СОВРЕМЕННАЯ БОТАНИКА - П. РЕЙВН - 1990
РАЗДЕЛ VI. РЕГУЛЯЦИЯ И РОСТОВЫЕ РЕАКЦИИ
ГЛАВА 25. ВНЕШНИЕ ФАКТОРЫ И РОСТ РАСТЕНИЯ
Живые существа должны приспосабливаться к окружающим условиям внешней среды. Многие животные, будучи подвижными, могут в какой-то мере менять окружающую обстановку, т. е. перемещаться в пространстве в поисках пищи, ухаживая за половым партнером, разыскивая убежище и даже строя его в плохую погоду. Растение, напротив, с появлением первого корня становится неподвижным. Однако оно способно реагировать на разнообразные изменения внешней среды и приспосабливаться к ним. Это обусловлено главным образом изменением характера роста.
Тропизмы
Ростовая реакция, вызывающая изгибание или искривление части растения в сторону внешнего стимула, определяющего направление движения, или от него, называется тропизмом. Если движение направлено к стимулу, говорят о положительном тропизме; если в обратную сторону — об отрицательном.
Пожалуй, наиболее известное взаимодействие между растениями и внешней средой выражается в изгибании растущих верхушек побегов по направлению к свету (см. рис. 24-2). Эта ростовая реакция, называемая фототропизмом, обусловлена действием ауксина (ИУК), вызывающего растяжение клеток теневой стороны верхушки побега. Какую роль играет здесь свет? На этот вопрос возможны три ответа: свет (1) уменьшает чувствительность клеток освещенной стороны к ауксину, (2) разрушает ауксин, (5) способствует перемещению ауксина на теневую сторону растущей верхушки побега.
Чтобы подтвердить какую-либо из этих гипотез, Уинслоу Бриггс и его сотрудники провели серию экспериментов, основанных на более ранней работе Ф. Вента (см. рис. 24-3). Исследователи установили, что в верхушке побега как на свету, так и в темноте образуется одинаковое общее количество ИУК. Однако после воздействия света количество гормона, поступающее от теневой стороны, больше, чем от освещавшейся. Если верхушку расщепить и между двумя ее половинками поместить барьер в виде тонкого кусочка стекла, различия в распространении ИУК не будет. Другими словами, Бригес ясно продемонстрировал, что ауксин (или, возможно, его предшественник) перемещается от освещенной стороны к затененной и что искривление побега является реакцией на неравномерное его распределение. Эксперименты с использованием ИУК, меченной 14С, определенно показали, что мигрирует именно ауксин.
Далее было отмечено, что наиболее эффективно перемещение этого гормона в верхушке побега стимулирует свет с длиной волны от 400 до 500 нм, т. е. в реакции в качестве медиатора участвует пигмент, поглощающий синий свет. Структура фоторецептора синего света (медиатора также многих других реакций у растений и грибов) окончательно еще не установлена, но имеющиеся данные показывают, что это желтоокрашенный пигмент, называемый флавином. Согласно широко известной гипотезе, которая еще окончательно не доказана, синий свет вызывает химическую реакцию восстановления флавина, и вследствие этого усиливается транспорт электронов через плазматическую мембрану и устанавливается градиент pH.
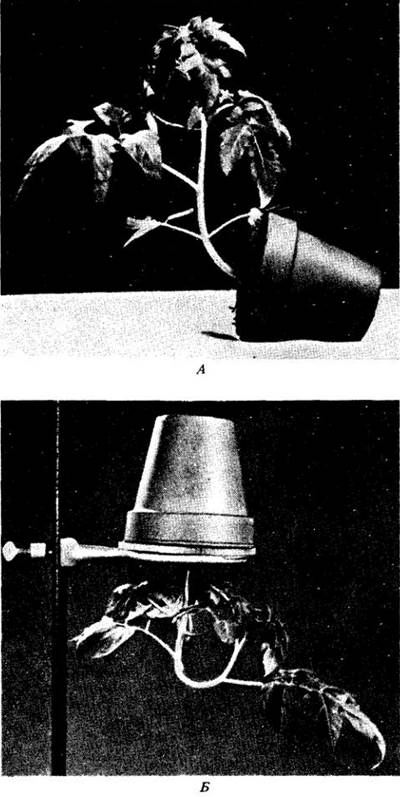
Другим известным тропизмом является гравитропизм, или геотропизм, — реакция на силу тяжести, отчетливо проявляющаяся у проростков. Если проросток положить горизонтально, его корень изогнется вниз (положительный геотропизм), а побег будет расти вверх (отрицательный геотропизм). Первоначально это явление объясняли асимметричным перераспределением ауксина к нижней стороне органа путем латерального полярного транспорта молекул сверху вниз. В таких условиях нижняя сторона побега должна расти вверх (рис. 25-1). У корня верхняя сторона будет удлиняться быстрее, чем нижняя, и он изогнется вниз. Когда каждая часть растения займет вертикальное положение, латеральная асимметрия в концентрации ауксина исчезнет и рост будет продолжаться в вертикальном направлении.
Рис. 25-1. Гравитропическая реакция у побега молодого растения томата (Lycopersicon esculentum). А. Горшок с растением был положен на бок и находился в неподвижном состоянии. Б. Горшок с растением переворачивали и помещали вверх дном на кольцевую подставку. Стебли, первоначально прямые, изгибались и росли вверх. Если горшок с растением положить горизонтально и медленно вращать вдоль оси стебля, изгибания (гравитропической реакции) не произойдет: растение будет продолжать расти горизонтально. Можете ли вы объяснить различия в росте вращаемого в горизонтальной плоскости растения и растений, показанных здесь?
Обоснованность этой гипотезы применительно к гравитропизму в настоящее время вызывает сомнения. Хотя было показано существование латерального транспорта ИУК по направлению к нижней стороне колеоптилей кукурузы (Zea) и овса (Avena) и ее асимметричное распределение у этих растений, в отношении побегов двудольных остается много неясного. Более того, достаточно убедительно нс установлено, что асимметричное распределение ИУК в колеоптилях кукурузы и овса само по себе может вызвать изменение скорости роста.
Проблема асимметрии в распределении регуляторов роста в корне менее изучена. Сейчас доказано, что ИУК, цитокинины, гиббереллины и АБК присутствуют в корнях, хотя их физиологическая роль неясна. Вещества, тормозящие рост, тоже обнаружены в корнях, и во всяком случае одно из них образуется в корневом чехлике. Есть сведения, что ингибитор из корневого чехлика может быть связан с положительным гравитропизмом первичного корня.
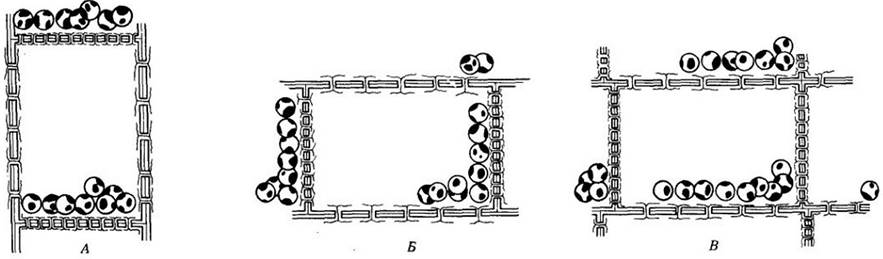
Имеется много данных в пользу того, что восприятие силы тяжести связано с осаждением амилопластов (пластид, содержащих крахмал) в специализированных клетках побега и корня. Подобные клетки присутствуют вдоль всего побега, часто входя в состав обкладок проводящих пучков. В корнях они локализованы в корневом чехлике, особенно в его центральной части (колонке) (рис. 25-2). Когда корень помещают горизонтально, пластиды, располагавшиеся у поперечных клеточных стенок вертикально растущих корней, перемещаются вниз и задерживаются вблизи клеточных стенок, ранее ориентированных вертикально (рис. 25-3). Через несколько часов корень изгибается вниз и пластиды возвращаются в первоначальное положение, т. е. скапливаются вдоль поперечных стенок. Вопрос о том, каким образом движение этих рецепторов гравитации (статолитов) приводит к формированию гормональных градиентов, еще нуждается в убедительном объяснении. Недавно было высказано предположение, что ключевую роль в сопряжении гравирецепции и гравитропизма в корнях играет кальций, регулирующий транспорт гормонов. Как известно, ионы кальция обнаружены в амилопластах клеток колонки.
Рис. 25-2. А. Микрофотография срединного продольного среза корневого чехлика первичного корня фасоли (Phaseolus vulgaris). Стрелки указывают на амилопласты (пластиды, содержащие крахмал), сгруппировавшиеся вблизи поперечных оболочек центральных клеток корневого чехлика. Б. Электронная микрофотография паренхимных клеток корневого чехлика, сходных с теми, которые показаны слева. Здесь тоже амилопласты (стрелки) оседают у основания каждой клетки вблизи поперечных стенок
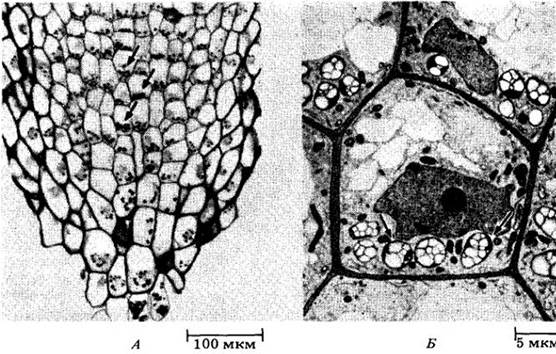
Рис. 25-3. Рисунок показывает, как реагируют на гравитацию амилопласты (пластиды, содержащие крахмал) в паренхимных клетках корневого чехлика. А. В корневом чехлике корня, растущего вертикально вниз, амилопласты обычно оседают около поперечных стенок. Если тот же корень располагается горизонтально (Б, В), амилопласты опускаются вниз и оседают у тех стенок, которые в норме являются вертикальными, а теперь параллельны поверхности почвы. Перемещение статолитов играет важную роль в создании градиентов ростовых веществ, обеспечивающих вертикальный рост корня
Менее понятным, хотя и распространенным тропизмом является тигмотропизм (от греческого thigma — прикосновение) — реакция на контакт с твердым предметом. Один из наиболее обычных примеров тигмотропизма демонстрируют усики, которые у одних растений представляют собой модификации листьев, а у других — стеблей (см. гл. 22). Усики обвиваются вокруг любого предмета, с которым соприкасаются (рис. 25-4), давая возможность растению цепляться за опору и карабкаться вверх. Реакция может быть быстрой; усик способен менее чем за час один или более раз обвиться вокруг опоры. Клетки, касающиеся опоры, слегка укорачиваются, а клетки противоположной стороны — удлиняются. Имеются данные, что в этой реакции принимает участие ауксин.
Рис. 25-4. Усики у смилакса (Smilax). Закручивание вызывается различной скоростью роста клеток внутренней и внешней сторон усика

Исследования, проведенные М. Джаффом из Лесного университета Уэйка, шт. Северная Каролина, обнаружили, что усики молодых растений гороха (Pisum sativum) могут накапливать сенсорную информацию, а спустя какое-то время восстанавливать ее и соответствующим образом реагировать. Например, если усики в течение трех дней держать в темноте, а затем раздражать, они не закрутятся, пока снова не будут выставлены на свет. Более того, их можно оставить в темноте на два часа после раздражения, но затем, снова оказавшись на свету, они немедленно закрутятся, т. е., хотя сенсорная информация сохранялась в темновой период, моторная функция в этих условиях не могла осуществиться. Почему так, до сих пор неясно. Высказано, однако, предположение, что АТР (который, по всей видимости, необходим для закручивания) может быть израсходован на протяжении темнового периода и восстанавливается только при освещении в результате фотосинтеза; возможно, кроме того, что в темноте накапливается ингибитор закручивания, который быстро исчезает при освещении.
Циркадианные ритмы
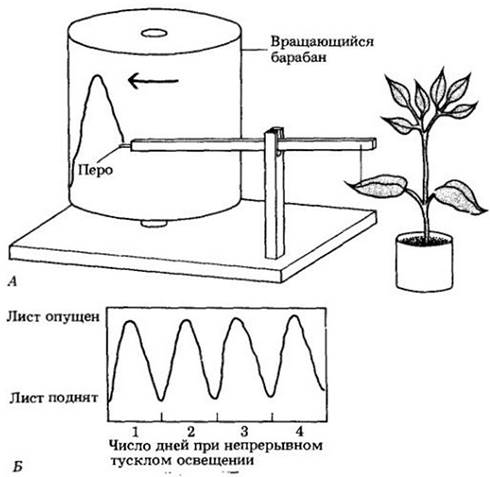
Общеизвестно, что некоторые растения открывают цветки утром и закрывают их в сумерки, расправляют листья при солнечном свете и опускают ночью (рис. 25-5). Уже в 1729 г. французский исследователь Жан-Жак де Меран заметил, что эти суточные движения продолжаются и при постоянном освещении растения слабым светом (рис. 25-6). Последующие исследования показали, что и процессы, не имеющие столь явных внешних проявлений, например, фотосинтез, образование ауксина и скорость клеточного деления, тоже имеют правильные суточные ритмы, сохраняющиеся при постоянных условиях среды. Эти регулярные, приблизительно 24-часовые (суточные), циклы называются циркадианными ритмами: от латинских слов circa — «приблизительно» и dies — «день». Циркадианные ритмы, обнаруженные почти у всех эукариот, по-видимому, отсутствуют у бактерий. Почему это так — неизвестно.
Рис. 25-5. Листья кислицы (Oxalis) в течение дня (А) и ночью (Б). Одна из гипотез состоит в том, что складывание листьев предохраняет их от поглощения лунного света ясными ночами и предотвращает тем самым возникновение фотопериодических реакций. Другая гипотеза, предложенная Дарвином сто лет назад, заключается в том, что складывание листьев защищает их ночью от охлаждения

Рис. 25-6. У многих растений листья располагаются перпендикулярно стеблю и солнечным лучам днем и параллельно стеблю ночью. Эти движения «засыпания» можно зарегистрировать на вращающемся барабане, используя тонко настроенную систему пера и рычага, прикрепленного к листу при помощи тонкой нитки (А). У многих растений, таких, как обыкновенная фасоль (Phaseolus vulgaris), показанная здесь, листья продолжают эти движения в течение нескольких дней даже при непрерывном тусклом освещении. К. Запись этого циркадианного ритма, показывающая его постоянство в условиях непрерывного тусклого освещения
Являются ли ритмы эндогенными?
Регулируются ли циркадианные ритмы эндогенно, т. е. внутренними механизмами или же вызываются внешними факторами? В течение ряда лет биологи спорили, не устанавливаются ли эти ритмы под действием таких факторов окружающей среды, как космические лучи, магнитное поле Земли или ее вращение. Можно отмстить два важных наблюдения, касающихся данного вопроса: во-первых, изученные ритмы — циркадианные (т. е. период их составляет около 24 ч); во-вторых, в их отношении имеются небольшие индивидуальные различия между организмами.
Попытки решить этот периодически возобновляющийся спор способствовали проведению многочисленных экспериментов в разнообразных условиях. Организмы помещались в соляные копи, отправлялись на кораблях на Южный полюс, облетали почти полсвета на самолетах и уже совсем недавно выводились на околоземную орбиту в спутниках. Хотя до сих пор раздаются голоса (их явное меньшинство) в пользу того, что циркадианные ритмы находятся под влиянием трудноуловимого геофизического фактора, большинство исследователей в настоящее время согласно с тем, что они эндогенные, т. е. регулируются внутренними факторами. Внутренний механизм отсчета времени называют биологическими часами организма.
Установка часов
При постоянных условиях среды период циркадианного ритма является свободнотекущим, т. е. его естественная продолжительность (обычно 21 — 27 ч) не должна устанавливаться заново для каждого цикла. Другими словами, циркадианный ритм ведет себя как саморегулирующийся осциллятор. Хотя этот ритм возникает в самих организмах, окружающая среда действует как синхронизирующий агент (время- задатель). Действительно, она ответственна за то, чтобы циркадианный ритм поспевал за суточным 24-часовым циклом света и темноты. Если период циркадианного ритма растения будет больше или меньше суток, ритм скоро перестанет согласовываться с циклом смены дня и ночи. Затем, если бы в соответствии с таким ритмом происходило цветение (обычно наблюдаемое в светлый период), оно приходилось бы каждый день на разнос время, включая и темный период. Поэтому растению необходимо ресинхронизироваться, т. е. «захватываться» ритмом 24-часовых суток.
«Захватывание» — это процесс, благодаря которому периодическое повторение света и темноты или некоторого внешнего цикла синхронизирует циркадианный ритм с цикличностью внешнего фактора. Циклы света и темноты и температурные циклы считаются основными факторами захватывания (рис. 25-7).
Рис. 25-7. А. Микрофотография динофлагелляты Gonyaulax polyedra, одноклеточной морской водоросли. Б. У G. polyedra три разные функции осуществляются при разных циркадианных ритмах: биолюминесценция, которая достигает максимума в середине ночи (цветная кривая); фотосинтез, достигающий максимума в середине дня (черная кривая); деление клеток (не поймано), копюрое происходит в предрассветные часы. Если Gonyaulax культивируется при непрерывном тусклом освещении, эти три функции продолжают осуществляться с тем же ритмом в течение многих дней и даже недель после того, как пройдет ряд клеточных делений. В. Ритм биолюминесценции у Gonyaulax, подобно большинству циркадианных ритмов, может варьировать при изменении циклов освещения. Например, если исследователи подвергают культуры водоросли воздействию чередующихся периодов света и темноты продолжительностью 6 ч каждый, ритмическая функция приобретает такую же цикличность (слева). Если затем культуры будут помещены в условия непрерывного тусклого освещения, организмы вернутся к своему первоначальному ритму с цикличностью около 24 ч
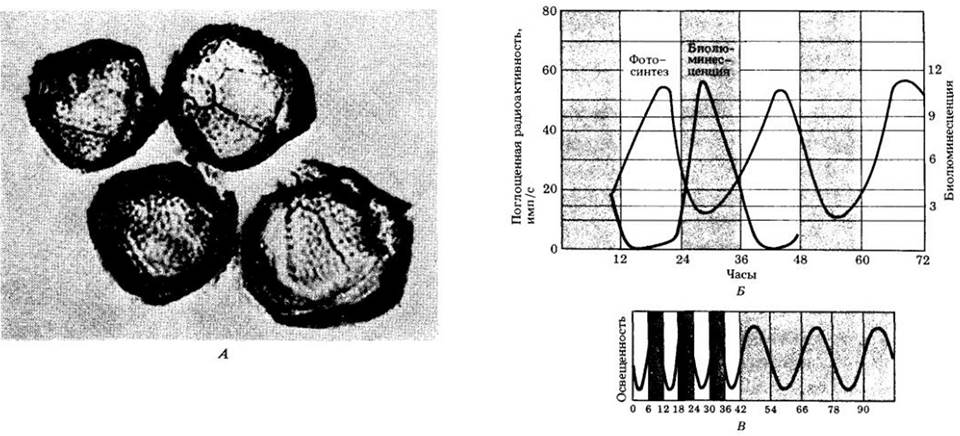
Другой интересной чертой является то, что циркадианные ритмы не ускоряются автоматически при подъеме температуры. Причина, по которой можно ожидать ускорения, заключается в том, что биохимические процессы (а внутренние биологические часы, несомненно, должны иметь биохимическую основу) протекают быстрее при высоких температурах, чем при низких. Одни часы с повышением температуры идут несколько быстрее, зато другие — более медленно, а у многих из них ход почти не меняется. Таким образом, они должны иметь компенсаторный механизм — систему обратной связи, позволяющую им приспосабливаться к температурным изменениям. Такая температурно-компенсаторная система была бы очень полезна растениям.
Некоторые последние данные наводят на мысль о том, что ключом к биологическим часам могут служить клеточные мембраны. Хотя механизм еще неясен, вероятно, что к его работе имеют отношения такие известные функции мембран, как регуляция транспорта ионов, поступающих в клетки и выходящих из них и субклеточных компартментов, а также регуляция энергетического метаболизма на уровне преобразующих энергию мембран хлоропластов и митохондрий.
Некоторые биологические ритмы регулируют взаимоотношения между организмами. Например, некоторые растения выделяют нектар в определенное время дня. В результате пчелы, имеющие свои собственные биологические часы, привыкают посещать цветки именно в это время, получая тем самым максимум вознаграждения для себя и обеспечивая перекрестное опыление цветков.
Однако для большинства организмов использование биологических часов для таких специальных целей имеет, невидимому, второстепенное значение. В первую очередь физиологические часы дают возможность растениям или животным реагировать на смену сезонов в результате точного измерения меняющейся продолжительности светового дня. Таким образом, непостоянство окружающей среды вызывает ответную реакцию, которая проявляется в регуляции роста, репродукции и других процессов.
Фотопериодизм
65 лет тому назад среди растений табака (Nicotiana tabacum) на поле вблизи Вашингтона, округ Колумбия, появился мутант. Новые растения достигали в высоту более 3 м и имели необычно крупные листья. По мере наступления соответствующего сезона обычные растения цвели, а экземпляры Мэриленд Маммот1 (так стали называть эту разновидность) только увеличивались в размерах. Два исследователя из Сельскохозяйственного департамента США У. Гарнер и X. Аллард поместили черенки Мэриленд Маммот в теплицу, где они были защищены от мороза. Черенки зацвели в декабре, хотя при этом достигли в высоту только 1,5 м, т. е. оказались вдвое короче, чем родительские экземпляры. Новые растения, выросшие из их семян, тоже не цвели почти до самой зимы.
1В переводе с английского «Морилендский мамонт». — Прим. ред.
Случилось так, что в это же время Гарнер и Аллард проводили эксперименты с разновидностью сои (Glycine mах) Билокси. Агрономы стремились получить ее урожаи в разное время, высевая семена с двухнедельными интервалами с раннего мая до конца июня. Однако эти опыты не дали желаемых результатов; все растения назависимо от того, когда были высеяны семена, зацвели в одно и то же время — в сентябре.
Исследователи приступили к выращиванию табака Мэриленд Маммот и сои Билокси в разнообразных контролируемых условиях температуры, влажности, питания и света. В результате проведенных экспериментов они обнаружили, что критическим фактором для обоих видов является длина светового дня. Ни одно растение не цвело, если она превышала некоторое критическое число часов. Таким образом, соя независимо от того, когда высевалась, зацветала только тогда, когда дни становились достаточно короткими, т. е. в сентябре, а Мэриленд Маммот независимо от того, насколько высокими были его растения, не цвел до декабря, когда дни становились еще короче.
Гарнер и Аллард назвали это явление фотопериодизмом. Фотопериодизм — это биологическая реакция на изменения освещенности, происходящие в 24-часовом суточном цикле. Хотя концепция фотопериодизма сформировалась на основе изучения растений, в настоящее время она нашла подтверждение и в других областях биологии, например, при исследовании брачного поведения яблонной плодожорки, еловой листовертки-почкоеда, тлей, выемчатокрылой моли, рыб, птиц и млекопитающих.
Длиннодневные и короткодневные растения
Гарнер и Аллард провели эксперименты и подтвердили свое открытие на многих видах растений. После этого они смогли ответить на множество вопросов, долго волновавших как профессиональных ботаников, так и садоводов-любителей. Почему, например, нет амброзии (Ambrosia) на севере шт. Мэн? Ответ: потому что она начинает цвести, когда продолжительность дневного освещения составляет 14,5 ч или менее. Между тем длинные летние дни укорачиваются до 14,5 ч на севере шт. Мэн только в августе, а тогда семена амброзии уже не успевают созреть до морозов. Вопрос: почему шпинат (Spinacea olerácea) не растет в тропиках? Ответ: потому что для цветения он нуждается по крайней мере в 14 ч ежедневного освещения в течение хотя бы двух недель, а эти условия для тропиков не характерны.
Исследователи обнаружили, что все растения можно разделить на три основные группы: короткодневные, длиннодневные и нейтральные к длине дня. Первые цветут ранней весной или осенью и нуждаются для этого в более короткой длине дня, чем некая критическая для них. Например, дурнишник обыкновенный (Xanthium strumarium) побуждается к цветению 16-часовым или более коротким днем (рис. 25-8 и 25-9). Среди других короткодневных растений можно назвать некоторые виды хризантем (рис. 25-10, А), пуансеттию, землянику и первоцветы.
Рис. 25-8. Относительная длина дня и ночи определяет время цветения растений. Четыре кривые изображают годовые изменения длины дня в четырех городах Северной Америки, находящихся на четырех различных широтах. Горизонтальные цветные линии отражают эффективный фотопериод трех различных короткодневных растений. Дурнишник, например, цветет, если продолжительность светового дня составляет 16 или менее часов. В Майами он может цвести довольно рано, но в Виннипеге бутоны появляются только в начале августа, т. е. так поздно, что мороз может убить растения до того, как созреют семена
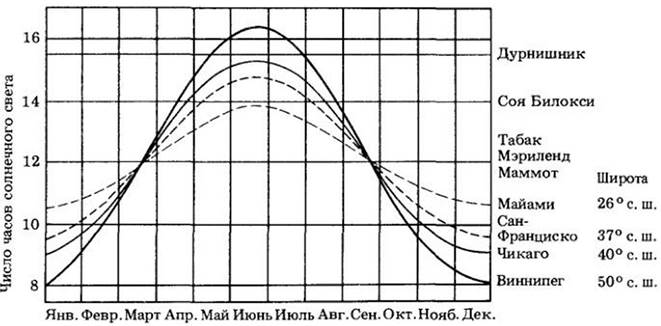
Рис. 25-9. Короткодневные растения зацветают при длине светового дня, меньшей критической. Обыкновенный дурнишник (Xanthium strumarium) цветет, если длина светового периода составляет менее 16 ч. Белене (Hyoscyamus niger), чтобы зацвести, необходимо около 10 ч света (в зависимости от температуры) или более. Однако, если темный период прерывается вспышкой света, белена будет цвести даже при коротком дне. «Импульс» света во время темного периода оказывает противоположное влияние на короткодневные растения — он препятствует цветению. Диаграммы наверху показывают продолжительность периодов света и темноты в течение суток
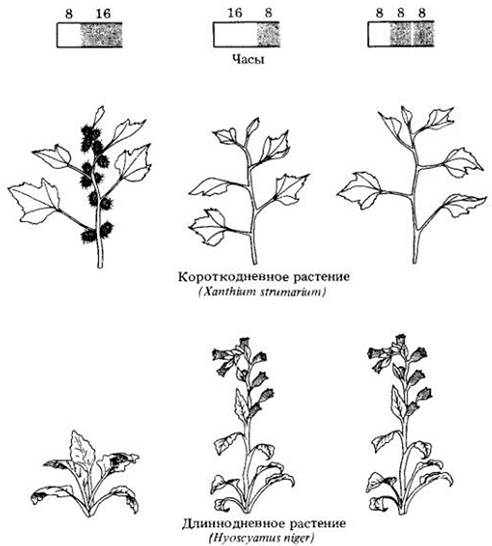
Длиннодневные растения цветут главным образом летом и нуждаются для этого в большей, чем некая критическая для них, длине дня. Примерами могут служить шпинат, некоторые сорта картофеля, отдельные разновидности пшеницы, латук и белена (Hyoscyamus niger) (рис. 25-9 и 25-10, Б).
Рис. 25-10. Представители короткодневных и длиннодневных растений, из которых каждое выращивалось в условиях короткого дня (слева) или длинного (справа) дня. A. Chrysanthemum — короткодневное растение. Б. Шпинат (Spinacia oleracea) — длиннодневное растение. Заметьте, что растения, находившиеся в условиях длинного дня, имеют более высокие стебли, чем растения, находившиеся в условиях короткого дня

И дурнишник, и шпинат будут цвести при длине дня 14 ч, однако лишь второй из них — длиннодневное растение. Важна, как уже говорилось, не абсолютная длина фотопериода, но и то, будет ли он длиннее или короче, чем некий критический. Нейтральные к длине дня растения цветут вне зависимости от нес. Примерами могут служить огурец, подсолнечник, табак, рис, кукуруза, горох огородный.
Среди отдельных видов растений, широко распространенных от севера до юга, часто наблюдаются различные фотопериодические экотипы, адаптированные к местным условиям. Так, у многих степных злаков, встречающихся от южной Канады до Техаса, северные экотипы цветут раньше южных, если они выращиваются вместе в одинаковых условиях. Различные популяции очень хорошо приспособлены к фотопериодическому режиму среды обитания.
Фотопериодическая реакция может быть удивительно точной. Так, при 22,5°С длиннодневная белена будет цвести, если фотопериод составит 10 ч 20 мин (рис. 25-9), а при 10- часовом фотопериоде при этой же температуре цвести уже не будет. Кроме того, условия среды тоже влияют на фотопериодические реакции. Например, при 28,5°С для цветения белены требуется не менее 11,5 ч света, в то время как при 15,5°С — лишь 8,5.
У различных видов реакция может быть неодинаковой. Некоторым растениям достаточно одноразового воздействия цикла «день—ночь», в то время как другие (например, шпинат) нуждаются в нескольких неделях экспозиции. У многих растений наблюдается корреляция между числом индукционных циклов и скоростью зацветания или числом сформированных цветков. Некоторые растения для индукции цветения должны достигнуть определенной степени зрелости, в то время как другие будут реагировать на соответствующий фотопериод уже на стадии проростков. Ряд видов по мере старения зацветает в конце концов даже без воздействия, соответствующего фотопериода, хотя при надлежащей экспозиции они зацвели бы значительно раньше.
Измерение периода темноты
В 1938 г. другие два исследователя, Карл Хамнер и Джеймс Боннер, занялись изучением фотопериодизма, используя дурнишник в качестве экспериментального объекта. Как упоминалось ранее, дурнишник является короткодневным растением, требующим для цветения не более 16 ч света в 24- часовом цикле. Это растение особенно удобно для экспериментальных целей, поскольку в лабораторных условиях уже одноразовое воздействие короткодневного цикла вызывает через две недели зацветание, даже если растение будет немедленно возвращено в условия длинного дня. Дурнишник может переносить и грубую хирургическую обработку, например, выживет даже в том случае, если будут удалены все его листья. Хамнер и Боннер показали, что фотопериод у дурнишника воспринимается пластинкой листа. У лишенного листьев растения нельзя индуцировать цветение. Однако, если на стебле оставлена хотя бы одна восьмая часть полностью развившегося листа, единственная коротко дневная экспозиция сможет вызвать цветение. В ходе этих исследований, проводимых при различных экспериментальных условиях, Хамнер и Боннер сделали решающее и совершенно неожиданное открытие. Если период темноты прерывается светом от лампочки в 25 Вт хотя бы на одну минуту, цветение не происходит. Прерывание же светового периода темнотой совершенно не сказывается на цветении. Соответствующие эксперименты с другими короткодневными растениями показали, что и они нуждаются скорее в периодах непрерывной темноты, чем непрерывного света.
При этом существенно, прерывается ли ночной период в середине или нет. Если короткодневное растение, такое, как дурнишник, освещают в течение 8 ч, а затем помещают на продолжительное время в темноту, то растение переходит от состояния возрастающей чувствительности к световому перерыву (в течение примерно 8 ч) к состоянию, при котором световой перерыв имеет все меньшее значение. Действительно, одноминутная световая экспозиция после 16 ч темноты стимулирует цветение. Согласно гипотезе немецкого физиолога растений Эрвина Бюннинга, фотопериодический контроль цветения регулируется и эндогенным ритмом, причем фазы ритма различаются в зависимости от того, оказывает ли свет стимулирующее (фотофильная фаза) или ингибирующее (скотофильная фаза) действие. В связи с этим изменения в чувствительности соответствуют различным фазам биологических часов. Эксперименты по так называемому световому перерыву дают одно из лучших доказательств гипотезы Бюннинга.
На основании открытия Гарнера и Алларда садоводы, выращивающие для коммерческих целей хризантемы, сделали вывод, что можно задерживать цветение короткодневных растений, увеличивая дневное освещение при помощи искусственного света. Исходя из новых экспериментов Хамнера и Боннера, они стали задерживать цветение, включая свет на короткое время в середине ночи.
Что можно сказать относительно длиннодневных растений? Они тоже измеряют продолжительность темнового периода. Длиннодневное растение будет цвести, если находится в лаборатории в условиях 16-часового освещения и 8 ч темноты; оно будет цвести и получая 8 ч света и 16 ч темноты, если период темноты хотя бы на короткое время будет прерываться светом.
Химическая основа фотопериодизма
Следующий важный подход к изучению фотопериодических реакций растений был разработан группой ученых научно- исследовательской станции Сельскохозяйственного департамента США в Белтсвилле, Мэриленд. Это направление развилось на основе более ранних исследований, выполненных на семенах латука (Lactuca sativa). Семена его прорастают только в том случае, если их выставляют на свет. Это требование относится ко многим мелким семенам, нуждающимся для прорастания в рыхлой почве и неглубоком посеве. Более ранние работы по изучению потребностей в свете прорастающих семян латука показали, что красный свет стимулирует прорастание, а свет с несколько большей длиной волны (дальний красный) задерживает прорастание даже более эффективно, чем отсутствие освещения.
Хамнер и Боннер обнаружили, что, если темный период прервать единственной вспышкой света от обычной лампочки, дурнишник цвести не будет. Исходя из этого, исследователи из Белтсвилла начали эксперименты со светом различной длины волны, варьируя интенсивность и длительность вспышки. Они обнаружили, что красный свет с длиной волны около 660 нм (оранжево-красный) наиболее эффективен для предотвращения цветения дурнишника и других короткодневных растений. Они показали также, что такой свет наиболее эффективно стимулирует цветение длиннодневных растений.
Белтсвиллская группа обнаружила, что в том случае, когда за вспышкой красного света следует вспышка дальнего красного света, семена латука не прорастают. Красный свет длиной волны 660 нм, наиболее эффективный для индукции прорастания семян, вызывал и цветение. Более того, они нашли, что свет, наиболее эффективно противодействующий красному, имеет длину волны 730 нм. Чередующиеся вспышки красного и дальнего красного света могут повторяться снова и снова; число вспышек не имеет значения, но имеет значение природа последней вспышки. Если ряд вспышек заканчивается вспышкой красного света, большинство семян прорастает, если же — дальнего красного, то большинство семян не прорастает (рис. 25-11).
Рис. 25-11. Свет и прорастание семян латука. А. Семена, выставленные на короткое время на красный свет. Б. Семена, выставленные на красный свет, затем на дальний красный свет. В. Семена, выставленные последовательно на красный, дальний красный, красный свет. Г. Семена, выставленные последовательно на красный, дальний красный, красный, дальний красный свет. Так или иначе, семена прорастали в зависимости от конечной длины волны в серии экспозиций — красного света, способствующего прорастанию, и дальнего красного, подавляющего его

Дальний красный свет, испытанный на короткодневных и длиннодневных растениях, показал одинаковое «выключающее» действие. Если во время темпового периода давался только дальний красный свет, он был неэффективен. Однако вспышка дальнего красного света непосредственно вслед за вспышкой красного снимала его эффект.
Открытие фитохрома
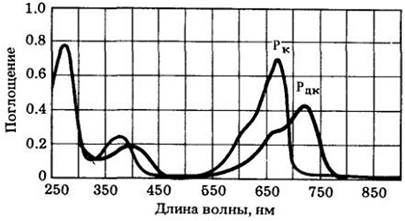
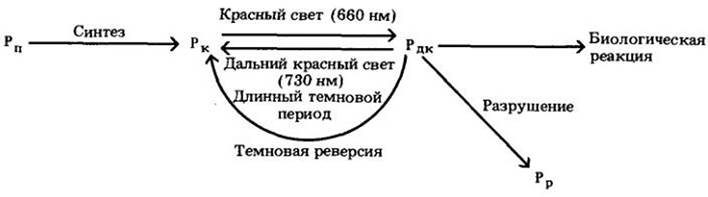
Растения содержат пигмент, существующий в двух различных взаимопревращающихся формах: Рк (форма, которая поглощает красный свет) и Рдк (форма, поглощающая дальний красный свет). Когда молекула Рк поглощает фотон красного света с длиной волны 660 нм, она превращается за несколько миллисекунд в Рдк; когда молекула Рдк поглощает фотон дальнего красного света с длиной волны 730 нм, она очень быстро превращается обратно в форму Рк. Эти реакции называются фотоконверсиями. Форма Рдк биологически активна (т. е. она обеспечивает реакцию, такую, как прорастание семян), в то время как форма Рк неактивна. Таким образом, молекула пигмента может функционировать как «биологический переключатель» и направлять ход реакции в 17 или другую сторону в зависимости от того, в какой форме находится.
С этой точки зрения легко интерпретировать результаты экспериментов по прорастанию семян латука. Поскольку Рк поглощает красный свет более эффективно (рис. 25-12), то при этой длине волны большая часть молекул переходит в форму Рдк, индуцируя тем самым прорастание. Затем дальний красный свет, поглощаемый формой Рдк, побуждает по существу все молекулы к обратному переходу, снимая тем самым эффект красного света.
Рис. 25-12. Спектры поглощения двух форм фитохрома Рк и Рдк. Это различие в спектре поглощения позволило выделить пигмент
Что можно сказать о цветении при естественном цикле день — ночь? Поскольку в белом свете представлены различные длины волн (как красные, так и дальние красные), обе формы пигмента подвергаются одновременному действию фотона, достаточного для их фотоконверсии в противоположные формы. Через несколько минут на свету устанавливается фоторавновесие, т. е. прямая (Рк —> Рдк) и обратная (Рдк —> Рк) реакции сбалансированы. На данный момент времени в растительной клетке на дневном свету существует постоянное соотношение фитохромов (около 60% Рдк и 40% Рк), и это соотношение сохраняется, пока продолжается действие света.
Если растение в конце светового периода поместить в темноту, уровень Рдк в течение нескольких часов постоянно уменьшается. Если высокий уровень Рдк восстанавливается кратковременным освещением красным светом в середине темнового периода (см. рис. 25-9), он будет задерживать цветение коротко дневных (так называемых «длинноночных») растений, которые в другом случае стали бы цвести, и будет способствовать цветению длиннодневных (так называемых «коротконочных») растений, которые в другом случае не зацвели бы. В любом случае эффект красного импульса на восстановление высокого уровня Рдк может сниматься немедленным коротким облучением дальним красным, который переводит Рдк снова в Рк.
В 1959 г. Гарри Бортуик и его сотрудники из Белтсвилла назвали этот пигмент фитохромом и представили убедительное физическое доказательство его существования. Основные характеристики пигмента схематически показаны на рис. 25-13. Молекула синтезируется непрерывно как Рк и накапливается в этой форме в растущих в темноте растениях. Свет вызывает фотопревращение Рк в Рдк, который индуцирует биологическую реакцию. Рдк может превращаться в Рк путем фотоконверсии, поглощая дальний красный свет, или в темноте (этот процесс назван «темновая реверсия»; он длится от нескольких минут до нескольких часов). Рдк, по- видимому, может разрушаться в результате гидролиза протеазами — так называемой деструкции, которая происходит в течение нескольких часов. Все три альтернативных пути удаления Рдк обеспечивают возможность отмены индуцированной реакции. Следует отметить, однако, что темновой вариант отмены обнаружен лишь у двудольных растений, но не у однодольных.
Рис. 25-13. Фитохром был сначала синтезирован из аминокислот в форме Рк (обозначение Рn — для предшественника). Рк превращается в Рдк при освещении красным светом. является активной формой, которая вызывает биологическую реакцию. Рдк превращается снова в Рк, если подвергается действию дальнего красного света. В темноте Рдкпревращается в Рк или разрушается (обозначение Рр — для продукта разрушения)
Выделение фитохрома
По сравнению с таким пигментом, как хлорофилл, фитохром присутствует в растениях в малых количествах. Для идентификации фитохрома необходим спектрофотометр, который чувствителен к крайне малым изменениям в поглощении света. Такой спектрофотометр был создан только через семь лет после того, как предположили существование фитохрома; впервые новый прибор и был применен для обнаружения и выделения этого пигмента.
Для того чтобы избежать ошибок, вызванных присутствием хлорофилла, который, подобно фитохрому, поглощает свет с длиной волны около 660 нм, в качестве источника фитохрома использовали растущие в темноте проростки (в которых хлорофилл еще не успел образоваться). Выделенный пигмент имел синий цвет и переходил из одной формы в другую под действием красного и дальнего красного света; при этом наблюдались небольшие обратимые изменения его окраски в ответ на поглощение света соответствующей длины волны.
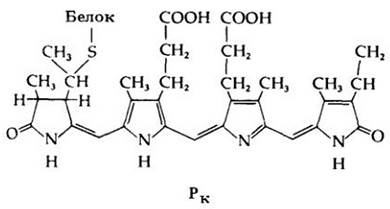
Было показано, что молекула фитохрома состоит из двух различных частей: хромофора (поглощающего свет) и большого белкового участка (рис. 25-14). Хромофор очень напоминает фикобилин — вспомогательный пигмент цианобактерий и красных водорослей.
Рис. 25-14. Хромофор фитохрома (формы Рк); видна его связь с белковой частью молекулы
Специфический механизм работы фитохрома пока еще не установлен. Однако ясно, что фитохром регулирует морфогенез, изменяя транскрипцию генов.
Другие реакции, опосредованные фитохромом
Показано, что фитохром включается и в ряд других реакций растения. Прорастание многих семян, например, происходит в темноте. У проростков стебель удлиняется быстро, проталкивая побег (или, у большинства однодольных, семядолю) через почву. На данной стадии роста не происходит существенного развития листьев, поскольку это могло бы препятствовать прорастанию побега. Нельзя сказать, что почва необходима для подобного типа роста; любой проросток в темноте будет вытягиваться, становясь длинным и тонким и развивая мелкие листья. Он будет желтым или бесцветным, поскольку пластиды не зеленеют, пока проросток не выходит на свет. Такой проросток называют этиолированным (рис. 25-15).
Рис. 25-15. Выращенные в темноте проростки (слева) — тонкие и бледные; они имеют более длинные междоузлия и более мелкие листья, нем нормальные проростки (справа). Этот комплекс физических признаков, называемый этиоляцией, имеет немаловажное значение для выживания проростков, поскольку благодаря ему увеличиваются шансы проростков выйти на свет прежде, чем будут использованы запасенные ими энергетические ресурсы

Когда верхушка проростка появляется над землей, этиолированный рост уступает место нормальному. У двудольных изогнутый в виде петли гипокотиль выпрямляется, скорость роста стебля может уменьшаться и начинается рост листьев (см. рис. 19-2). У злаков рост мезокотиля (часть зародышевой оси между щитком и колеоптилем) останавливается, стебель удлиняется и листья разворачиваются.
Проростки бобов, растущие в темноте и получающие в день, например, пять минут красного света, реагируют на этот свет начиная с четвертого дня. Если после красного света проростки пять минут выдержать при дальнем красном, никаких изменений, обычно происходящих при красном свете, нс наблюдается (рис. 25-16). Подобным же образом освещение красным светом проростков злаков служит сигналом для окончания роста мезокотиля, а действие красного света погашается дальним красным светом.
Рис. 25-16. Все три растения фасоли освещались каждый день на протяжении 8 ч. Растение, расположенное в центре, выставлялось в начале каждого темпового периода на 5 мин на дальний красный свет, способствующий росту в длину. Растение справа освещалось в течение 5 мин дальний красным светом, а затем — красным, который нейтрализовал действие дальнего красного. Растение слева служило контролем; его фитохром находится в форме Рдк, поскольку растение вступает в темновой период после экспозиции на дневном свету

Недавно в одной из работ, опубликованной в Англии, высказано предположение о еще одной функции фитохрома у растений, произрастающих в естественных условиях; он может выступать как детектор, улавливающий затенение другими растениями. Излучение с длиной волны менее 700 нм почти полностью отражается или поглощается растениями, в то время как излучение с длиной волны между 700 и 800 нм (в дальнем красном диапазоне) в значительной степени пропускается. Это вызывает критический сдвиг вверх отношения Рк к Рдк (т. е. больше Рдк переходит в Рк) у затененных растений и приводит к быстрому увеличению скорости роста междоузлий.
Обратимые реакции на красный/дальний красный свет имеют отношение и к образованию антоциана у яблок, брюквы и капусты; к прорастанию семян; к изменению хлоропластов и остальных пластид, а также к огромному разнообразию других реакций растения во всех фазах жизненного цикла.
Фитохром и фотопериодизм
Когда впервые было показано существование фитохрома, открывшие его исследователи высказали гипотезу, что поведение фитохрома может объяснить явление фотопериодизма — т. е. что обратимые реакции на красный/дальний красный свет могут включаться в механизм измерения времени, или в биологические часы. Согласно этой гипотезе, Рдк задерживает цветение у короткодневных растений, но ускоряет цветение длиннодневных растений. У короткодневных растений Рдк будет накапливаться на свету и удаляться в последующий темный период путем разрушения или темновой реверсии. Если ночи были достаточно длинными, весь
(или критическое количество) Рдк будет удален и цветение не будет больше тормозиться. С другой стороны, длиннодневным растениям требуются короткие ночи, в течение которых не произойдет полного разрушения Рдк; если ночь была достаточно короткой, к ее концу останется достаточно Рдк, чтобы ускорить цветение.
Эксперименты, однако, показали, что Рдк исчезает у многих растений в течение трех или четырех часов темноты. На основании этих экспериментов и других наблюдений в настоящее время пришли к общему мнению, что явление фотопериодизма, связанное с измерением времени, не контролируется одними лишь взаимопревращениями Рк и Рдк. Следует искать более сложное объяснение.
Гормональный контроль цветения
Хамнер и Боннер в своих ранних опытах с дурнишником показали, что бутон стимулируется к цветению листом, «воспринимающим» свет. По-видимому, какое-то вещество, оказывающее решающее влияние на рост и развитие, перемещается от листа к бутону. Это гипотетическое вещество было названо гормоном, или стимулом, цветения.
Эксперименты, подтверждающие существование стимула цветения, были независимо друг от друга проведены в нескольких лабораториях в 1930-е гг. Наиболее убедительными оказались опыты, проведенные в числе первых советским физиологом растений М. X. Чайлахяном за несколько лет до начала работ с дурнишником. Используя короткодневное растение Chrysanthemum indicum, Чайлахян показал, что, если верхнюю часть растения лишить листьев, а листья, оставшиеся на нижней его части, поместить в условия короткого дня, растение зацветет. Напротив, если верхнюю, лишенную листьев часть стебля поместить в условия короткого дня, а нижнюю облиственную — в условия длинного, цветения нс произойдет. На основании полученных результатов Чайлахян предположил, что в листьях образуется гормон, который перемещается к верхушке стебля и вызывает цветение. Чайлахян назвал этот гипотетический гормон флоригеном.
Дальнейшие эксперименты показали, что цветения нс происходит, если лист удаляется сразу же после фотоиндукции. Но, если лист оставить на растении в течение нескольких часов после окончания индукционного цикла, последующее удаление листа не оказывает никакого влияния на цветение. Гормон цветения может передаваться от фотоиндуцированного растения к неиндуцированному через прививку. В отличие от ауксина, который может проходить через агар или другое неживое вещество, флориген перемещается из одной растительной ткани в другую только в том случае, если между ними имеются анатомические связи. Если ветвь окольцована, т. е. удалена кольцевая полоска коры, передвижение флоригена прекращается. На основании этих данных было сделано заключение, что флориген передвигается по флоэме, т. е. тем путем, которым осуществляется транспорт большинства органических веществ в растениях.
Впоследствии Антон Ланг из Калифорнийского технологического института показал, что у некоторых длиннодневных растений и двулетников, таких, как сельдерей и капуста, можно вызвать цветение, обрабатывая их гиббереллином, даже в тех случаях, когда растения выращиваются не при соответствующем фотопериоде. Это открытие заставило Чайлахяна изменить его флоригенную гипотезу и высказать предположение, что в действительности флориген состоит из двух гормонов, гиббереллина и пока еще неидентифицированного антезина. Согласно этой гипотезе, длиннодневные растения в течение неиндукционного фотопериода вырабатывают антезин, но не гиббереллин и обработка гиббереллином в это время может вызвать цветение. С другой стороны, короткодневные растения образуют гиббереллин, но не антезин, если растут в неиндукционных условиях. Хотя концепция сложной природы флоригена стимулировала многие исследования, она, к сожалению, не смогла ответить на такой, например, принципиально важный вопрос: почему короткодневные растения, растущие в неиндукционных условиях, не вызывают цветения привитых длиннодневных растений, которые тоже не подвергались воздействию индуцирующего фотопериодического сигнала.
У некоторых растений — примером может служить соя Билокси (Glycine max) — листья с растения-привоя должны быть удалены, иначе оно не зацветет. Это наблюдение наводит на мысль, что в листьях неиндуцированных растений может образовываться ингибитор. Действительно, основываясь на подобных данных, некоторые исследователи пришли к заключению, что вещества, вызывающего цветение, не существует; скорее можно говорить о веществе, которое, если оно не удалено, задерживает цветение. Сейчас имеются серьезные основания говорить о том, что, по крайней мере у некоторых растений, в контроль цветения включаются вещества как тормозящие процесс, так и ускоряющие его.
Самое убедительное доказательство существования этих веществ в одном растении получено благодаря экспериментальным исследованиям Антона Ланга в Университете шт. Мичиган, а также М. X. Чайлахяна и И. А. Фроловой в Московском институте физиологии растений им. К. А. Тимирязева. Эти ученые избрали для своих исследований три сорта растений табака: нейтральный к длине дня Nicotiana tabacum сорта Трабезонд, короткодневный табак сорта Мэриленд Маммот и длиннодневный Nicotiana silvestris. Обнаружилось, что формирование цветков у табака, нейтрального к длине дня, ускоряется путем прививки на него длиннодневного растения, если привитые растения помещаются в условия длинного дня. Аналогичные результаты получали и в том случае, когда на нейтральные к длине дня растения прививались короткодневные и привои помещались в условия короткого дня. Если длиннодневные растения, привитые на растения, нейтральные к длине дня, выставлялись на короткий день, цветение нейтрального к длине дня подвоя в значительной степени тормозилось (рис. 25-17). Напротив, если короткодневные растения, привитые на растения, нейтральные к длине дня, выставлялись на длинный день, задержки в цветении не наблюдалось, или она была небольшой.
Рис. 25-17. Прививки Nicotiana silvestris (длиннодневного растения) на Nicotiana tabacum сорта Трабезонд (нейтрального к длине дня растения). В условиях длинного дня развитие цветков у нейтрального к длине дня табака ускоряется за счет прививки (растение слева). В условиях короткого дня нейтральный к длине дня табак остается в вегетативном состоянии в течение всего (от 90 до 94 дней) эксперимента (растение справа)

Эти результаты свидетельствуют о том, что в листьях длиннодневных растений в условиях длинного дня образуются вещества, вызывающие цветение, а в условиях короткого дня вещества, тормозящие цветение, и, кроме того, оба типа веществ могут переходить из привоя в подвой и транспортироваться по растению. В листьях короткодневного растения табака в условиях длинного дня, по-видимому, образуется мало веществ, задерживающих цветение, или не образуется вообще. Ингибиторы цветения короткодневных растений гораздо менее эффективно задерживают цветение, чем образующиеся у длиннодневных растений.
Доказательство существования ингибиторов и стимуляторов цветения вполне убедительно, хотя все попытки выделить эти вещества пока оказались безуспешными.
Покой
Растения не растут постоянно с одинаковой скоростью. В неблагоприятное время года они ограничивают свой рост или вообще прекращают его. Эта способность позволяет растениям пережить неблагоприятные периоды, связанные с недостатком воды или низкой температурой.
Покой — это особое состояние задержки роста, характерное как для семян, так и для почек. Не прорастающее семя, несмотря на то что оно в достаточной степени обеспечено водой, кислородом и находится в благоприятных температурных условиях, называется покоящимся. Покой почек определяется как прекращение видимого роста. Только некоторые «сигналы» из окружающей среды, часто вполне определенные, могут «активировать» покоящуюся почку или зародыш. Эта адаптация имеет жизненно важное значение для растения. Например, весной почки растений раскрываются, формируются цветки, прорастают семена — однако каким образом растения узнают, что наступила весна? Если достаточно одной только теплой погоды, то в некоторые годы во время бабьего лета все растения зацвели бы, а проростки тронулись в рост только для того, чтобы затем быть убитыми зимним морозом. То же самое произошло бы и в оттепели, которые часто прерывают зимний сезон. Покоящиеся семя или почка не реагируют на эти на первый взгляд благоприятные условия благодаря эндогенным ингибиторам, которые должны быть удалены или нейтрализованы до окончания периода покоя. В противоположность такому «нежеланию» прорастать слишком быстро некоторые семена могут прорастать сразу же после образования. Например, это относится к иве (Salix) и сахарному клену (Acer saccharum), чьи семена теряют жизнеспособность уже в течение недели, если находятся на воздухе. У мангровых растений (Rhizophora mangle) семена прорастают прямо на родительском растении. Они образуют тяжелые корешки и падают, как дротик, погружаясь ими в ил (рис. 25-18). Для практики сельского хозяйства семена специально отбирают на основе их способности быстро прорастать в благоприятных условиях; подобная особенность была бы очень опасной для семян дикорастущих видов.
Рис. 25-18. Прорастающее семя (А) и молодой сеянец (Б) мангрового растения Rhizophora mangle. Семена прорастают еще на родительском растении и образуют дротиковидный корень, погружающийся в ил, как только прорастающее семя падает вниз

Покой семян
Семена почти всех растений, обнаруженных в районах с выраженными сезонными изменениями температуры, нуждаются для прорастания в холодном периоде, который устанавливается в нормальных условиях с наступлением зимы. Семена многих декоративных растений тоже нуждаются в периоде низких температур. Если влажное семя обработать низкими температурами в течение нескольких дней (средний оптимум температуры и времени — 5°С в течение 100 дней), покой может быть прерван и семя прорастет. Этот садоводческий прием называется стратификацией. Многие семена для прорастания нуждаются в высушивании (хотя некоторые из них могут до высушивания находиться в непокоящемся состоянии). Эта потребность препятствует их прорастанию во влажной среде плода родительского растения. Одни семена, например, латука, для прорастания требуют света, а прорастание других, наоборот, светом ингибируется.
Некоторые семена не прорастают в естественных условиях до тех пор, пока не растрескаются под действием почвы. Такое растрескивание способствует сбрасыванию семенной кожуры, что облегчает поступление в семя воды и кислорода, а в некоторых случаях и удаление ингибиторов. Твердая семенная кожура, препятствующая поглощению воды и росту зародыша, часто встречается у бобовых.
Семена некоторых видов пустынных растений прорастают только в тех случаях, когда выпадает достаточное количество осадков для вымывания тормозящих прорастание веществ, находящихся в семенной кожуре. Кроме того, и для укоренения проростков необходимо определенное количество воды, которое должно быть запасено растением. Механическое разрушение семенной кожуры (скарификация) ножом, напильником или наждачной бумагой способствует нарушению состояния «твердосемянности» или удалению ингибитора, или, наконец, восстановлению метаболитической активности, необходимой для прорастания. Прорастание может быть вызвано и погружением семян в спирт и другие растворяющие жир жидкости (чтобы растворить восковые вещества, препятствующие поступлению в семена воды) или в концентрированные кислоты. Эти приемы широко используются в садоводстве.
Некоторые семена в покоящемся состоянии могут долгое время (годы, десятилетия и даже века) оставаться жизнеспособными. В 1879 г. семена 20 видов обычных мичиганских сорных трав были помещены У. Билом из Университета шт. Мичиган во влажный, хорошо аэрируемый песок для того, чтобы определить их долговечность. Через 100 лет семена трех видов были еще жизнеспособны. Недавно обнаружены жизнеспособные семена (семи из 40 видов растений) в кирпичных стенах исторических зданий, построенных в Калифорнии и северной Мексике между 1769 и 1837 гг. Хотя выносливость этих семян и производит большое впечатление, им все же далеко до семян лотоса (Nelumbo nucífera), найденных японскими ботаниками в торфяных отложениях Маньчжурии. Радиоуглеродный анализ показал, что эти семена имеют возраст около 2000 лет, и тем не менее, когда их семенная кожура была разрушена, чтобы облегчить проникновение воды, каждое семя проросло.
В 1967 г. даже этот рекорд был превзойден семенами люпина (Lupinus arcticus) из арктической тундры. Некоторые из этих семян, найденные в замороженной норе лемминга на Юконе вместе с останками животного, возраст которых по крайней мере 10 000 лет, проросли в течение 48 ч. Возраст самих семян, однако, установлен не был.
В последние годы исследователи проявляют повышенный интерес к факторам, позволяющим семенам сохранять жизнеспособность. В хранящихся семенах может наблюдаться постепенная потеря активности ферментов, что в конце концов ведет к полной утрате ими жизнеспособности. Какие условия могут продлить жизнеспособность семян? Подобные вопросы уместны в связи с широким интересом к созданию семенных банков с целью сохранения генетических признаков диких и культивируемых разновидностей сельскохозяйственных растений для использования в будущих программах растениеводства.
Необходимость в таких банках семян возникает в связи с постепенным замещением старых разновидностей новыми и уничтожением замещенных разновидностей главным образом вследствие нарушения их местообитаний. Кроме того, по тем же причинам многие дикорастущие виды подвергаются опасности вымирания, и их семена по возможности должны быть сохранены (см. приложение 2 к гл. 30).
Покой почек
Покой почек имеет существенное значение для выживания травянистых и древесных многолетников умеренных широт, которые подвергаются в зимний период воздействию низких температур. Хотя покоящиеся почки не увеличиваются, т. е. не наблюдается их видимого роста, тем не менее в разные фазы покоя в них обнаружена меристематическая активность.
У многих деревьев почки переходят в состояние покоя в середине лета, задолго до осеннего листопада. Покоящаяся почка представляет собой зачаточный побег, состоящий из апикальной меристемы, узлов и междоузлий (еще нерастянувшихся), рудиментарных листьев или листовых примордиев с почками или примордиями почек в их пазухах. Весь этот зачаточный побег окружен почечными чешуями (рис. 25-19). Почечные чешуи имеют очень важное значение, поскольку препятствуют обезвоживанию, ограничивают поступление кислорода и изолируют почку от потери тепла. Кроме того, известно, что ингибиторы роста накапливаются в почечных чешуях, в осевых частях почек и в листьях внутри почек. Поэтому во многих отношениях роль почечных чешуй сходна с ролью семенной кожуры.
Рис. 25-19. Продольный срез покоящейся пазушной почки клена (Acer). Почка состоит из зачаточного побега, окруженного почечными чешуями
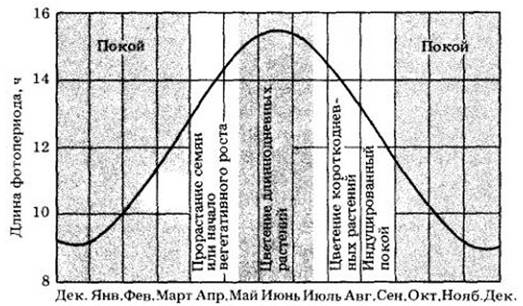
По мере роста и после его прекращения и перехода к покоящемуся состоянию в тканях растений начинают происходить многочисленные физические и физиологические изменения, подготавливающие растение к зиме, т. е. совершается процесс, называемый акклимацией. Уменьшение длины дня в первую очередь способствует переходу почек в состояние покоя (рис. 25-20). Обычно в листьях и почках больше ингибиторов накапливается в условиях короткого дня, чем длинного. Акклимация к холоду приводит к холодостойкости — способности растения выдерживать в зимних условиях сильный холод и засуху.
Рис. 25-20. Связь между длиной дня и циклом развития растений в северной умеренной зоне
Как семенам, так и почкам многих видов растений для прерывания покоя необходим холод. Если ветви деревьев и кустарников осенью срезать и занести в помещение, они не будут цвести; но, если такие ветви выдержать на открытом воздухе до поздней зимы или ранней весны, а затем перенести в теплую комнату, они зацветут. Плодовые деревья с опадающими листьями, такие, как яблоня, каштан и персик, не могут расти в климатической зоне, где не бывает холодных зим. Сходным образом развитие луковиц (например, у тюльпанов, гиацинтов и нарциссов) можно ускорить искусственно, т. е. заставить их цвести в помещении зимой, но только в том случае, если они хранились в холодном месте. Как обсуждалось в гл. 22, такие луковицы фактически представляют собой большие почки, в которых листья видоизменены в мясистые чешуи и накапливают запасные вещества.
Исследования, выполненные в контролируемых условиях, подтверждают предположения, что многим видам растений для того, чтобы выйти из состояния покоя, нужен холод. Например, большинство сортов персика должны находиться в течение 600 — 900 ч при температуре ниже 4°С, прежде чем они смогут реагировать на активирующее влияние более теплых температур и более длинного дня. Некоторые растения реагируют уже на краткое воздействие мороза; если одну почку куста сирени, выращиваемого в теплице, поморозить в течение краткого времени, то вскоре эта единственная почка раскроется. Однако не во всех случаях для прерывания покоя требуется холод. Например, картофелю, «глазки» которого не что иное, как покоящиеся почки, необходимы по крайней мере два месяца сухого хранения; низкая температура не является решающим фактором. У многих растений, особенно деревьев, зимний покой нарушается фотопериодической реакцией, а покоящиеся почки являются при этом рецепторными органами.
Иногда прервать покой могут гиббереллины. Например, обработка гиббереллином почки персика может вызвать ее развитие после того, как почку выдерживали 164 ч при температуре ниже 8°С. Означает ли это, что при нормальных условиях увеличение количества гиббереллинов приводит к прерыванию состояния покоя? Не обязательно. Покой может определяться состоянием баланса между ингибиторами и стимуляторами роста. Добавление какого-либо стимулятора роста (или удаление ингибитора, например, абсцизовой кислоты) может так изменить баланс, что начнется рост.
По-видимому, не существует какого-либо общего механизма, при помощи которого покой индуцируется или прерывается. Этот факт, хотя и в значительной степени усложняет решение проблем, стоящих перед физиологами растений, несомненно, согласуется с нашим представлением об эволюции. Покой стал полезным признаком для растений сравнительно недавно, когда семенные растения начали распространяться по различным экологическим областям. Вероятно, покой возникал независимо у многих групп растений, каждая из которых находила свое собственное решение.
Холод и реакция цветения
Холод может воздействовать и на цветение. Например, если озимая рожь (8еса1е сегеа1е) высевается осенью, она прорастает в течение зимы и зацветает следующим летом, через 7 недель после начала роста. Если рожь высевается весной, она не зацветает в течение 14 недель и остается в вегетативном состоянии большую часть вегетационного периода. В 1915 г. немецкий физиолог растений Густав Гасснер обнаружил, что, регулируя температуру проращиваемых семян, можно воздействовать на цветение озимой ржи и других хлебных злаков. Он заметил, что если семена озимых сортов во время проращивания выдерживать при температурах, близких к замерзанию (1°С), то озимая рожь, даже если она высевается поздней весной, будет цвести этим же летом. Эта процедура, называемая яровизацией, широко применяется в сельском хозяйстве. Даже после яровизации растение должно быть подвергнуто воздействию фотопериодического сигнала; обычно его помещают в условия длинного дня. Яровизированная озимая рожь ведет себя как типичное длиннодневное растение, зацветающее в ответ на длинные летние дни. Аналогичным примером может служить и двулетняя белена (Hyoscyamus niger). Розетка, образованием которой заканчивается первый год вегетации, формирует цветонос только в том случае, если растение подвергнется действию холода. После этого оно становится длиннодневным, с той же фотопериодической реакцией, которая свойственна однолетним растениям.
Как видно из вышеприведенных примеров, обработка холодом некоторых растений влияет на фотопериодическую реакцию. Шпинат можно считать длиннодневным растением, и, как правило, он не цветет, пока длина дня не увеличится до 14 ч. Однако, если семена шпината подвергнуть холодовой обработке, он будет цвести даже при 8-часовой длине дня. Сходным образом холодовая обработка клевера Trifolium subterraneum может полностью устранить зависимость его цветения от длины дня.
У двулетней белены и большинства других двулетних длиннодневных растений, образующих розетки, обработка гиббереллином может заменить действие холода; в этом случае растения быстро растут и затем цветут. Обработка гиббереллином короткодневных растений или длиннодневных, не проходящих через фазу розетки, оказывает незначительный эффект на реакцию цветения (или задерживает ее). Однако, если синтез гиббереллина ингибируется в то время, когда растение подвергается воздействию соответствующего индукционного цикла, оно не зацветет до тех пор, пока гиббереллин не будет восстановлен.
Настические движения
Настическими называют происходящие в ответ на какое- либо воздействие движения растений, направления которых не зависят от направления воздействия. По-видимому, наиболее широко встречающимися настическими движениями можно считать складывание листьев, обсуждавшееся ранее в разделе «Циркадианные ритмы». Известные под специальным названием никтинастических движений (от греческих слов, означающих «ночное смыкание»), они представляют собой движение листьев вверх и вниз в ответ на суточные изменения освещенности; при этом листья ориентируются вертикально в темноте и горизонтально на свету. Такие движения особенно характерны для бобовых растений.
Чаще всего никтинастические движения листьев происходят в результате изменений размеров клеток основной паренхимы в листовых подушечках, расположенных у основания каждого листа (или, если лист сложный, у основания каждого листочка). Листовая подушечка представляет собой гибкий цилиндр, в центре которого находится проводящая система. Большую часть листовой подушечки занимают тонкостенные клетки паренхимы, окружающие проводящую ткань. Движения связаны с изменениями тургора и сопутствующими сокращениями-растяжениями клеток основной паренхимы на противоположных сторонах листовой подушечки. Тургорные изменения при сокращении и растяжении клеток происходят за счет челночного передвижения ионов калия между двумя сторонами подушечки — «местоположения» биологических часов и фоторецепторного фитохрома.
Настические движения, которые вызываются прикосновением, называют тигмонастическими. Такие движения характерны для хорошо известного чувствительного растения мимоза стыдливая (Mimosa púdica), чьи листочки, а иногда и листья целиком внезапно опускаются в ответ на прикосновение (рис. 25-21). Как и в случае никтинастического движения, эта реакция является результатом внезапного изменения тургорного давления в некоторых клетках листовой подушечки у основания листочков или листьев. Потеря этими клетками воды следует за выходом из них ионов калия. Достаточно стимуляции лишь одного листочка, чтобы стимул распространился на другие части листа, а далее и по всему растению.
Рис. 25-21. Чувствительное растение Mimosa púdica. А. Нормальное положение листьев и листочков. Реакция на прикосновение иглой показана на рис. Б и В. Она происходит за счет изменения тургорного давления в листовых подушечках, расположенных у основания листочков, причем и в том случае, если стимулируется только один листочек (В)

По-видимому, два отдельных механизма — электрический и химический — включаются при распространении стимула в восприимчивом растении. Имеются разные мнения по поводу значения тигмонастических движений для выживания растения. Mimosa púdica часто растет в засушливых областях, где может подвергаться действию суховеев; сильные ветры, раскачивая листья, могут заставить их сложиться, что способствует сохранению воды. Другое предположение заключается в том, что «увядшее» растение непривлекательно для многочисленных фитофагов. Наконец, вполне допустимо, что эффект складывания листьев пугает растительноядных насекомых; имеются указания, что другие «нечувствительные» виды Mimosa, растущие вблизи Mimosa púdica, больше подвергаются нападению насекомых.
До недавнего времени предполагалось, что в реакции на прикосновение у насекомоядной венериной мухоловки (Dionaea muscipula), которая дает возможность растению захватить свою жертву, участвуют изменения тургорного давления. Однако теперь представляется, что быстрое закрывание листьев венериной мухоловки связано с необратимым растяжением клеток, которое может начинаться с подкисления клеточных стенок до pH 4,50 и ниже. По мере закрывания листьев клетки нижней эпидермы центральной части каждого листа частично растягиваются, в то время как размеры клеток верхней эпидермы не подвергаются значительному изменению. При открывании листьев растягиваются клетки верхней эпидермы центральной части листа, в то время как клетки нижней эпидермы остаются неизменными. Эти изменения могут быть предотвращены нейтральными буферами, которые препятствуют подкислению клеточных стенок до pH 4,5 — 4,74. Изменения в уровне АТР, измеряемом по мере закрывания листа, показывают, что около трети клеточного АТР исчезает за время, требующееся для захлопывания ловушки (1 — 3 с). Возможно, что АТР используется для очень быстрого транспорта ионов водорода из клеток.
Каждая половина листа венериной мухоловки снабжена тремя чувствительными волосками. Если насекомое попадает на один из листьев, привлеченное нектаром, выделяемым на его поверхность, оно задевает волоски и запускает таким образом механизм закрывания листа, действующий подобно ловушке. Зубчатые края смыкаются, половинки листа постепенно сжимаются, и насекомое оказывается прижатым к переваривающим железкам, расположенным на поверхности ловушки (рис. 25-22).
Рис. 25-22. Реакция на прикосновение у венериной мухоловки (Dionaea muscipula). Здесь можно видеть неосторожную муху, привлеченную нектаром, выделенным на поверхность листа, до и после его закрывания. Каждая половинка листа имеет три чувствительных волоска, которые контролируют «ловушку». Когда муха касается одного чувствительного волоска дважды или последовательно двух волосков, она тем самым вызывает смыкание половинок листа
Ловчий механизм настолько специализирован, что может отличать живую добычу от неживых объектов, например, песчинок и мелких частиц сухих растений, случайно попадающих на лист; лист не сомкнется до тех пор, пока не будут затронуты подряд два его волоска или один из волосков не будет затронут дважды.
Гелиотропизм
Листья и цветки многих растений в течение суток могут поворачиваться, ориентируясь перпендикулярно или параллельно солнечным лучам. Это явление имеет специальное название гелиотропизм (от греч. helios — солнце). В отличие от фототропизма стебля движение листа гелиотропного растения не является результатом асимметричного роста. В большинстве случаев в движении участвуют подушечки у основания листьев и/или листочков. Некоторые черешки, по-видимому, имеют свойства подушечек вдоль всей длины или большей ее части. Среди обычных растений, у которых наблюдается гелиотропизм листьев, можно назвать хлопчатник, сою, люпин и подсолнечник (рис. 25-23).
Рис. 25-23. А. Листья люпина (Lupinus arizonicus), поворачивающиеся вслед за солнцем. Это явление называют гелиотропизмом. Б. Гелиотропизм у подсолнечника

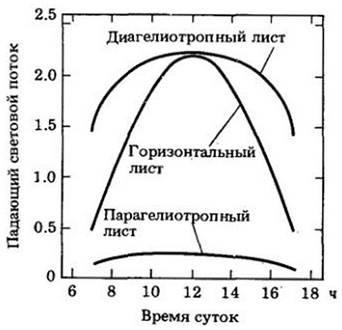
Различают два типа гелиотропизма. При одном (диагелиотропизм) листовые пластинки поворачиваются таким образом, что в течение всего дня остаются перпендикулярными к прямым солнечным лучам. Такие листья получают больше квантов (рис. 25-4), вовлекаемых в фотосинтез и, по- видимому, имеют большую скорость фотосинтеза в течение всего дня, чем «не следящие» за солнцем или парагелиотропные листья (см. ниже).
В засушливые периоды некоторые гелиотропные растения активно уклоняются от прямого солнечного света, ориентируя свои листовые пластинки параллельно солнечным лучам (парагелиотропизм). Кроме того, что такая ориентация скорее уменьшает поглощение солнечного света, чем увеличивает его (рис. 25-24), она снижает и температуру листа, и транспирационную потерю воды, способствуя выживанию в засушливые периоды.
Рис. 25-24. Сравнение фотосинтетически активной солнечной радиации (400 — 700 нм), падающей на диагелиотропный, негелиотропный (горизонтальный) и парагелиотропный листья в течение суток
Выводы
Каждый этап в жизни растения находится под контролем не одного какого-либо фактора, а комплекса различных факторов. Внутренние и внешние факторы взаимодействуют между собой. Они могут усиливать, изменять или нейтрализовать друг друга. Как сказал У. Хиллмен из Брукхейвенской национальной лаборатории (США): «В конце концов, если бы растения были такими простыми, какими хотели бы их видеть физиологи, тогда последним нечего было бы делать».
Заключение
Растения обладают многими приспособлениями, которые позволяют им обнаруживать изменения в окружающей среде и реагировать на них. Примером такой адаптации может служить фототропизм, или изгибание растущего побега по направлению к свету. Неравномерный рост проростка вызывается латеральным перемещением ростового гормона ауксина под влиянием света. Фоторецептором для такой реакции служит пигмент, поглощающий синий свет. Гравитропизм, или геотропизм — это реакция побега или корня на земное притяжение. Перемещение ауксина к нижней поверхности горизонтально ориентированного побега может способствовать изгибанию побега вверх. Изгибание вниз горизонтально ориентированною корня частично происходит в результате асимметричного распределения тормозящих рост гормональных веществ, образующихся в корневом чехлике и поступающих в зону роста корня. Тигмотропизм представляет собой реакцию на контакт с твердым предметом.
Циркадианные ритмы — это циклы физиологической активности организма, повторяющиеся с интервалом около 24 ч при постоянных условиях среды. Они, вероятно, являются эндогенными и вызываются не внешними факторами, такими, как чередование света и темноты или вращение Земли, а контролируются каким-то внутренним регуляторным механизмом. Такой регуляторный механизм, химическая и физическая природа которого нам неизвестна, называется биологическими часами. Биологические часы позволяют организму воспринимать изменения во внешних суточных циклах, таких, как изменение длины дня. В результате такие явления, как покой, опадение листьев, цветение, могут осуществляться синхронно с внешними условиями.
Реакция организмов на изменения 24-часовых циклов света и темноты известна как фотопериодизм, контролирующий у многих растений начало цветения. Некоторые растения цветут только при длине дня, превышающей критическую. Такие растения называют длиннодневными. Другие растения — короткодневные — цветут только при длине дня меньше критической. Нейтральные к длине дня растения цветут независимо от фотопериодов. На фотопериодическую реакцию могут оказывать влияние такие факторы, как температура и возраст растения. Прерывание темновой фазы фотопериода даже короткой вспышкой света может менять направление фотопериодического воздействия.
Фитохром — пигмент, присутствующий обычно в малых количествах в тканях высших растений, — чувствителен к чередованию света и темноты. Пигмент может существовать в двух формах: Рк и Рдк; Рк поглощает красный свет с длиной волны 660 нм и таким образом превращается в Рдк; Рдк поглощает дальний красный свет (730 нм) и превращается в Рк. Рдк в темноте превращается в Рк или разрушается. Рдк — это активная форма пигмента: она ускоряет цветение длиннодневных растений и задерживает цветение короткодневных. Рдкответственна и за изменения, происходящие в проростках, когда они выходят из почвы на свет, за прорастание семян и образование антоцианов.
Как у длиннодневных, так и у короткодневных растений фотопериод воспринимается листьями, а ответная реакция возникает в почке. Фитохром — это фоторецептор, воспринимающий стимул из окружающей среды и, по-видимому, каким-то образом взаимодействующий с эндогенным циркадианным ритмом, вызывая образование стимула цветения. Хотя до настоящего времени стимул цветения не выделен и не идентифицирован, это химическое вещество, называемое флоригеном, перемещается из листьев к почке, где вызывает цветение. Эксперименты показали, что перемещение происходит по флоэме и что структура и функция флоригена сходны у длиннодневных, короткодневных и нейтральных к длине дня растений. Есть серьезные основания предполагать, что по крайней мере у некоторых растений в процессе цветения участвуют как вещества, индуцирующие цветение, так и задерживающие его.
Чередование периодов роста и покоя позволяет растению переносить недостаток воды, а также сильную жару и холод. Покой — это особое состояние остановки роста, при котором все растение или такие структуры, как семена или почки, не возобновляют рост без специального воздействия со стороны окружающей среды. Потребность в таком воздействии, скажем холода, высушивания и соответствующего фотопериода, предохраняет ткани от нарушения покоя во время некоторых благоприятных условий, например, существующих внутри сочного плода родительского растения или появляющихся иногда осенью, во время бабьего лета. По- видимому, нет единого механизма, общего для всех групп растений, который бы вызывал покой или нарушал его. Уменьшение длины светового дня является первым фактором, который включается в индукцию покоя в почках. Акклимация к холоду приводит к холодостойкости, способности растения переносить Сильные зимние холода. Яровизация способствует ускорению цветения озимых сортов растений путем хранения семян при низких температурах.
Движения растений, которые происходят в ответ на раздражение, но направление которых не зависит от направления стимула, называют настическими. Среди них различают широко встречающиеся движения «засыпания» — поднимание и опускание листьев в ответ на суточный ритм света и темноты. Настические движения, возникающие как реакция на прикосновение, включают и действие ловчего механизма венериной мухоловки.
Листья и цветки некоторых растений «следят» за солнцем в течение дня, увеличивая или уменьшая поглощение солнечной энергии.