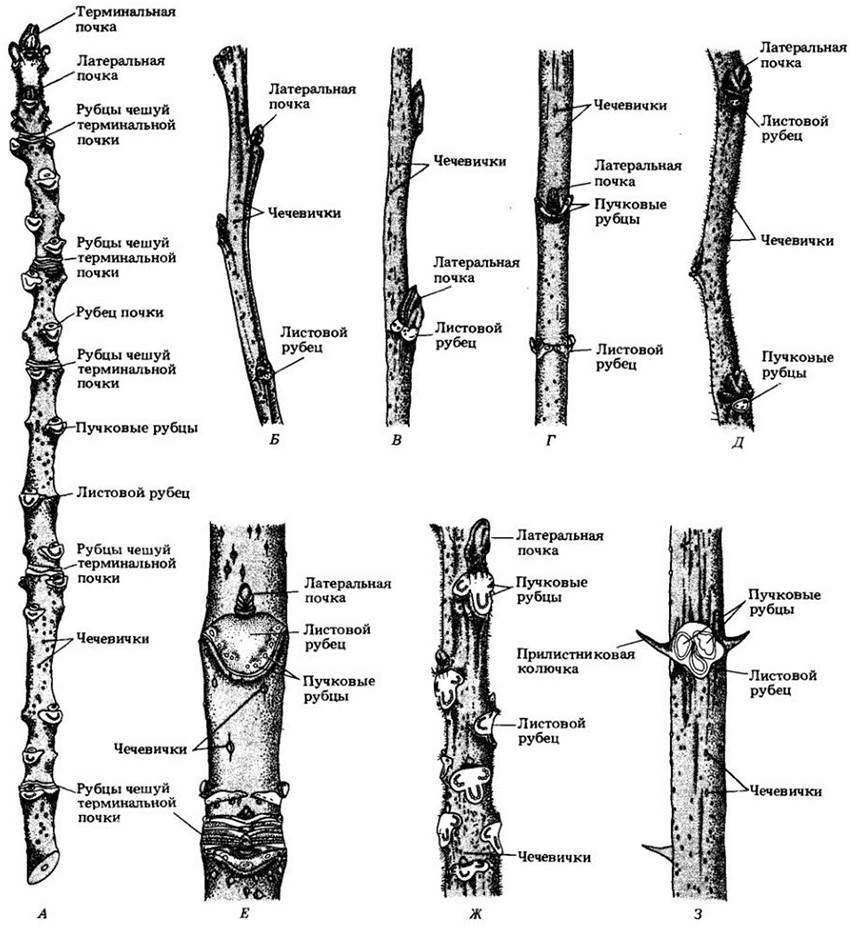
Рис. 23-19. Кора четырех древесных пород. А. Тонкая шелушащаяся кора березы бумажной (Betula papyrifera). Штрихи на ее поверхности — чечевички. Б. Лохматая кора карии овальной (Сагуа ovata). В. Чешуйчатая кора платана западного (Platanus occidentalis). Г. Глубокобороздчатая кора дуба бархатного (Quercus velutina)

Часть внутренней коры, активно проводящая питательные вещества, называется функциональной флоэмой. Хотя ситовидные элементы снаружи от нее мертвые, паренхимные клетки флоэмы и лучей (соответственно осевая и лучевая паренхима) могут оставаться живыми и продолжать функционировать многие годы в качестве запасающих. Эту часть внутренней коры называют нефункциональной флоэмой. Только наружная кора целиком состоит из мертвой ткани (см. рис. 23-15 и 23-17).
Древесина: вторичная ксилема
Если не считать использования различных растительных тканей в пищу, ни одна из них не играла в жизни человека на протяжении всей его истории более существенной роли, чем древесина, или вторичная ксилема (см. табл. 23-1). Обычно различают твердую и мягкую древесину. Первую дают двудольные, а вторую — хвойные. Два типа древесины имеют существенные структурные различия, причем термины «твердая» и «мягкая» не обязательно отражают ее плотность или твердость. Например, одна из самых легких и мягких древесин у бальзы заячьей (Ochroma lagopus), тропического двудольного. И напротив, у некоторых хвойных, например, сосны Эллиота (Pinus elliottii), она тверже, чем у многих лиственных пород.
Древесина хвойных
Структура этой древесины по сравнению с имеющейся у большинства двудольных относительно проста. Главная ее особенность — отсутствие сосудов (гл. 20) и относительно небольшое количество осевой, или древесинной, паренхимы. Преобладают длинные трахеиды с заостренными концами. У некоторых родов, в частности у сосен, паренхимные клетки осевой системы связаны только со смоляными ходами — относительно крупными межклеточными пространствами, выстланными тонкостенными паренхимными клетками, которые секретируют в них смолу. У сосен они встречаются как в осевой системе, так и в лучах (рис. 23-20 и 23-21). Поранение, сжатие, повреждение морозом или ветром могут стимулировать образование смоляных ходов в древесине хвойных, что дало некоторым исследователям повод считать их во всех случаях травматическими по происхождению. Смола, по-видимому, защищает растения от поражения грибами- паразитами и жуками-короедами.
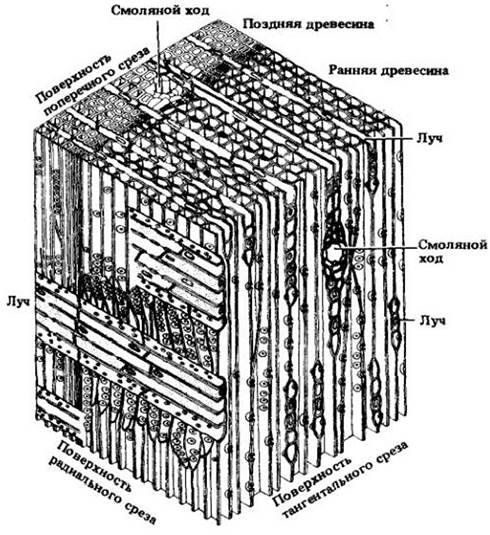
Рис. 23-20. Трехмерная схема вторичной ксилемы сосны веймутовой (Pinus sirobus). За исключением паренхимных клеток, окружающих смоляные ходы, осевая система целиком состоит из трахеид. Лучи, кроме содержащих смоляные ходы, однорядные. Ранняя и поздняя древесина описаны на с. 87
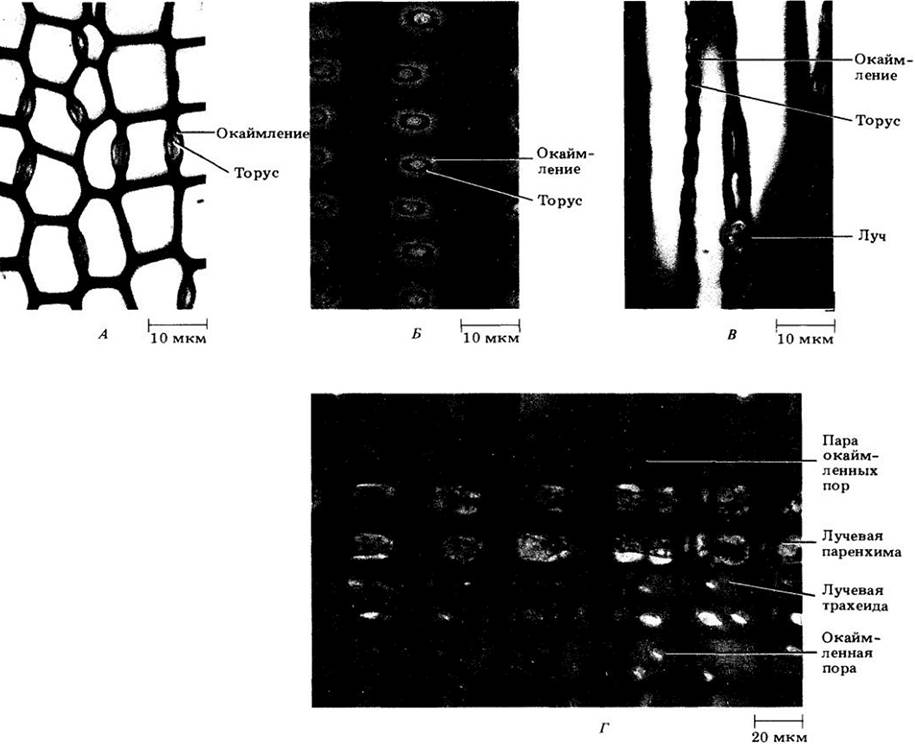
Для трахеид хвойных характерны крупные округлые окаймленные поры, которых особенно много на концах клеток, в области смыкания с другими трахеидами (рис. 23-20 — 23-22). Все пары пор (см. гл. 2, рис. 2-32) между ними имеют торус, т. е. утолщение в центральной части поровой мембраны (см. рис. 7.3-25), которое несколько шире отверстия, или апертуры, окаймленной поры (рис. 23-22). Поровая мембрана гибкая, поэтому иногда торус блокирует одну из апертур, задерживая таким образом прохождение через пару пор воды или газов (рис. 23-22).
На рис. 23-20 показана трехмерная схема строения древесины сосны веймутовой (Pinus strobus), построенная по трем срезам, изображенным на рис. 23-21. На срезе, проходящем перпендикулярно длинной оси корня или стебля (поперечном), трахеиды выглядят много- или прямоугольниками; можно также видеть вытянутые, пересекающие древесину лучи (рис. 23-21, А). Продольные срезы могут быть радиальными и тангентальными. Первые проходят параллельно лучам, которые на них выглядят как пластины из клеток, расположенных под прямыми углами к вытянутым вертикально трахеидам осевой системы (рис. 23-21, Б и 23-22, Г). Тангентальные срезы перпендикулярны лучам и показывают их ширину и высоту. У сосен лучи, за исключением содержащих смоляные ходы, однорядные, т. е. шириной в одну клетку (рис. 23-21, В). Детали строения древесины веймутовой сосны показаны на рис. 23-22.
Рис. 23-21. Древесина сосны веймутовой на поперечном (А), радиальном (Б) и тангентальном (В) срезах
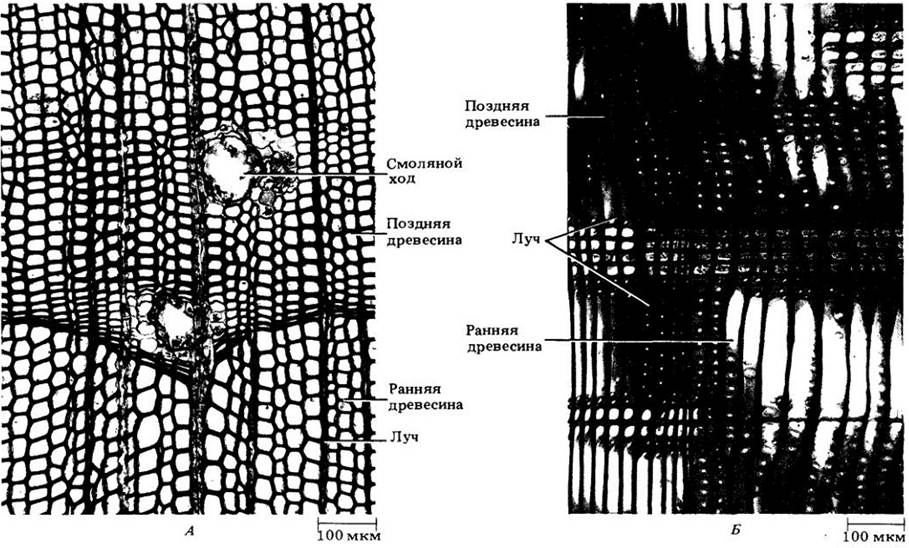
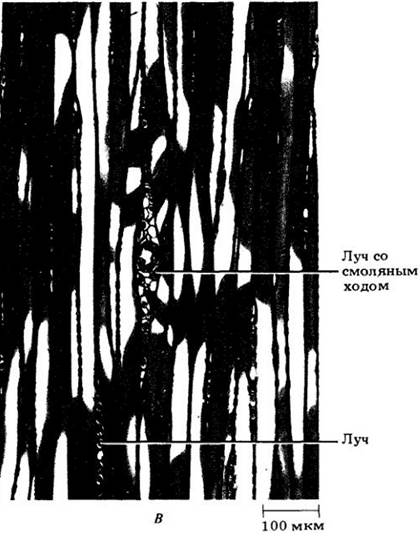
Рис. 23-22. Детали строения древесины сосны веймутовой. А. Поперечный срез с парами окаймленных пор трахеид. Б. Радиальный срез; фронтальный вид пар окаймленных пор в стенках трахеид. В. Тангентальный срез с парами окаймленных пор трахеид. Г. Радиальный срез с лучом. Лучи сосны и других хвойных состоят из лучевых трахеид и лучевых паренхимных клеток. Заметны окаймленные поры этих трахеид
Древесина двудольных
Структура древесины двудольных гораздо разнообразнее, чем у хвойных, отчасти из-за большого числа клеточных типов в осевой системе, содержащей членики сосудов (характерная особенность), трахеиды, несколько типов волокон и паренхимные клетки (рис. 23-23 и 23-24; см. также рис. 20-10 и 23-26).
Лучи здесь часто значительно крупнее. Если в древесине хвойных они преимущественно имеют одну клетку в ширину и большей частью 1 — 20 клеток в высоту, то у двудольных их ширина доходит до многих клеток, а высота до нескольких сотен. У некоторых пород, например, дуба, крупные лучи можно видеть невооруженным глазом (см. рис. 23-13). У дуба красного (рис. 23-24, В) их ширина от 12 до 30 клеток, а высота — сотни клеток. Наряду с крупными здесь много однорядных лучей. В древесине этой породы лучи составляют в среднем около 21% объема. В среднем в твердой древесине на их долю приходится около 17%, а у хвойных — около 8% объема.
Как и у хвойных, на поперечных срезах древесины двудольных видны радиальные ряды клеток осевой и радиальной систем, происходящих из камбиальных инициалей (рис. 23-23 и 23-24). Однако в последнем случае они не всегда такие правильные, как у хвойных, так как разрастание сосудов и удлинение волокон смещают многие клетки с исходных мест. Сдвигание лучей члениками сосудов хорошо видно на поперечном срезе дуба красного (Quercus rubra), показанном на рис. 23-23, А.
Кольца прироста
В результате периодической активности камбия, которая в умеренных зонах представляет собой сезонное явление, образуются концентрические слои прироста во вторичных ксилеме и флоэме (в последнем случае не всегда хорошо различимые). Если они соответствуют одному сезону, то называются годичными кольцами. Резкие изменения в содержании доступной воды и в других факторах окружающей среды иногда способствуют образованию за год нескольких таких колец, называемых «ложными» личными кольцами. Таким образом, возраст любой части старого древесного ствола можно определять путем подсчета для нее годичных колец, но ложные кольца в ряде случаев искажают получаемые результаты.
Ширина отдельных слоев прироста может сильно варьировать по годам в зависимости от освещенности, температуры, осадков, доступной почвенной влаги, продолжительности вегетационного периода и других экологических факторов. Она достаточно хорошо коррелирует с количеством выпавших в соответствующем году осадков. При благоприятных условиях, т. е. в периоды достаточных или обильных дождей, кольца широкие, при неблагоприятных — узкие.
В семиаридных зонах, где дождей мало, деревья — весьма чувствительные «дождемеры». Прекрасный пример такого рода — сосна остистая (Pinus longaeva) в западной части Большого Бассейна (рис. 23-27). Все годичные кольца у нее разные, и изучение их позволяет заглянуть на тысячи лет назад. Самому старому из ныне живущих известных экземпляров этой сосны 4900 лет. Дендрохронологи (ученые, исследующие прошлое по годичным кольцам), сопоставляя древесину живых и отмерших деревьев, реконструировали непрерывные ряды колец продолжительностью более 8200 лет. Установлено, что ширина их у сосны остистой в высокогорьях (у границы древесной растительности) тесно связана с изменениями температуры, т. е. может использоваться для оценки климатических условий в прошлом. Например, в горах Уайт-Маунтинс (Калифорния) в 3500 — 1300 гг. до н. э. лето было относительно теплым, и эта граница проходила примерно на 150 м выше ее современного уровня. Период 1300 — 200 г. до н. э. отличался холодным летом.
Структурная основа разграничения годичных колец — разница в плотности древесины, образованной в начале и в конце вегетационного периода (см. рис. 23-21,23-23 и 23-24). Ранняя древесина менее плотная (с более широкими клетками, имеющими относительно тонкие стенки), чем поздняя (с более узкими и толстостенными клетками). Переход на срезе от ранней к поздней древесине того же года может быть очень постепенным и почти неуловимым. Однако граница поздней древесины одного кольца с ранней древесиной следующего всегда резкая и хорошо заметная.
Рис. 23-23. Поперечные срезы древесины с годичными кольцами. А. Дуб красный (Quercos rubra). Крупные сосуды кольцепоровой древесины приурочены к ее ранней части. Темные вертикальные линии — лучи. Б. Рассеяннопоровая древесина тюльпанного дерева (Liriodendron ndipifera)

Рис. 23-24. Древесина дуба красного (Quercus rubra) на поперечном (А), радиальном (К) и тангентальном (В) срезах
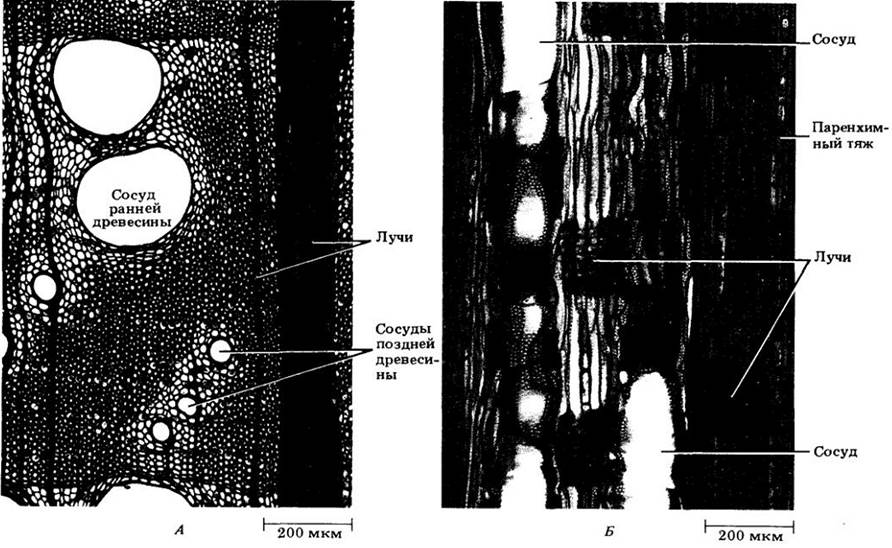
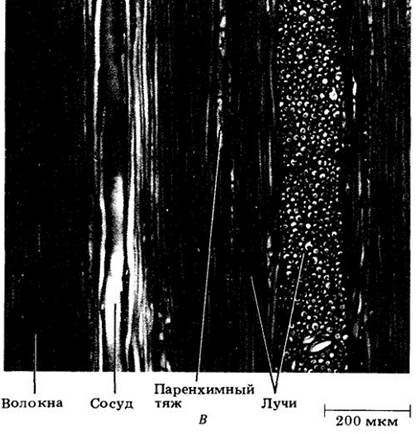
Рис. 23-25. Сканирующая электронная микрофотография поровой мембраны пары окаймленных пор трахеиды сосны веймутовой. Утолщенная часть — торус. Окружающая его часть мембраны называется марго
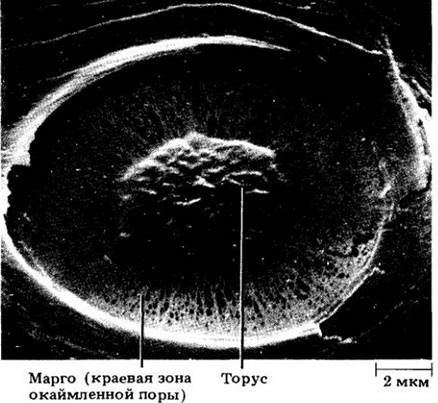
Рис. 23-26. Сканирующая электронная микрофотография трех поверхностей среза древесины ильма американского (Ulmus americana). Сравнивая этот снимок с рис. 23-20, 23-23 и 23-24, можно распознать каждую поверхность. Такая полукольцепоровая древесина с волнистыми рядами поздних сосудов — характерная черта ильмов. Найдите сосуды ранней и поздней древесины и лучи на всех трех поверхностях. Плотная часть древесины состоит главным образом из волокон. Клетки осевой паренхимы также присутствуют, но при таком увеличении они неразличимы

Рис. 23-27. А. Сосна остистая (Pinus longaeva) в горах Уайт-Mayнтинс (восточная Калифорния). Эти деревья, растущие близ верхней границы древесной растительности, — самые старые из ныне живущих; одному из них 4900 лет. Б. Поперечный срез древесины сосны остистой с различной шириной годичных колец. Начало этого среза соответствует примерно 6260 лет назад; темным выделена группа колец, образовавшихся с 4240 по 4210 г. до н. э. Совпадения в чередовании широких и узких колец на спилах мертвых деревьев позволяют определить относительное количество осадков в разные годы примерно на протяжении последних 8200 лет. Несмотря на достигающий 4900 лет возраст, сосны остистые, возможно, не самые старые на планете живые существа. В самом деле, известен, по- видимому, возникший из одного семени клон кустарника ларреи (Larrea divaricata), которому, как предполагают, около 12 000 лет. Этот кольцеобразный так называемый «Королевский клон» (см. рис. 32-13) растет в пустыне Мохаве примерно в 250 км к северо-востоку от Лос-Анджелеса (Калифорния)
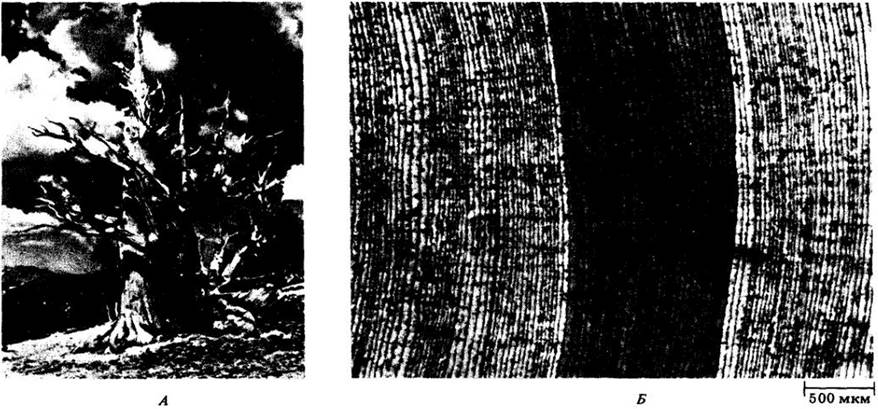
У некоторых двудольных, различия в диаметре сосудов, или «пор», ранней и поздней древесин четко выражены: первые гораздо шире вторых (термин «пора» используется дендроанатомами для обозначения поперечного среза сосуда). Такую древесину называют кольцепоровой (кольцесосудистой) (см. рис. 23-23, А и 23-24, А). У других двудольных сосуды распределены в годичном слое равномерно и почти не различаются размерами. Такую древесину называют рассеяннопоровой (рассеяннососудистой) (рис. 23-23, Б). В кольцепоровой древесине почти вся вода проводится наружным годичным кольцом примерно в 10 раз быстрее, чем в рассеяннопоровой древесине.
Заболонь и ядро
По мере того как древесина стареет и перестает выполнять проводящую функцию, ее паренхимные клетки отмирают. Однако до этого в ней часто происходят заметные изменения, включая утрату запасных питательных веществ и пропитывание различными веществами (маслами, камедями, смолами, таннинами и др.), окрашивающими, а иногда и ароматизирующими ткани. Такую обычно более темную непроводящую древесину называют ядровой (ядром), а, как правило, более светлую проводящую — заболонью (см. рис. 23- 13). У многих пород при прекращении функционирования сосудов в них образуются тилы (рис. 23-28), пузыревидные выросты лучевых или осевых паренхимных клеток, проникающие в сосуды через поры в их стенках и способные полностью закупорить их просвет. Часто они возникают преждевременно под влиянием патогенов и в этом случае выполняют защитную функцию, препятствуя распространению болезнетворного агента по ксилеме.
Соотношение заболони и ядра и степень видимых различий между ними весьма неодинаковы у разных видов. Некоторые деревья, например, клен (Acer), береза (Betula), ясень (Fraxinus), имеют толстую заболонь, в то время как другие, в частности белая акация (Robinia), катальпа (Catalpa) и тисс (Taxus), — тонкую. У третьей группы пород, например, у тополя (Populus), ивы (Salix) и пихты (Abies), заболонь и ядро четко не разграничены.
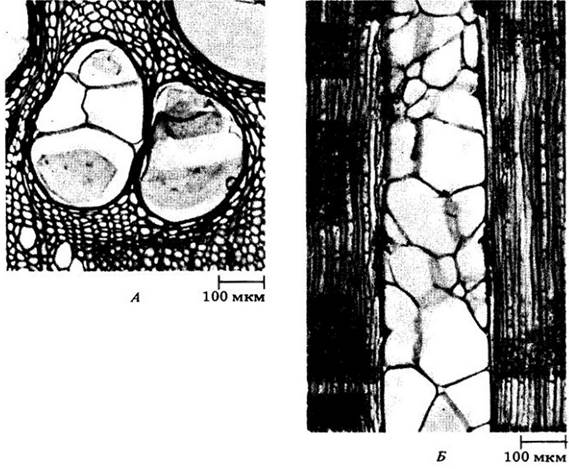
Рис. 23-28. Тилы — пузыревидные выросты паренхимных клеток, частично или полностью блокирующие просвет сосуда. Гилы в сосудах дуба белого (Quercus alba) на поперечном (А) и продольном (Б) срезах в световом микроскопе
Реактивная древесина
Реактивной древесиной называют аномалии вторичной ксилемы, свойственные склоненным стволам и ветвям. Ее образование связано с процессом выпрямления этих частей растения. У хвойных реактивная древесина развивается на нижней стороне изогнутой части (древесина сжатия, или креневая), а у двудольных — на верхней (древесина растяжения).
Древесина сжатия образуется за счет возрастания камбиальной активности на нижней стороне ствола, в результате чего возникают эксцентрические годичные кольца с нижними частями гораздо шире верхних (рис. 23-29). При этом выпрямление ствола или ветви происходит за счет разрастания тканей, толкающих ствол к вертикали. В этой древесине больше лигнина и меньше целлюлозы, чем в нормальной, а усадка се в длину при сушке часто в 10 или более раз сильнее (у нормальной обычно не более 0,1 — 0,3%). Различие в относительной усадке нормальной и креневой древесин в сохнущей доске часто приводит к ее скручиванию или принятию чашевидной формы. Такие пиломатериалы годятся только на дрова.
Рис. 23-29. Поперечный срез ствола тсуги с креневой древесиной (более широкие годичные кольца ни нижней стороне). Трещины образуются при высыхании

Древесина растяжения образуется за счет возрастающей активности камбия на верхней стороне ствола. Как и креневую, ее можно обнаружить по присутствию эксцентрических колец нарастания (рис. 23-30). Чтобы выпрямить ствол, она должна «тянуть» его за собой (отсюда и ее название). Точная идентификация древесины растяжения требует микроскопического изучения срезов ствола. Главный ее анатомический признак — присутствие студенистых волокон, характеризующихся слабой лигнификацией или полным ее отсутствием и желеобразным видом части вторичной оболочки. Усадка в длину при высыхании древесины растяжения редко превышает 1%, но содержащие се доски скручиваются. При распиловке в сыром виде она распускается на пучки волокон, что придает доске шероховатость.
Рис. 23-30. Поперечный срез ствола дуба красного (Quercus rubra) с древесиной растяжения (более широкие годичные кольца на верхней стороне)
Макроскопические признаки древесины
Внешний вид древесины определяется ее цветом, волокнистостью, текстурой и рисунком. Некоторые из этих характеристик не только позволяют различать происхождение древесины, но и обусловливают ее декоративность.
Цвет варьирует в зависимости от типа древесины и породы растения. У ядровой древесины он может иметь важное значение для ее и идентификации, а кроме того, отчасти определяет предпочтительное направление использования. Так, темно-шоколадное или пурпурно-бурое ядро ореха черного (Juglans nigra) и красно-бурое у черемухи поздней (Prunus serótina) — традиционный излюбленный материал для изготовления высококачественной мебели.
Волокнистость — термин, указывающий на относительное расположение всей совокупности элементов древесины — волокон, трахеид, паренхимных клеток и члеников сосудов. Если все они ориентированы параллельно продольной оси, говорят, что древесина прямослойная. Если их расположение не совпадает с продольной осью участка ствола, древесину называют косослойной, а если оно спиральное — тангентально-косослойной. В последнем случае, если удалить с бревна кору, оно выглядит скрученным (рис. 23-31). Если направление спиралей меняется по радиусу бревна на противоположное через более или менее правильные интервалы, древесина будет сложно-косослойной.
Рис. 23-31. Отмерший ствол дуба белого (Quercus alba) с опавшей корой. Видна тангентально-косослойная древесина

Текстура характеризует относительные размеры и их изменчивость у элементов внутри годичного кольца. Грубая текстура означает присутствие обширных зон крупных сосудов и широких лучей. Она свойственна некоторым кольцепоровым породам. Тонкая текстура — у древесины с мелкими сосудами и узкими лучами. Однородная текстура возникает при невыраженных различиях в размерах клеток, и соответственно между ранней и поздней древесиной, а неоднородная — при явной разнице между ними в годичном кольце.
Рисунок — термин, означающий узор, заметный на продольных спилах дерева. В узком смысле он применяется для характеристики декоративной древесины, высоко ценимой в производстве мебели и при отделке помещений. Рисунок зависит от волокнистости, текстуры и направления спила.
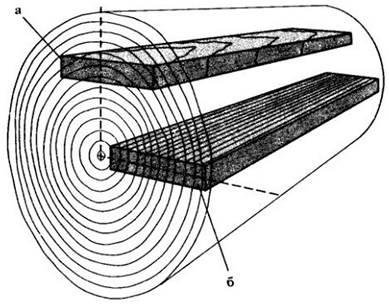
Доску можно выпилить из бревна двумя способами (рис. 23-32). В одном случае ее широкие поверхности окажутся примерно параллельными тангентально-продольной плоскости бревна (тангентальная распиловка). Годичные кольца на такой доске выглядят волнообразными полосами. При втором способе доска проходит вдоль бревна через его центр (радиальная распиловка). При этом годичные кольца выглядят параллельными линиями, проходящими по всей длине доски и пересекающимися под прямым углом лучами. Такие доски во многих случаях предпочтительнее, так как радиальные поверхности более однородны с точки зрения износостойкости и обрабатываемости. Однако радиальная распиловка требует больше времени и часто менее экономична, чем тангентальная.
Рис. 23-32. Схема, показывающая направления тангентальной (а) и радиальной (б) распиловки бревна на доски. В первом случае годичные кольца проходят более или менее параллельно широким поверхностям доски, во втором — почти перпендикулярно им
Заключение
Вторичный рост (увеличение толщины в зонах, завершивших удлинение) происходит у всех голосеменных и большинства двудольных. Он связан с активностью двух латеральных меристем — камбия и пробкового камбия, или феллогена. У травянистых растений вторичный рост слабый или вообще отсутствует, в то время как деревья и кустарники могут расти в толщину многие годы. На рис. 23-33 представлена схема развития корня и стебля древесного растения от апикальной меристемы до вторичных тканей, сформированных за первый вегетационный период.
Рис. 23-33. Схемы развития стебля и корня деревянистых двудольных в первый год роста. Звездочками отмечены ткани, вместе образующие перидерму
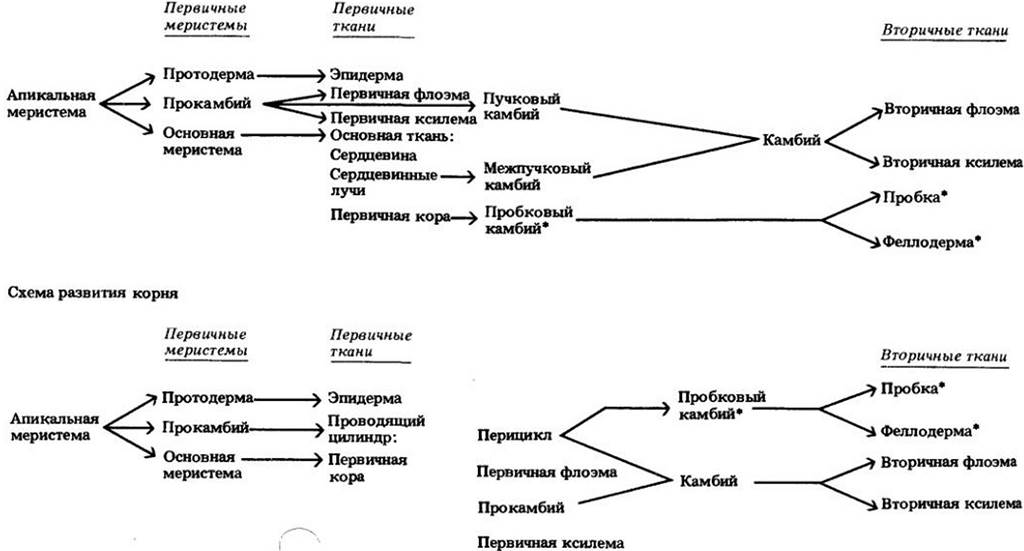
Камбий содержит два типа инициалей — веретеновидные и лучевые. За счет периклинальных делений первые из них дают начало компонентам осевой системы, а вторые — клеткам лучей, образующим проводящие лучи, или радиальную систему. Нарастание камбия по окружности обусловлено антиклинальными делениями инициалей.
Первый пробковый камбий в большинстве стеблей закладывается в слое клеток непосредственно под эпидермой, а в корне — в перицикле. Этот камбий откладывает наружу пробку, а внутрь феллодерму и вместе с ними составляет перидерму. Хотя большая часть перидермы состоит из плотно упакованных клеток, отдельные ее участки, называемые чечевичками, имеют многочисленные межклетники.
Кора включает все ткани снаружи от камбия. В старых корнях и стеблях большая часть флоэмы в се составе нефункциональна. Ситовидные элементы недолговечны и, как правило, являются проводящими, или функциональными, только в кольце прироста текущего года. Вслед за первой перидермой многократно и каждый раз все глубже во вторичной коре из паренхимных клеток нефункциональной флоэмы формируются последующие перидермы.
Древесину обычно подразделяют на мягкую и твердую; первую дают хвойные породы, а вторую — двудольные. По сравнению с древесиной двудольных у хвойных она проще и состоит из трахеид и паренхимных клеток. У некоторых пород присутствуют смоляные ходы. Древесина двудольных может содержать одновременно членики сосудов, трахеиды, несколько типов волокон, паренхимные клетки.
Слои прироста за год называются годичными кольцами. Разница в плотности между поздней древесиной одного кольца и ранней древесиной следующего позволяет провести между ними границу. Плотность древесины — хороший показатель ее прочности.
У многих растений можно визуально отличить непроводящую ядерную древесину от активно проводящей заболони.
Обычно на нижней стороне наклонных стволов и ветвей хвойных и на верхней их стороне у двудольных развивается реактивная древесина. Ее образование способствует выпрямлению частей растения. У хвойных она называется древесиной сжатия, а у двудольных — древесиной растяжения.
Таблица 23-1. Использование древесины некоторых широкораспространенных североамериканских пород
Ольха красная (AInus rubra) — основная лиственная порода тихоокеанского северо-запада. Используется в производстве мебели, особенно стульев, оконных переплетов, дверей и других столярных изделий, фанеры, древесного угля; важное сырье для бумажного производства.
Ясень американский (Fraxinus americana). Черенки и рукоятки, особенно длинные (к лопатам, граблям, заступам), в связи с прямослойной волокнистостью, прочностью, умеренной плотностью и другими достоинствами; почти все бейсбольные биты, весла, теннисные ракетки, хоккейные клюшки; кухонные шкафы, игрушки и деревянная посуда.
Липа американская (Tilia americana). Фанерный шпон для декоративных панелей и отделки мебели; мягкая упаковочная стружка; оконные переплеты, двери и другие столярные изделия; фортепьянные клавиши; ящики и упаковочные клети; сундуки и гробы.
Бук американский (Fagus americana) — одна из трех важнейших северных пород (две другие — береза желтая и клен сахарный). Доски. особенно идущие на настилку полов; фанерный шпон; топливо; перегонка до уксусной кислоты, метанола и других химических веществ; игрушки и деревянная посуда.
Береза. Betula alleghaniensis. Фанерный шпон; перегонка; шпалы; мебель; игрушки и деревянная посуда; музыкальные инструменты; зубочистки.
Черемуха поздняя (Prunus serótina). Прекрасное сырье для краснодеревщиков; резная мебель; колодки для типографских клише; фортепьянные молоточки; отделка интерьеров; панели; рукоятки; игрушки и деревянная посуда.
Тополь дельтовидный (Populus deltoides). Масса для изготовления высококачественной бумаги, применяемой для печатания книг и журналов; фанеровочный материал для мебели; мягкая упаковочная стружка; кадки и бадьи для пищевых продуктов; ящики и упаковочные клети.
Ильм (Ulmus spp.). Имеет сложно-косослойную древесину, вследствие чего трудно расщепляется. Бочарная клепка и обручи; ящики и упаковочные клети; фанерный шпон для фруктовых и овощных контейнеров и круглых коробок для сыра; гнутые части мебели; отделка интерьеров.
Кария сердцевидная (Carya cordyformis). Ручки к инструментам, особенно топорища, черенки заступов; санные полозья; лестницы, мебель, деревянная посуда; прекрасные дрова для копчения мяса; первосортное топливо.
Белая акация (Robinia pseudo-acacia). Крепежный лес для шахт; шпалы; столбы для заборов; сооружения, от которых требуются прочность и долговечность.
Клен сахарный (Acer saccharum). Фанерный шпон; перегонка; шпалы, топливе); мебель; полы, особенно для кегельбанов и танцплощадок; игрушки и деревянная посуда; музыкальные инструменты.
Дубы красный и белый (Quercus rubra, Q. alba). Шпалы; фанерный шпон; полы; оконные переплеты, двери и другие столярные изделия; топливо; корабельный лес; сундуки и гробы.
Хурма виргинская (Diospyros virginiana). Ткацкие челноки, катушки и веретена; биты для гольфа; ящики и упаковочные клети; рукоятки.
Платан западный (Platanus occidentales). Имеет сложно-косослойную древесину. Фанерный шпон; ящики и упаковочные клети; отделка интерьера, панели; бочки для муки и сахара; фанеровочный материал для мебели.
Орех черный (Jugions nigra). Прекрасное местное сырье для краснодеревщиков в континентальной части США; фанерный шпон для мебели; высокосортные столы и стулья; основная древесина для ружейных прикладов; сундуки и гробы.
Тюльпанное дерево (Liriodendron tulipifera). Фанерный шпон для отделки интерьеров, мебели и других тонких столярных работ; сырье для изготовления бумаги; ящики и упаковочные клети; оконные переплеты, двери и другие столярные изделия.
Дугласия (Pseudotsuga menziesii). Порода, образующая примерно 50% лесов запада США и дающая больше деловой древесины, чем любой другой вид США. Строевой лес; фанерный шпон, главным образом для клееной фанеры; шпалы; крепежный лес; ящики и упаковочные клети; масса для изготовления бумаги; корабельный лес.
Тсуга канадская (Tsuga canadensis). Производство бумаги; строевой лес; ящики и упаковочные клети; оконные переплеты и двери, кухонная мебель.
Сосна горная веймутова (Pinus montícola). Спички; ящики и упаковочные клети; оконные переплеты, двери и другие столярные изделия; строевой лес; внутренняя часть клееной фанеры, особенно для столешниц.
Сосна желтая (Pinus ponderosa). Ящики и упаковочные клети, оконные переплеты, двери и другие столярные изделия; строевой лес; мачты; сваи, балясины, колонны; столбы; игрушки; сундуки и гробы.
Сосна Эллиота (Pinus elliottii). Масса для изготовления бумаги; крупномерные лесоматериалы; шпалы; фанерный шпон; скипидар и канифоль; ящики; кузова и упаковочные клети.
Сосна Ламберта (Pinus lambertiana). Ящики и упаковочные клети; оконные переплеты, двери и другие столярные изделия; вывески; фортепьянные клавиши и органные трубы.
Секвойя вечнозеленая (Sequoia sempervirens). Строевой и корабельный лес; сигарные и кондитерские ящики; садовая мебель; кровельная дрань; сундуки и гробы.
Ель красная (Picea rubens). В основном бумажная масса; хорошо резонирующая древесина для музыкальных инструментов; весла; лестничные перила; корабельный лес; ящики и упаковочные клети.
Приложение. Плотность древесины
Плотность — единственный важнейший показатель прочности древесины, который можно использовать для характеристики ее твердости, сопротивления пробиванию и легкости обработки. Плотная древесина, как правило, усыхает и набухает сильнее, чем легкая. Кроме того, первая дает лучшее топливо.
Плотность сухого твердого вещества древесины (т. е. воздушно-сухого материала клеточных оболочек) всех растений равна примерно 1,5 г/см3. Поэтому различия в плотности древесины в целом зависят от соотношения в ней клеточных оболочек и пустот между ними. Особенно большое значение при этом имеют волокна. Если они толстостенные и с узкими просветами, плотность высока. И напротив, если волокна тонкостенные с широкими просветами, она низкая. Присутствие многочисленных тонкостенных сосудов также понижает плотность.
Плотность древесины некоторых североамериканских лиственных пород, г/см3
Ясень американский (Fraxinus americana) 0,55
Тополь осинообразный (Populus tremuloides) 0,35
Бальза заячья (Ochroma lagopus) 0,12
Липа американская (Tilia americana) 0,32
Бук американский (Fagus americana) 0,56
Береза желтая (Betula lútea) 0,55
Конский каштан голый (Aesculus glabra) 0,33
Орех серый (Juglans cinérea) 0,36
Черемуха поздняя (Prunus serótina) 0,47
Тополь дельтовидный (Populus deltoides) 0,37
Ильм американский (Ulmus americana) 0,46
Кария овальная (Caria ovala) 0,64
Гледичия трехколючковая (Gleditsia triacanthos) 0,60
Кругиодендрон железный (Krugiodendron ferreum) 1,30
Гваякум лекарственный (Guaiacum officinale) 1,25
Белая акация (Robinia pseudoacacia) 0,66
Магнолия крупноцветковая (Magnolia grandiflora) 0,46
Клен красный (Acer rubrum) 0,49
Клен сахарный (Acer saccharum) 0,56
Дуб виргинский (Quercus virginiana) 0,81
Дуб красный (Quercus rubra) 0,57
Дуб белый (Quercus alba) 0,59
Маклюра оранжевая (Madura pomífera) 0,76
Хурма виргинская (Diospyros virginiana) 0,64
Амбровое дерево (Liquidambar styraciflua) 0,46
Платан западный (Platanus occidentalis) 0,46
Орех черный (Juglans nigra) 0,51
Ива черная (Salix nigra) 0,34
Плотность древесины некоторых североамериканских хвойных пород, г/см3
Болотный кипарис (Taxodium distichum) 0,42
Дугласия (Pseudotsuga menziesii) 0,45
Пихта бальзамическая (Abies balsamea) 0,34
Тсуга канадская (Tsuga canadensis) 0,38
Тсуга западная (Tsuga heterophylla) 0,42
Кедр ладанный (Calocedrus decurrens) 0,35
Лиственница западная (Larix occidentalis) 0,48
Сосна веймутова (Pinus strobus) 0,34
Сосна ладанная (Pinus taeda) 0,47
Сосна скрученная (Pinus contorta) 0,38
Сосна желтая (Pinusponderosa) 0,38
Сосна Эллиота (Pinus elliottii) 0,64
Сосна Ламберта (Pinus lambertiana) 0,35
Секвойя вечнозеленая (Sequoia sempervirens) 0,38
Ель черная (Picea mariana) 0,36
Ель Энгсльмана (Picea engelmannii) 0,38
Лиственница американская (Larix laricina) 0,49
Тисс коротколистный (Taxus brevifolia) 0,60
Плотность выражается массой единицы объема, обычно в граммах на кубический сантиметр. У воды она равна 1 г/см3. Если плотность древесины 0,5 г/см3 — значит этот материал вдвое легче воды. В книге мировых рекордов Гиннеса сказано, что у маслины Olea capensis из Южной Африки древесина самая тяжелая, а у вида Aeschynomene hispida с Кубы — самая легкая. Их плотность равна соответственно 1,49 и 0,044 г/см3. У большинства используемых в хозяйстве видов древесины она составляет 0,35 — 0,65 г/см3.