СОВРЕМЕННАЯ БОТАНИКА - П. РЕЙВН - 1990
РАЗДЕЛ VII. ПОГЛОЩЕНИЕ И ТРАНСПОРТ ВЕЩЕСТВ У РАСТЕНИЙ
ГЛАВА 26. ПИТАНИЕ РАСТЕНИЙ И ПОЧВА
Общие пищевые потребности
Растения должны получать из окружающей среды определенные вещества, вовлекаемые в сложные биохимические реакции, в результате которых поддерживаются структура и рост клеток. Помимо света высшим растениям для метаболизма и роста нужны вода и некоторые химические элементы. Многие эволюционные приобретения растений связаны со структурными и функциональными специализированными приспособлениями для эффективного поглощения этих веществ и для распределения их по живым клеткам своего тела.
Подобно растениям, животные нуждаются в воде и определенных химических элементах, однако они должны получать извне также сахара или другие компоненты, которые могут служить источником энергии, а кроме того, еще некоторые аминокислоты и витамины. По сравнению с животными пищевые потребности растений относительно просты. При благоприятных внешних условиях большинство зеленых растений может использовать энергию света для превращения СО2 и Н2О в органические соединения, служащие источником энергии. Растения могут синтезировать и все необходимые аминокислоты, и витамины, используя неорганические питательные вещества, поступающие из окружающей среды.
Особенно поразительна способность к самообеспечению азотфиксирующих цианобактерий. В процессе фиксации азота они превращают атмосферный азот в такие его формы, которые могут быть использованы для синтеза аминокислот и белков. Помимо этого, цианобактерии обладают еще и фотосинтетической активностью.
Питание растений включает поглощение из среды всех исходных веществ, необходимых для биохимических реакций, распределение этих веществ по растению и использование их в процессах метаболизма и роста.
Необходимые неорганические питательные вещества
Уже к 1800 г. химики и биологи установили, что некоторые химические элементы поглощаются растениями из среды. Однако мнения расходились — являются ли поглощенные элементы загрязнением или компонентами, необходимыми для нормального функционирования. К середине 1800-х гг. было выяснено, что по крайней мере 10 химических элементов, присутствующих в растениях, необходимы для нормального роста. В отсутствие любого из них у растений наблюдаются характерные нарушения роста или симптомы повреждения. Часто такие растения не могут нормально размножаться. Эти десять элементов — углерод, водород, кислород, калий, кальций, магний, азот, фосфор, сера и железо — были определены как химические элементы, необходимые для роста растений. Они, таким образом, относятся к необходимым минеральным, или необходимым неорганическим питательным веществам.
В начале нашего века было установлено, что и марганец относится к необходимым элементам. В течение последующих 50 лет благодаря улучшению технологии очистки питательных растворов от примесей к числу необходимых было отнесено еще пять элементов: цинк, медь, хлор, бор и молибден, причем хлор был помещен в эту группу только в 1954 г. В настоящее время данные 16 элементов считаются необходимыми для нормального развития большинства растений (рис. 26-2).
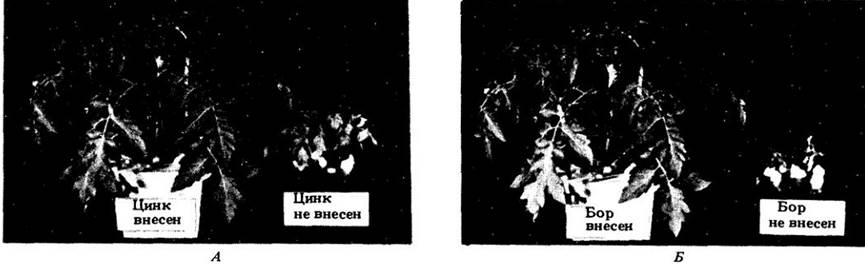
Рис. 26-2. Недостаток одного или нескольких необходимых неорганических питательных элементов приводит к нарушениям роста и развития растения. Показано влияние дефицита цинка и бора на томаты (А и Б соответственно). На каждой фотографии слева помещено контрольное растение
Концентрация питательных веществ в растениях
Обычным приемом определения относительного количества различных элементов, необходимых для нормального роста разных видов растений, является химический анализ содержания неорганических веществ. Типичные результаты подобных исследований представлены в табл. 26-1. Они особенно важны для сельского хозяйства, показывая обеспеченность растений питательными веществами и возможную необходимость внесения удобрений. Анализ неорганических веществ может предсказать и потенциальные нарушения питания скота, потребляющего определенные растения.
Таблица 26-1. Примеры анализа содержания неорганических элементов в растениях (в пересчете на сухую массу)
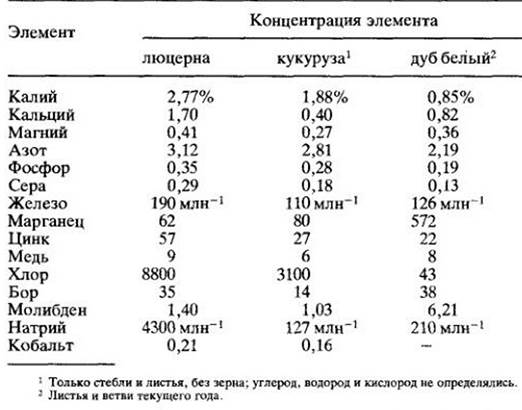
Известно, что отдельные элементы содержатся в растениях в различных концентрациях. Исходя из этого основные неорганические вещества делятся на две большие группы — макро- и микроэлементы. Макроэлементы требуются растениям в больших количествах, микроэлементы (следовые элементы) — в очень малых, следовых количествах (табл. 26-2).
После углерода и кислорода наибольших концентраций в растениях достигает калий. Когда велико его содержание в почве, в растениях он может составить до 10% сухой массы.
Таблица 26-2. Функции неорганических питательных элементов в растениях
Элемент |
Основная форма, в которой поглощается элемент |
Обычная концентрация в здоровом растении (в пересчете на сухую массу) |
Главные функции |
Макроэлементы |
|||
Углерод |
СО2 |
-44% |
Компонент органических соединений |
Кислород |
Н2O или О2 |
-44% |
Компонент органических соединений |
Водород |
H2O |
-6% |
Компонент органических соединений |
Азот |
NO-3 или NH+4 |
1-4% |
Компонент аминокислот, белков, нуклеотидов, нуклеиновых кислот, хлорофилла и коферментов |
Калий |
К+ |
0,5 — 6% |
Структура ферментов, аминокислот. Синтез белка. Активатор многих ферментов. Открывание и закрывание устьиц |
Кальций |
Са2+ |
0,2 — 3,5% |
Компонент клеточных оболочек. Кофактор ферментов. Клеточная проницаемость. Компонент кальмодулина — регулятора работы мембран и ферментов |
Фосфор |
Н2РО-4 или НРО2-4 |
0,1 — 0,8% |
Образование «высокоэнергетических» фосфатных соединений (АТР в A DP), фосфорилирование сахаров. Компонент нуклеиновых кислот, нескольких важных коферментов, фосфолипидов |
Магний |
Mg2+ |
0,1 — 0,8% |
Компонент молекулы хлорофилла. Активатор многих ферментов |
Сера |
SO2- |
0,5 — 1% |
Компонент некоторых аминокислот, белков, кофермента А |
Микроэлементы |
|||
Железо |
Fe2+ или Fe3+ |
25 — 300 млн-1 |
Синтез хлорофилла. Компонент цитохромов, нитрогеназы |
Хлор |
СI |
100-1000 млн1 |
Осмос и ионный баланс; по-видимому, необходим для фотосинтетических реакций образования кислорода |
Медь |
Cu2+ |
4 — 30 млн-1 |
Активатор некоторых ферментов |
Марганец |
Mn2+ |
15 — 800 млн-1 |
Активатор некоторых ферментов |
Цинк |
Zn2+ |
15 — 100 мл н-1 |
Активатор многих ферментов |
Молибден |
MoO2-4 |
0.1 — 5,0 млн-1 |
Фиксация азота, восстановление нитратов |
Бор |
ВО-3 или B4O2-7 |
5 — 75 млн-1 |
Влияет на использование Са2+. Неизвестные функции |
Элементы, необходимые для некоторых растений или других организмов |
|||
Кобальт |
Co2+ |
Следы |
Требуется азотфиксирующим микроорганизмам |
Натрий |
Na+ |
Следы |
Осмос и ионный баланс, но лишь у некоторых растений. Требуется некоторым видам в пустынях и на солончаках, возможно всем растениям с С4-типом фотосинтеза |
Некоторые виды или таксономические группы характеризуются необычно высоким или низким содержанием отдельных элементов (рис. 26-3). Даже произрастая на одном и том же питательном растворе, растения могут резко различаться по концентрации минеральных веществ. Сравните результаты анализов у кукурузы (однодольное) и люцерны (двудольное), приведенные в табл. 26-1. Двудольным обычно требуется значительно больше кальция и бора, чем однодольным.
Исследования показали, что некоторые элементы необходимы только определенным растениям или растениям, произрастающим в специфических условиях. Люцерна (Medicado sativa) и другие бобовые хорошо отзываются на добавку кобальта. Однако кобальт требуется нс самой люцерне, а симбиотическим азотфиксирующим бактериям, растущим на ее корнях. Как следует из табл. 26-1, в отдельных растениях содержится довольно много натрия. Давно было установлено, что натрий может частично удовлетворить потребность в калии у некоторых видов. Относительно недавно удалось показать, что натрий необходим еще ряду растений. Например, он требуется галофитам (растениям, произрастающим на засоленных почвах). Недавно выяснилось, что в отсутствие никеля растения сои замедляют рост и на их листьях появляются некротические пятна, где в токсических концентрациях накапливается мочевина Добавление никеля в питательную среду предотвращает появление этих симптомов, что указывает на необходимость данного элемента для сои.
В табл. 26-1 включены сведения о двух элементах — натрии и кобальте, которые не считаются необходимыми для всех сельскохозяйственных растений. Однако эти элементы нужны травоядным животным, питающимся данными растениями. Например, если концентрация кобальта в кормовых растениях ниже 0,1 млн -1, то у животных появляются симптомы его недостаточности.
Функции неорганических питательных веществ в растении
Неорганические вещества необходимы для роста и метаболизма растений по разным причинам. В табл. 26-2 перечисляются элементы, требующиеся большинству растений; формы, в которых они, как правило, поглощаются из среды; их обычные концентрации и некоторые из функций.
Специфические и неспецифические функции
Неорганические ионы влияют на осмотическое давление (гл. 4) и таким образом помогают регулировать водный баланс. Поскольку некоторые из ионов в этой роли взаимозаменяемы, то потребность растений в них можно считать неспецифической. С другой стороны, неорганический компонент может функционировать как часть незаменимой биологической молекулы. В этом случае потребность в нем высокоспецифична. Например, специфическую функцию выполняет магний, входящий в состав молекулы хлорофилла (см. рис. 7-8). Некоторые неорганические вещества входят в состав клеточных мембран, другие — контролируют их проницаемость. Ряд элементов — это обязательные компоненты ферментных систем, катализирующих биологические реакции в клетке. Другие формируют ту особую ионную среду, в которой могут протекать биологические реакции.
Вследствие того, что неорганические питательные элементы необходимы для удовлетворения основных потребностей организма и вовлечены в фундаментальные процессы, их недостаток вызывает разнообразные структурные и функциональные изменения растений.
Катализаторы
Очень важна роль неорганических питательных элементов как катализаторов некоторых ферментативных реакций в растительной клетке. В одних случаях они представлены необходимой составной частью («простетической группой») ферментов. В других — выступают как активаторы или регуляторы ферментов. Считается, например, что калий, влияющий на активность 50 или 60 ферментов, регулирует конформацию некоторых из них. В результате изменения конфигурации молекулы фермента се реакционный центр становится более или менее доступным для связывания субстрата (см. рис. 3-18).
Перенос электронов
Многие из биохимических процессов, в том числе фотосинтез и дыхание, представляют собой систему окислительновосстановительных реакций. В этих реакциях электроны передаются молекулам, функционирующим в качестве акцепторов. К переносчикам электронов относятся цитохромы, в состав которых входит железо (см. рис. 6-9).
Структурные компоненты
Некоторые минеральные элементы входят в состав различных клеточных компонентов, в том числе физических (см. рис. 26-3, Б) и химических структур, участвующих в метаболизме. Кальций соединяется с пектиновой кислотой в срединной пластинке клеточной оболочки. Фосфор встроен в «остов» спиралей ДНК и РНК, а также входит в состав фосфолипидов клеточных мембран. Азот— обязательный компонент аминокислот, хлорофилла и нуклеотидов. Сера присутствует в нескольких аминокислотах, являясь, таким образом, важным структурным элементом многих белков.
Рис. 26-3. А. Растения семейства крестоцветных, такие, как сурепка (Barbarea vulgaris), используют серу для синтеза горничных масел, которые придают им характерный горький вкус. Б. Хвощи (Equisetum) включают в клеточные оболочки кремний, что делает их несъедобными для большинства травоядных, но зато пригодными для чистки горшков и кастрюль, что практиковалось в колониальной Северной Америке

Осмос
Поступление воды в растительную клетку и из нее, как обсуждалось в гл. 4, в значительной степени зависит от концентрации растворенных веществ в клетке и в окружающей среде. Поглощение ионов растительными клетками может сопровождаться поэтому поступлением в них воды. Возникающее тургорное давление, направленное на клеточную оболочку, приводит к растяжению и, следовательно, росту незрелых клеток и поддержанию тургора зрелых (см. гл. 4). Это пример превращения одной формы энергии в другую, осуществляемого живой системой: химическая энергия (АТР), затраченная на активное поглощение ионов растительными клетками, переходит в физическую энергию движения воды.
Влияние на проницаемость клеток
Кальций оказывает непосредственное влияние на физические свойства клеточных мембран. Его недостаток приводит к тому, что мембраны теряют свою целостность и растворенные вещества начинают выходить из клеток.
Почва
Почва служит растениям основной питательной средой. Она постоянно обеспечивает им не только физическую опору, но и снабжает их необходимыми минеральными веществами и водой, а также создает подходящую газовую среду для корневых систем. Знания о происхождении разных почв, их химических и физических свойствах и способности удовлетворить потребности растения крайне важны для планирования режима питания посевов.
Выветривание земной коры
Все неорганические питательные элементы, используемые растениями, происходят либо из атмосферы, либо из продуктов выветривания горных пород. Земная кора построена примерно из 92 природных элементов, которые часто находятся в форме минералов. Минералами называются неорганические природные соединения, состоящие обычно из двух или более элементов в определенных массовых соотношениях. Примерами минералов могут служить кварц (SiO2) и кальцит (СаСО3).
Горные породы обычно состоят из нескольких различных минералов и по своему происхождению и строению делятся на три основные группы. Магматические породы, такие, как гранит, произошли непосредственно из расплавленного материала и, как правило, сформировались еще в те времена, когда Земля остывала и затвердевала. В результате выветривания магматические породы разрушаются и распадаются на растворимые и нерастворимые составные части. Переносимые водой, ветром или ледниками, они образуют — обычно в воде — осадки, которые со временем уплотняются и отвердевают, превращаясь в осадочные породы, такие, как глинистый сланец, песчаник или известняк. Несмотря на то что осадочные породы составляют только около 5% земной коры, они имеют очень большое значение, поскольку широко распространены вблизи (или на) поверхности Земли. В условиях экстремально высокой температуры и огромного давления в глубинах Земли осадочные и магматические породы превращаются в породы третьего типа — метаморфические. Так, из песчаника образуется кварцит, из глинистого сланца — сланец кристаллический, из известняка — мрамор.
В результате выветривания, приводящего к физическому разрушению и химическому распаду минералов и горных пород, образуются неорганические вещества, из которых формируется почва. Выветривание включает замерзание и оттаивание, нагрев и растрескивание пород. Вода и ветер часто переносят их обломки на большие расстояния, перетирая их при этом и дробя на более мелкие частицы. Вода вымывает из них растворимые вещества. Из воды, углекислого газа и некоторых примесей, содержащихся в воздухе, таких, как двуокись серы или окись азота, образуются разбавленные кислоты, способствующие растворению тех веществ, которые плохо растворяются в чистой воде. Почва образуется либо в месте выветривания, либо там, куда сила тяжести, ветер, вода или ледник перенесут остатки материнской породы. Почва содержит и органические вещества. Если позволяют свет и температура, на выветривающихся породах и минералах поселяются бактерии, грибы, водоросли, лишайники, моховидные и мелкие сосудистые растения. Растущие корни еще больше разрушают горные породы, а разлагающиеся остатки растений и животных способствуют накоплению органических веществ. Затем появляются более крупные растения, укрепляющие почву своими корневыми системами, и возникает новое сообщество (рис. 26-4).
Рис. 26-4. Мочковатые корни злаков укрепляют почву в прерии

На вертикальном почвенном разрезе (рис. 26-5) можно увидеть различия в цвете, количестве живого и мертвого органического материала, пористости, структуре и степени выветривания. В результате довольно отчетливо выявляются отдельные слои, которые почвоведы называют горизонтами. Выделяют не менее трех горизонтов.
Рис. 26-5. Три основных горизонта, или почвенных слоя, выделяемых в типичной почве

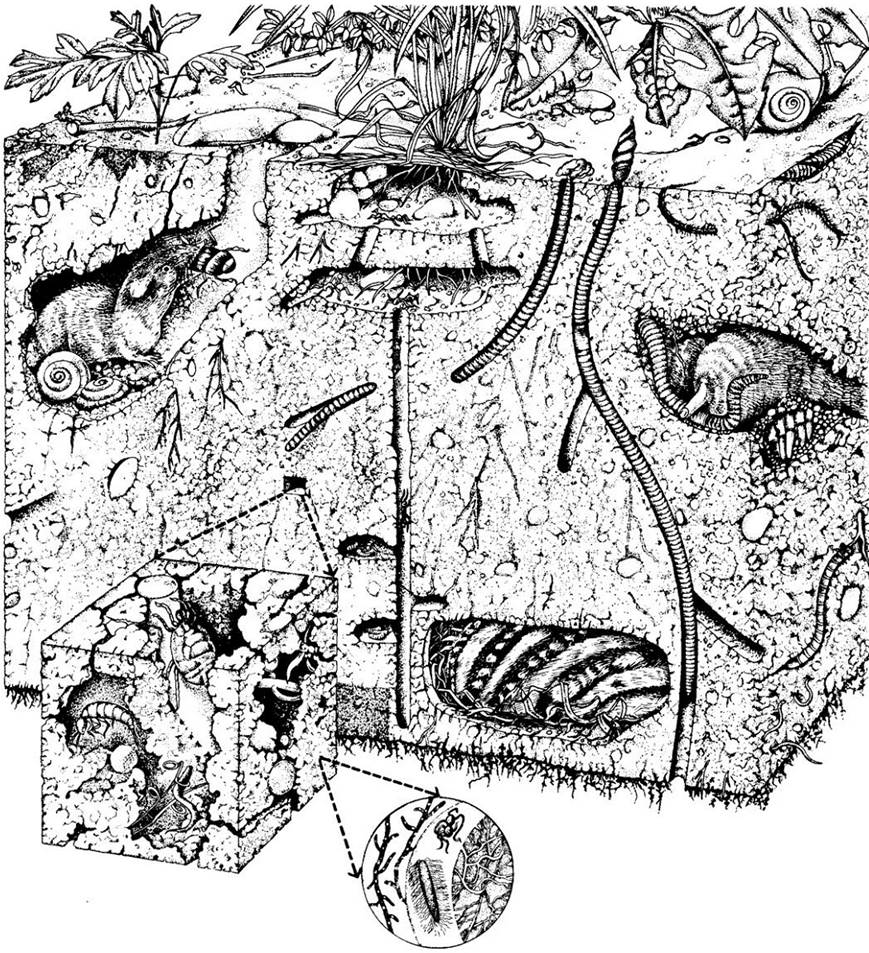
Горизонт А — это верхний слой с наибольшей физической, химической и биологической активностью. В нем находится большая часть органического материала почвы как живого, так и мертвого, в том числе много опавших и гниющих листьев и других частей растений, большое количество насекомых и других мелких членистоногих, земляных червей, простейших, нематод и других организмов-деструкторов (рис. 26-6).
Рис. 26-6. В почве наряду с растениями обитает множество живых организмов — от микробов до мелких млекопитающих, таких, как кроты, землеройки и бурундуки. Масса роющих существ, среди которых особое место занимают муравьи и дождевые черви, аэрирует почву и повышает ее способность впитывать воду. Дождевые черви, которых Аристотель назвал «кишками земли», улучшают почву, пропуская ее через свой пищеварительный тракт. Эта улучшенная почва выбрасывается на поверхность. За один год дождевые черви совместными усилиями могут переработать до 500 т почвы на гектар. Эта почва очень плодородна и содержит по сравнению с окружающей в 5 раз больше азота, в 7 раз — фосфора, в 11 раз — калия, в 3 раза — магния и в 2 раза — кальция. Бактерии и грибы — это основные деструкторы органического вещества в почве
Горизонт В — это слой накопления. В нем задерживаются окислы железа, частицы глинозема и некоторое количество органики, которые попадают сюда из горизонта А с водой, просачивающейся вглубь. Горизонт В содержит значительно меньше органических веществ и менее выветрен по сравнению с вышележащим горизонтом.
Горизонт С состоит из обломков материнской породы и минералов, из которых образуется собственно почва верхних горизонтов.
Состав почвы
Почва состоит из твердого материала и порового пространства (пространства между почвенными частицами). В зависимости от условий увлажнения поровое пространство заполняется воздухом и водой в разных соотношениях. Значительная часть почвенной воды представлена пленкой на поверхности почвенных частиц.
Фрагменты пород и минералов в почве варьируют по размерам от песчинок, видимых невооруженным глазом, до глинистых частиц, которые нельзя рассмотреть под микроскопом при малом увеличении. Вот одна из классификаций почвенных частиц (называемых также фракциями) в соответствии с их размерами:
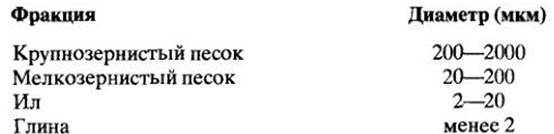
Почвы содержат смесь частиц разного размера и делятся на классы механического состава в зависимости от количественного соотношения этих частиц. Например, почвы, которые содержат 35% и более глины и 45% и более песка, называются глинисто-песчаными; содержащие 40% и более глины и 40% и более ила — иловато-глинистыми. Суглинистые почвы содержат песок, ил и глину в соотношениях, которые делают эти почвы идеальными для земледелия.
Твердый материал разных почв состоит из неорганических и органических компонентов в различных соотношениях. Органические компоненты содержат остатки организмов на разных стадиях разложения, а также много разнообразных живых растений и животных. В живую фазу почвы могут быть включены такие крупные образования, как корни деревьев, однако преобладают в ней грибы, бактерии и другие микроорганизмы.
Kaтионный обмен
Неорганические питательные элементы, поглощаемые корнями растений, существуют в почвенном растворе в виде ионов. Большинство металлов образует положительно заряженные ионы — катионы, такие, как Са2+, К+, и Nа+. Глинистые частицы играют роль резервуара катионов для растения. В разных точках их кристаллической решетки имеется избыток отрицательного заряда, где катионы связываются и таким образом удерживаются, несмотря на вымывающее действие воды, просачивающейся сквозь почву.
Катионы, связанные с глинистыми частицами, в процессе катионного обмена могут замещаться другими и переходить в почвенный раствор, становясь доступными для растений. Это одна из причин того, что глинистые частицы являются необходимым компонентом плодородной почвы.
Основные отрицательно заряженные ионы (анионы), содержащиеся в почве, — это NO-3, SO2-4, HCO-3, и ОН-. Анионы вымываются из почвы быстрее, чем катионы, поскольку они не прикрепляются к глинистым частицам. Исключение представляет собой фосфат, который устойчив к вымыванию из-за того, что образует нерастворимые осадки и избирательно поглощается или удерживается соединениями, содержащими железо, алюминий и кальций.
Доступность неорганических питательных элементов для растений определяется также кислотностью или щелочностью почвы. Диапазон pH различных почв достаточно широк, а многие растения переносят лишь небольшие его изменения. В щелочных почвах некоторые катионы образуют нерастворимые соединения, и такие элементы, как железо, магний, медь и цинк, становятся недоступными для растений.
Почва и вода
Около 50% общего объема почвы занимает поровое пространство, заполненное воздухом и водой в разных соотношениях, зависящих от условий увлажнения. Если вода заполняет поровое пространство не более чем наполовину, корни растений снабжаются кислородом в достаточном количестве.
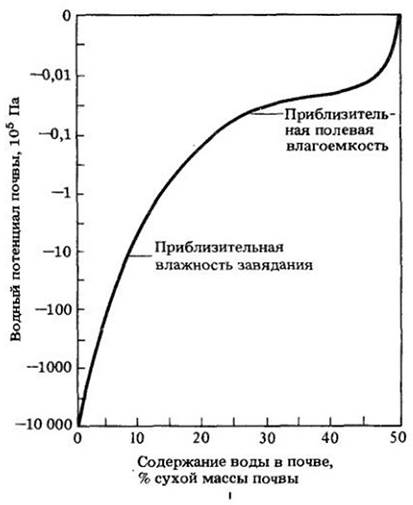
После сильного дождя или полива почва сохраняет определенное количество воды и остается влажной даже после ухода из нее под влиянием силы тяжести слабо связанной воды. Если фрагменты, составляющие почву, велики, то велики и поры, и все пространство между фрагментами. Вода в этом случае быстро проходит сквозь почву, и се остается сравнительно мало в горизонтах А и В. Из глинистых почв благодаря мелким порам под действием силы тяжести уходит меньшее количество воды. Таким образом, в глинистых почвах может удерживаться в 3 — 6 раз больше воды, чем в песчаных того же объема. Процент воды, удерживаемый почвой, несмотря на действие силы тяжести, называется полевой влагоемкостью.
Если растению предоставить ограниченный объем почвы и не поливать его, то оно в конце концов не сможет извлекать воду из почвы, поникнет и завянет. В случае сильного увядания растение не сможет вернуться к жизни, даже если его поместить во влажную камеру. Количество воды (в процентах), которое еще остается в почве, когда у растения наступает необратимое увядание, называется влажностью устойчивого завядания для данной почвы.
На рис. 26-7 показано соотношение между содержанием воды и потенциалом, удерживающим воду в супесчаной почве. Силы, сохраняющие воду в почве, можно выразить в тех же единицах (в данном случае через величину водного потенциала), что и силы, определяющие поглощение воды клетками и тканями (см. гл. 4). Водный потенциал почвы постепенно уменьшается по мере уменьшения влажности ниже уровня полевой влагоемкости. Когда содержание воды снижается до влажности устойчивого завядания (около — 15 бар), происходит резкое уменьшение водного потенциала почвы.
Рис. 26-7. Соотношение между водным потенциалом и содержанием воды в супесчаной почве
Круговорот питательных веществ
Органическое вещество почвы образуется главным образом из опавших листьев и других частей растений, а также из разлагающихся тел животных. Органические остатки перемешаны с неорганическими частицами почвы, и эта смесь населена поразительным числом мелких организмов, которые проводят всю жизнь или часть жизни под поверхностью почвы. В одной чайной ложке почвы может находиться 5 млрд, бактерий, 20 млн. мелких нитчатых грибов и 1 млн. протистов. Почвенные животные и микроорганизмы (см. рис. 26-6) разлагают органическое вещество, высвобождая его неорганические компоненты, которые затем снова утилизируются растениями. Таким образом, несмотря на то что питательные вещества вымываются из почвы, уносятся в водотоки и выпадают в осадки на дне океана, они опять попадают в почву. Макро- и микроэлементы совершают постоянный круговорот через растительные и животные организмы, возвращаются в почву и снова поглощаются растениями. Каждый элемент имеет свой особый цикл, включающий различные организмы и разные ферментные системы. Конечный результат, однако, один и тот же — значительное количество элемента постоянно возвращается в почву и становится доступным для растений.
Азот и круговорот азота
Азот почвы происходит из атмосферы. Хотя атмосфера на 78% состоит из азота, большинство организмов не может использовать его для построения белков и других органических веществ. В отличие от углерода и кислорода азот химически инертен. Высокоспециализированной способностью превращать азот атмосферы в форму, которую могут усваивать живые клетки, обладают лишь некоторые бактерии. Этот уникальный процесс, называемый азотфиксацией, будет рассмотрен лишь в основных чертах.
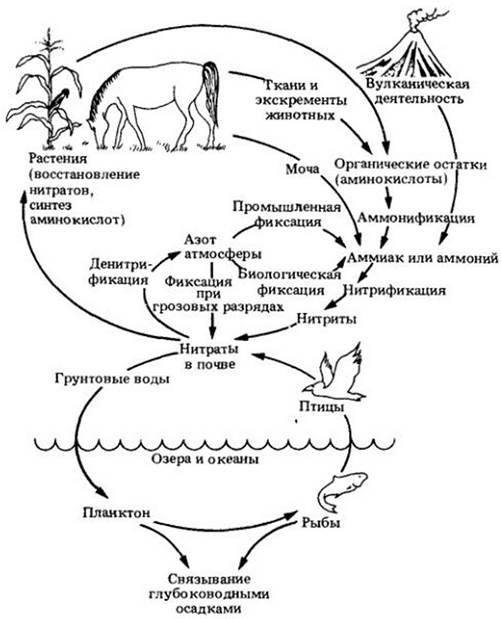
Азот в доступной форме повсеместно является основным лимитирующим питательным элементом для роста сельскохозяйственных растений. Процессы, благодаря которым азот циркулирует из атмосферы через растения и почву при участии живых организмов, составляют круговорот азота и показаны на рис. 26-8.
Рис. 26-8. Круговорот азота
Аммонификация
Большая часть азота поступает в почву из мертвого органического материала, представляющего собой сложные органические соединения, такие, как белки, аминокислоты, нуклеиновые кислоты и нуклеотиды. Эти азотистые соединения, как правило, быстро разлагаются на более простые живущими в почве гнилостными бактериями и различными грибами, которые включают азот в аминокислоты и белки, а его избыток выделяют в форме иона аммония (NН+4); этот процесс называют аммонификацией. Азот может выделяться в виде аммиака (NH3), но обычно это происходит только при разложении большого количества богатого азотом материала, например, в навозных или компостных кучах. Как правило, аммиак, получающийся в процессе аммонификации, растворяется в почвенной воде, где соединяется с протонами и образует ионы аммония.
Нитрификация
Некоторые виды почвенных бактерий способны окислять аммиак или ионы аммония. Окисление аммиака, или нитрификация, — это процесс, дающий энергию. Высвобождающаяся энергия используется этими бактериями для восстановления углекислоты приблизительно так же, как фотосинтезирующие автотрофы используют для этой цели энергию света. Такие организмы называются хемосинтезирующими автотрофами (в отличие от фотосинтезирующих автотрофов). Например, хемосинтезирующая нитрифицирующая бактерия Nitrosomonas окисляет аммиак до нитритов (NO-2):
2NН3 + 3O2 —> 2NO-2 + 2Н+ + 2Н2O.
Нитриты токсичны для высших растений, но они редко накапливаются в почве. Другой род бактерий, Nitrobacter, окисляет нитриты до нитратов (NO-3) снова с высвобождением энергии:
2NO-2 + О2 —> 2NО-3.
Практически весь азот поглощается растениями в форме нитратов.
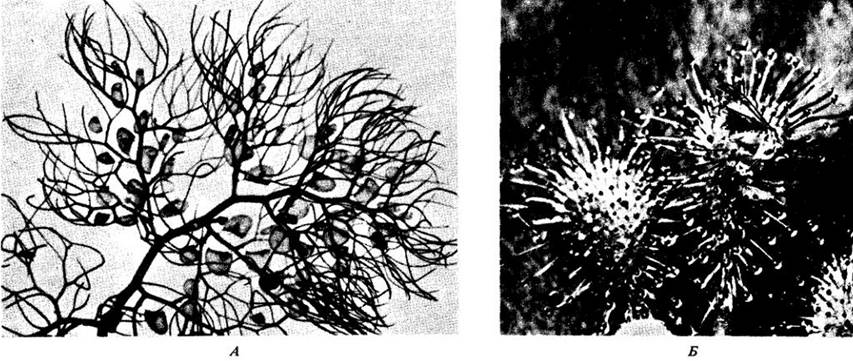
Некоторые виды растений могут использовать в качестве источника азота белки животных. Эти насекомоядные растения (рис. 26-9) имеют специальные приспособления для привлечения и ловли насекомых и других очень мелких животных. Растения переваривают пойманные организмы и всасывают содержащиеся в них азотистые соединения и другие органические и минеральные продукты, такие, как калий и фосфаты. Большинство насекомоядных растений всего мира живет на болотах, где субстрат слишком кислый и поэтому нс подходит для обитания нитрифицирующих бактерий.
Рис. 26-9. Насекомоядные растения ловят жертву различными способами. А. Пузырчатка обыкновенная (Utricularia vulgaris) — свободноплавающее водное растение. Ловушками служат маленькие уплощенные грушевидные пузырьки. Каждый пузырек имеет «ротовое отверстие», прикрытое висячей «дверцей». Ловчий механизм состоит из четырех жестких щетинок, расположенных у свободного нижнего края дверцы. Когда мелкое животное прикасается к щетинкам, волоски отгибают нижний конец дверцы, и она открывается. Вода устремляется в пузырек, неся с собой животное, и дверца за ним захлопывается. Ферменты, секретируемые внутренней поверхностью пузырька и обитающими в нем бактериями, переваривают добычу, а образовавшиеся минеральные и органические вещества поглощаются клеточными оболочками ловушки. Непереваренные наружные скелеты остаются в пузырьках. Б. Росянка средняя (Drosera intermedia) — маленькое растение, чаще всего достигающее нескольких сантиметров в поперечнике, — имеет булавовидные волоски на верхней поверхности листа. Верхушки этих железистых волосков выделяют прозрачную липкую слизь, которая привлекает насекомых. Когда насекомое попадает в слизь, волоски наклоняются навстречу добыче и лист постепенно смыкается вокруг нее. Известно, что волоски секретируют по крайней мере 6 ферментов, которые вместе с ферментами, выделяемыми бактериями, переваривают насекомое. Особое значение при этом имеет хитиназа. Образовавшиеся питательные вещества из слизи всасываются теми же железками, что и вырабатывают пищеварительные ферменты (см. также приложение 3)
Ассимиляция азота
Когда нитраты попадают в растительную клетку, они восстанавливаются до ионов аммония. В отличие от нитрификации, включающей окисление NH+4 и выделение энергии, процесс восстановления требует энергетических затрат. Ионы аммония, образующиеся в результате восстановления, переносятся на углеродсодержащие соединения с образованием аминокислот и других органических веществ, содержащих азот. Этот процесс называется аминированием. Включение азота в органические соединения происходит главным образом в молодых, растущих клетках корня. Таким образом, начальные этапы метаболизма азота протекают в корне — почти весь азот, поднимающийся по стеблю по ксилеме, уже входит в состав органических веществ, главным образом — молекул аминокислот.
Синтез аминокислот
Аминокислоты синтезируются из ионов аммония и кетокислот, которые обычно являются продуктами метаболического расщепления сахаров. Основная аминокислота, образующаяся подобным образом, — глутаминовая, главный переносчик азота в растении. Из аминокислоты, возникшей путем аминирования кетокислоты, синтезируются другие аминокислоты в ходе процесса переаминирования — переноса аминогруппы (-NH2) одной аминокислоты на кетокислоту с образованием другой аминокислоты.
Растения путем аминирования или переаминирования могут синтезировать все необходимые им аминокислоты, используя неорганический азот. Животные синтезируют только некоторые из необходимых аминокислот, а остальные должны получать с пищей. Таким образом, животные полностью зависят от растений, которые снабжают их не только углеводами, но и белками.
Другие соединения, содержащие азот
К важным органическим соединениям, содержащим азот, относятся нуклеотиды (такие, как АТР, ADP, NAD и NADP), хлорофилл и другие органические молекулы с порфириновыми кольцевыми структурами, а также нуклеиновые кислоты ДНК и РНК. Азот содержат многие витамины, например, витамины группы В. Все они, как и аминокислоты, могут синтезироваться растениями, а животные должны получать их с растительной пищей.
Потери азота
Как уже указывалось, содержащие азот соединения, входящие в состав растений, возвращаются в почву после гибели растений (или животных, питавшихся растениями) и перерабатываются почвенными организмами. Затем нитраты, растворенные в почвенной воде, поглощаются растениями и вновь превращаются в органические соединения. В этом цикле всегда «теряется» некоторое количество азота, т. с. он становится недоступным для растений определенных экосистем.
Основной источник потерь азота в определенных экосистемах — удаление растений из почвы. В обрабатываемых почвах часто наблюдается постоянное уменьшение содержания азота. Азот теряется и в тех случаях, когда верхний слой почвы уносится в результате эрозии или повреждается пожаром. Азот теряется и вследствие выщелачивания. Нитраты и нитриты, будучи анионами, легко вымываются из корнеобитаемой зоны водой, просачивающейся сквозь почву.
В анаэробных условиях нитраты часто восстанавливаются до газообразных форм — молекулярных азота (N2) и закиси азота (N2О), которые возвращаются в атмосферу. Этот процесс восстановления, называемый денитрификацией, осуществляется многочисленными микроорганизмами. Низкое содержание кислорода, которое является необходимым условием денитрификации, характерно для переувлажненных почв и таких биотопов, как болота. Поступление легко разлагающегося органического материала обеспечивает денитрифицирующие бактерии источником энергии и при прочих подходящих условиях способствует денитрификации.
Фиксация азота
Если бы азот, который выносится из почвы, постоянно в нее не возвращался, жизнь на Земле постепенно бы прекратилась. Запас азота в почве пополняется за счет фиксации азота.
Фиксация азота — это процесс, в котором N2 восстанавливается до NH+4 и становится доступным для реакции аминирования. От фиксации азота, которую в значительной мере могут осуществлять только некоторые бактерии, зависит в настоящее время существование всех живых организмов (в той же степени, в какой существование большинства из них зависит от фотосинтеза как источника энергии).
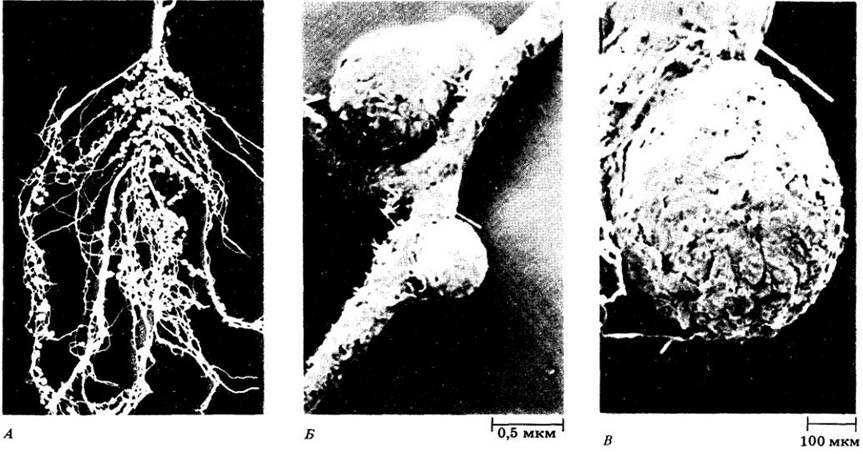
Среди разных классов азотфиксирующих организмов особое место по количеству фиксируемого азота принадлежит симбиотическим бактериям. Наиболее распространенной азотфиксирующей бактерией является Rhizobium, которая поселяется на корнях бобовых — люцерны (Medicago saliva), клевера (Trifolium), гороха (Pisum sativum), сои (Glycine mах),фасоли (Phaseolus) и т. д. (рис. 26-1 и 26-10).
Рис. 26-10. А. Фиксирующие азот клубеньки на корнях сои (Glycine mах), образующиеся в результате симбиотических взаимоотношений между корневыми
клетками этого растения семейства бобовых и бактериями рода Rhizobium. Б. и В. Микрофотографии клубеньков клевера ползучего (Trifolium repens), сделанная с помощью сканирующего электронного микроскопа
Благотворное влияние бобовых на почву известно с древности. Теофраст, живший в III в. до н. э., писал, что греки выращивали бобы (Vicia faba) для обогащения почвы. Там, где растут бобовые, в почву может выделяться некоторый «избыток» азота, который становится доступным для других растений. В современном сельском хозяйстве обычно практикуется севооборот со сменой зерновых, например, кукурузы (Zea mays), и бобовых, например, люцерны. При этом бобовые скашивают, оставляя в почве их богатые азотом корни или, еще лучше, растения просто запахивают в землю. Запаханный урожай люцерны может добавить от 300 до 350 кг азота на гектар почвы. По скромным подсчетам подобные биологические системы за год вносят в землю от 150 до 200 млн. т азота.
Промышленный способ химической фиксации атмосферного азота был разработан в 1914 г. С этого времени коммерческое производство фиксированного азота постоянно возрастало вплоть до настоящего уровня — около 50 млн. т в год. Большая часть этого азота используется в качестве удобрений. К сожалению, промышленная фиксация требует больших энергетических затрат.
Небольшое количество азота связывается при грозовых разрядах и увлекается осадками на Землю. Иногда дождевая вода приносит аммиак и окислы азота, которые попали в атмосферу. Измерения, проводимые на опытной станции в Англии более пяти лет, показали, что с дождевой водой в почву ежегодно попадает 7,1 кг азота на гектар.
Фиксирующие азот симбиозы
В симбиотической ассоциации бактерий рода Rhizobium и представителей семейства бобовых растения снабжают бактерии соединениями углерода, служащими источником энергии для фиксации азота и других реакций метаболизма, а также создают им защищенную среду обитания. В свою очередь растения получают азот в той форме, в которой он используется для синтеза белка.
Бактерии рода Rhizobium (ризобии, рис. 26-11, А) проникают в корневые волоски растений семейства бобовых на стадии проростков. Симбиотические отношения между Rhizobium japonicum и соей (Glycine mах) начинаются с прикрепления ризобий к растущим корневым волоскам (рис. 26-12, А). Многие из этих корневых волосков деформируются и искривляются. После прикрепления ризобии проникают в клеточные оболочки корневого волоска. Разрушение клеточной оболочки в месте внедрения происходит, вероятно, с помощью выделяемых бактериями ферментов.
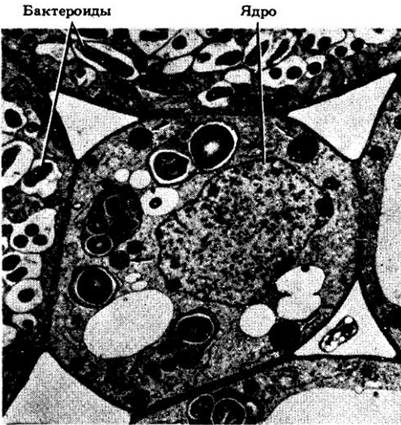
Рис. 26-11. Rhizobium. А. Свободноживущая форма бактерий. Б. Бактероидная форма ризобий, проникших в клетки корня
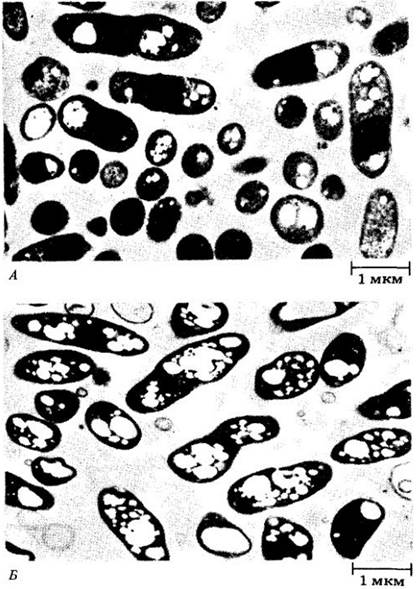
По мере проникновения бактериальной клетки в корневой волосок он прекращает удлиняться и начинает расти в месте внедрения, где откладывается дополнительное количество материала клеточной оболочки. В результате направленного врастания этого материала вглубь корневого волоска образуется трубчатая структура, которая называется инфекционной нитью (рис. 26-12, Б). В один корневой волосок могут проникнуть несколько ризобий. Попав в корневой волосок, бактерии продвигаются по инфекционным нитям сначала к его основанию, затем через клеточные оболочки в клетки коры корня. Бактериальный симбионт вызывает деление клеток в отдельных участках коры, в которую он попадает в результате роста и ветвления инфекционных нитей (рис. 26-12, Б, Г и 26-13). Выход ризобий из инфекционной нити во вместилища, образуемые плазматической мембраной корневого волоска растения-хозяина, а также продолжающееся деление бактероидов (так называются увеличившиеся в размерах ризобии, попавшие внутрь клеток корня, рис. 26-11, В) и коровых клеток корня приводят к образованию опухолеподобных клубеньков (см. рис. 26-10). Способы инфицирования и формирования клубеньков на корнях других бобовых примерно те же, что и у сои.
Рис. 26-12. Ранние этапы инфицирования сои бактериями Rhizobium japoniсum. А. На микрофотографии, полученной с помощью сканирующего электронного микроскопа, видны бактерии (указаны стрелками), прикрепившиеся к недавно появившемуся корневому волоску. Б. На микрофотографии, полученной с помощью дифференциально-интерференционного контраста, показан короткий искривленный корневой волосок, содержащий многочисленные инфекционные нити (указаны стрелками). В. Поперечный срез инфицированного корня, на котором видна ранняя стадия развития меристемы клубенька. На этой стадии большинство клеточных делений в коре направлено антиклинально (под прямым углом к поверхности). В клубеньковой меристеме, прилежащей к инфицированному корневому волоску, клетки делятся и в других направлениях. Г. Большее увеличение поперечного среза, на котором видны инфицированный корневой волосок и прилегающая меристема клубенька. Инфекционные нити (указаны стрелками) можно видеть в корневом волоске и в меристеме
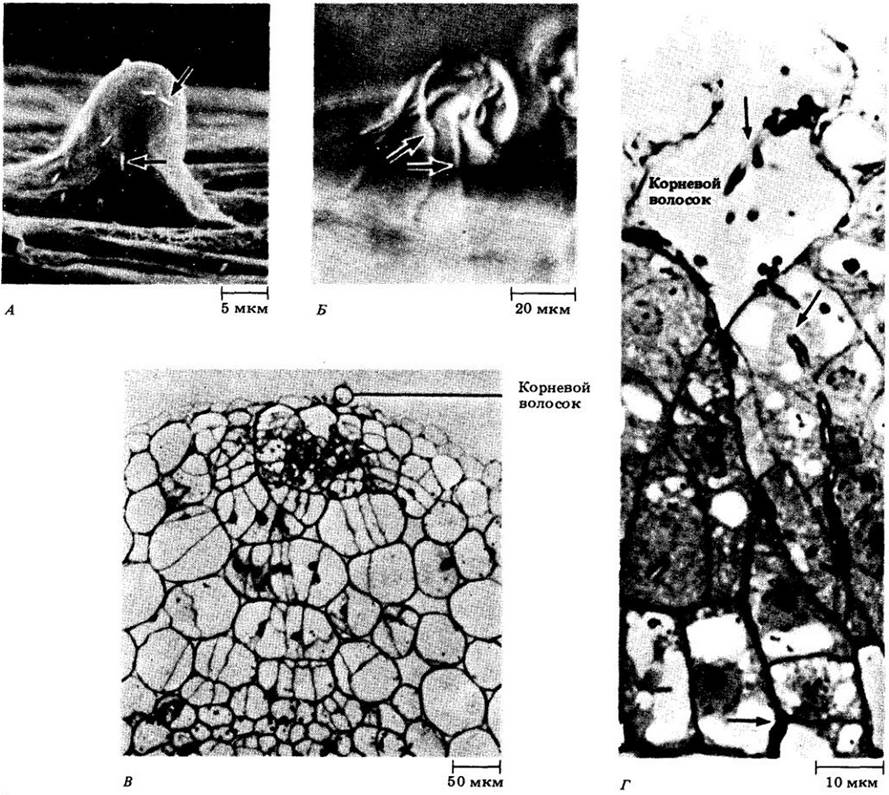
Рис. 26-13. Электронная микрофотография разветвленной инфекционной нити, содержащей бактероиды, в зараженной клетке клубенька сои
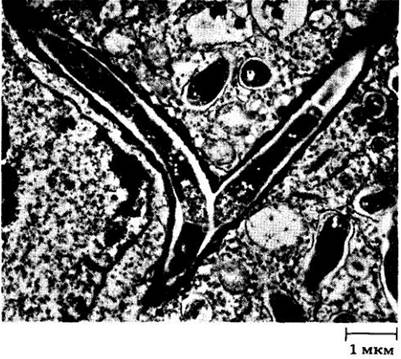
Корневые клубеньки состоят отчасти из увеличенных клеток, инфицированных ризобиями, и отчасти — из множества расположенных между ними более мелких незараженных клеток (рис. 26-14). До недавнего времени считалось, что незараженные клетки не имеют существенного значения в метаболизме свежефиксированного азота, а выполняют только опорные функции в клубеньке. Сейчас, однако, ясно, что у сои эти неинфицированные клетки играют основную роль в образовании уреидов (производных мочевины) из фиксированного зараженными клетками азота.
Рис. 26-14. Электронная микрофотография зрелого корневого клубенька сои. Вакуолизированные незараженные клетки видны среди темных инфицированных меток
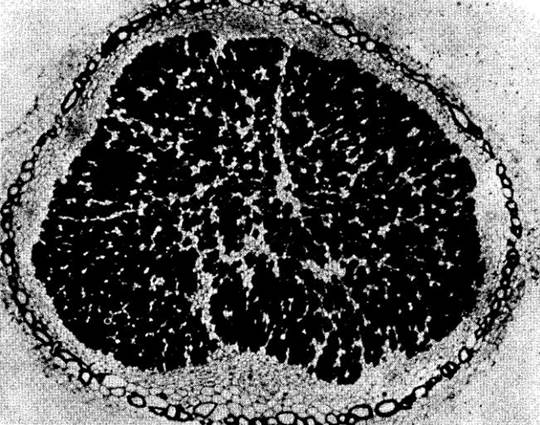
Бобовые можно разделить на несколько групп в зависимости от природы азотсодержащих веществ, поступающих из клубеньков в надземные части растения. Например, вика, люпин и горох относятся к растениям, у которых фиксированный азот переносится главным образом в форме аминокислоты аспарагина. Фасоль, вигна и бобы, с другой стороны, образуют группу растений, у которых для экспорта азота из клубеньков служат уреиды. Их синтез начинается в инфицированных клетках и заканчивается в незаряженных.
В незаряженных ризобиями клетках клубенька образуются крупные микротельца (пероксисомы) и обильный трубчатый эндоплазматический ретикулум (рис. 26-15). По-видимому, в этих клеточных органеллах содержатся ферменты, принимающие участие в заключительных стадиях образования уреидов.
Рис. 26-15. Электронная микрофотография участка клубенька сои с незараженной клеткой (в середине), примыкающей к зараженным меткам (сверху и слева). Инфицированные метки содержат множество бактероидов. В неинфицированной клетке видны многочисленные микротельца (указаны стрелками). Количество митохондрий в инфицированных метках в четыре раза больше, чем в незараженных, по-видимому, вследствие огромной потребности бактероидов в АТР
Симбиоз между видами Rhizobium и бобовых удивительно специфичен; например, бактерии, которые вызывают образование клубеньков на корнях клевера, не образуют клубеньков на корнях фасоли. В настоящее время на рынок поступают специфические штаммы Rhizobium, выращиваемые в лаборатории. Добавляя к семенам во время сева определенные бактерии, фермеры могут обеспечить образование эффективных ассоциаций для фиксации азота в культуре; бобовых. Исследуется механизм взаимодействия бактерий Rhizobium с корнями определенного вида бобовых. Бактерии и растение могут узнавать друг друга с помощью растительного белка лектина, находящегося на поверхности корней. Лектин связывается с полисахаридами на поверхности клеток «своего» вида Rhizobium и не взаимодействует с полисахаридами «чужого» вида.
Бобовые — самая большая группа растений, образующих «товарищество» по фиксации азота с симбиотическими бактериями. Однако существует несколько фиксирующих азот симбиозов, в которых участвуют другие растения. Например, на корнях ольхи поселяются и вызывают образование клубеньков не Rhizobium, а азотфиксирующие актиномицеты. Мирт болотный (Myrica gale), представители родов Соmptonia и Ceanothus тоже вступают в симбиотические отношения с актиномицетами (см. гл. 11).
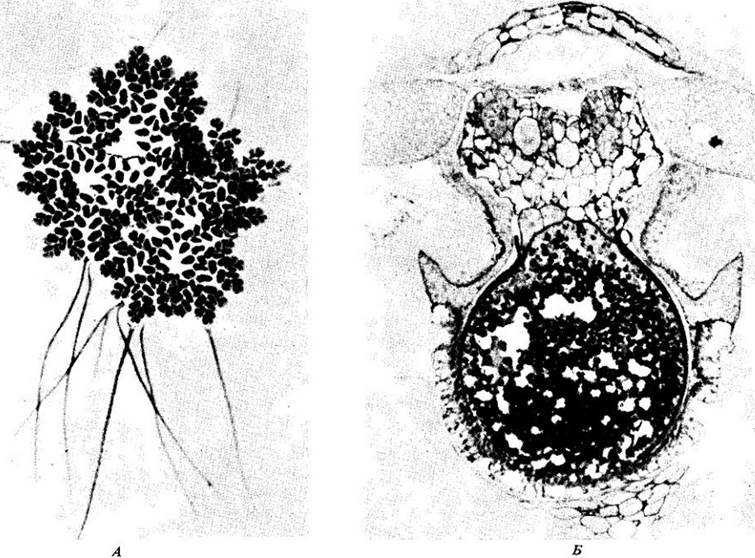
В некоторых частях света большое практическое значение имеет другой симбиотический союз: Azolla — мелкий плавающий папоротник и Anabaena — азотфиксирующая цианобактерия, которая живет в «кармашках» на его листьях (рис. 26-16). Azolla, инфицированная цианобактерией, может давать до 50 кг азота на гектар. Например, на Дальнем Востоке зарослям Azolla-Anabaena позволяют развиваться на рисовых полях (см. также рис. 11-12, Б). Растения риса в конечном результате затеняют Azolla, папоротник отмирает, азот высвобождается и используется рисом.
Рис. 26-16. А. Azolla caroliniana — водный папоротник, растущий в симбиозе с цианобактерией Аnabaena. Б. Симбиоз Azolla-Anabaena уникален среди других
азотфиксирующих симбиозов тем, что взаимоотношения поддерживаются в течение всего жизненного цикла хозяина. Здесь можно видеть нити Аnabаеna,
связанные с женским гаметофитом (мегагаметофитом), который развился из проросшей мегаспоры
Свободноживущие фиксирующие азот микроорганизмы
Фиксировать азот могут несимбиотические бактерии родов Azotobacter и Clostridium. Azotobacter — аэроб, Clostridium — анаэроб; оба — почвенные сапробионты. Подсчитано, что они добавляют около 7 кг азота на гектар почвы в год. Другая важная группа включает многие фотосинтезирующие микроорганизмы, такие, как цианобактерии.
Различия между фиксацией азота свободноживущими и существующими в симбиотических ассоциациях бактериями не всегда отчетливы. Например, аэробная азотфиксирующая бактерия Azotobacter, как правило, концентрируется вокруг корней некоторых злаков, таких, как сахарный тростник (Saccharum officinarum), и играет существенную роль в снабжении растений азотом. Злаки (и другие растения), вероятно, выделяют органические вещества, которые служат источниками энергии для «свободноживущих» бактерий. Ассоциация, таким образом, отчасти соответствует симбиотической.
Фермент, который осуществляет фиксацию азота, называется нитрогеназой. Нитрогеназа содержит молибден, железо и сульфидные простетические группы, поэтому указанные элементы необходимы биологическим системам для осуществления фиксации азота. Нитрогеназа использует и большое количество АТР в качестве источника энергии, что делает метаболический процесс азотфиксации энергетически дорогим.
Помимо восстановления N2 до NH+4нитрогеназа превращает ацетилен — молекулу с тройной связью в этилен — молекулу с двойной связью. Превращение ацетилена в этилен — пример смены субстратов фермента — распространенный тест на нитрогеназную активность.
Хотя растения, как указывалось выше, не нуждаются в кобальте как в микроэлементе, кобальт нужен микроорганизмам, фиксирующим азот. Поэтому кобальт, нс являясь составной частью нитрогеназы, необходим для осуществления симбиотической фиксации азота.
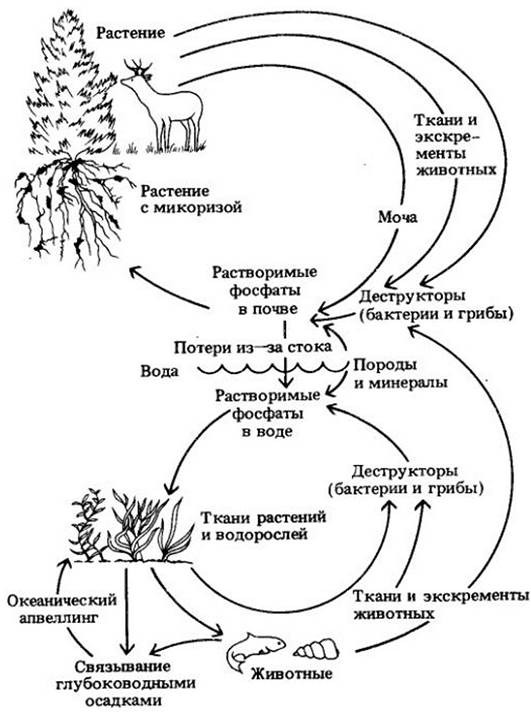
Круговорот фосфора
Круговорот фосфора (рис. 26-17) проще, чем круговорот азота, поскольку включает меньше стадий, среди которых к тому же нет зависящих от специфических групп микроорганизмов. В круговороте фосфора в отличие от круговорота азота первичным резервуаром служит не атмосфера, а земная кора. Как, указывалось ранее, основная масса фосфора попадает в почвенный раствор в результате длительного процесса выветривания пород и минералов.
По сравнению с азотом количество фосфора, требующееся растениям, относительно невелико (см. табл. 26-1 и 26-2). Тем нс менее среди элементов, для которых земная кора является первичным резервуаром, именно фосфор обычно ограничивает рост растений. В Австралии, например, где почвы крайне выветрены и бедны фосфором, распространение и границы естественных растительных сообществ часто определяются доступностью почвенных фосфатов.
Фосфор циркулирует от растений к животным и возвращается в почву в форме органических остатков и отходов. Органические соединения фосфора превращаются в неорганические фосфаты и таким образом становятся снова доступными растениям (рис. 26-17).
Рис. 26-17. Круговорот фосфора
В результате эрозии, загрязнений, а также со сточными водами большое количество фосфора попадает в реки и ручьи. В итоге фосфор оказывается в океане, где откладывается на дне в виде осадков. (Большое количество фосфора осаждается и при разложении погибших организмов.) В прошлом использование гуано (отложения помета морских птиц) в качестве удобрения возвращало некоторое количество фосфора из океана в наземные экосистемы. Однако большая часть фосфора из океанических осадков станет доступна только в результате крупных поднятий морского дна. Чтобы возместить эти потери, в больших масштабах разрабатываются залежи горных пород, содержащих фосфаты, для получения удобрений.
Воздействие человека на круговорот питательных элементов
Для нормального круговорота фосфора, азота и других веществ требуется постоянный транспорт элементов от одной стадии цикла к другой для того, чтобы ни на одной из них не происходило накопления или потерь. В течение миллионов лет организмы получали нужное количество необходимых неорганических питательных элементов благодаря их нормальному круговороту. Однако в последнее время деятельность человека оказывает серьезное влияние на круговорот некоторых веществ, что иногда приводит к их чрезмерному накоплению или потерям на одной из специфических стадий. Например, почвенная эрозия ускоряет вынос фосфора из почвы. Кроме того, сточные воды попадают в водотоки быстрее, чем содержащийся в них фосфор может вступить в круговорот, и в результате этого он попадает в океан и там теряется.
Для нормального круговорота азота необходимо поддержание баланса между процессами фиксации, извлекающими азот из атмосферного резервуара, и реакциями денитрификации, которые возвращают азот в атмосферу. За последнее время во внешнюю среду поступило огромное количество фиксированного азота (нитратов) благодаря усиленному использованию удобрений. Поскольку азот в форме нитратов легко вымывается из почвы, возросло загрязнение азотистыми веществами грунтовых вод, озер и водотоков. Проблема усложняется тем, что болота и затопляемые почвы, где наиболее активно протекает денитрификация, с опасной быстротой уничтожаются, превращаясь в места застройки, сельскохозяйственные угодья или свалки.
Для уменьшения потерь нитратов в почве предложено решение, которое могло бы стать распространенным земледельческим приемом. Речь идет о применении специального органического соединения, которое избирательно подавляет активность нитрифицирующей бактерии Nitrosomonas на определенный срок, а затем разрушается в почве. Этот прием помог бы сохранить большую часть азота удобрений в форме аммония до тех пор, пока его не усвоили растения. Однако, как это часто бывает при вмешательстве в природные процессы, и здесь возможны нежелательные последствия. Например, аммиак в высокой концентрации весьма токсичен для некоторых сельскохозяйственных растений.
Почва и земледелие
В естественных условиях имеющиеся в почве элементы рециркулируют и снова становятся доступными для растений. Как ранее указывалось, отрицательно заряженные глинистые частицы могут связывать такие положительно заряженные ионы, как Са2+, и К+. Эти ионы поглощаются корнями растений либо непосредственно, либо после того, как перейдут в почвенный раствор. Обычно в плодородной почве много требующихся растению катионов, а их количество, выносимое с одним урожаем, невелико. Однако, когда с поля снимается один урожай за другим и необходимые элементы постоянно изымаются из круговорота, содержание некоторых катионов (чаще всего калия) может настолько уменьшиться, что становится необходимым внесение удобрений, содержащих дефицитный элемент.
Программы снабжения сельскохозяйственных и садовых растений дополнительными питательными веществами должны скрупулезно учитывать количество веществ, необходимых для формирования урожая определенной культуры, и количество этих веществ, доступных растению из всех источников. Часто почва и растительные остатки не могут предоставить необходимые питательные элементы в полном объеме, и тогда требуется обеспечить их дополнительное количество путем внесения минеральных удобрений, органических остатков (таких, как компост) или их комбинаций.
Азот, фосфор и калий — три элемента, которые обычно включаются в минеральные удобрения. Удобрения, как правило, обозначают формулами, указывающими процентное содержание каждого из этих элементов. Например, 10 — 5 — 5 означает, что удобрение содержит 10% азота (N), 5% пятиокиси фосфора (Р2O5) и 5% окиси калия (К2O).
Другие необходимые неорганические питательные элементы, хотя и требуются в очень малых количествах, иногда становятся лимитирующими факторами в почве, на которой растут культурные растения.
Исследование питания растений
Изучение минеральных элементов, необходимых для сельскохозяйственных растений — в особенности изучение их количества, требующегося для высокого урожая, и исследование способности различных почв поставлять эти элементы, — имеют огромное практическое значение для полеводства и садоводства. Из-за постоянно растущих потребностей человечества в пище исследования такого рода без сомнения будут необходимы и в будущем.
Обедненность и токсичность почв
Мероприятия по улучшению почвы — внесение питательных веществ с удобрениями, увеличение pH известкованием или удаление избытка солей промыванием водой — не исчерпывают числа приемов, увеличивающих и обеспечивающих урожаи на малоплодородных почвах. Использование теории и методов растениеводства и физиологии питания растений делает возможным выведение сортов сельскохозяйственных растений, которые лучше других приспособлены к росту в условиях недостатка питательных веществ. Плодотворность этого направления исследований подтверждается существованием дикорастущих растений на почвах, весьма отличных от тех, на которых растут обычно сельскохозяйственные растения. Примером могут служить кислые сфагновые болота с pH менее 4,0 или отвалы рудников, где часто содержатся высокие концентрации потенциально токсичных металлов, таких, как цинк и никель.
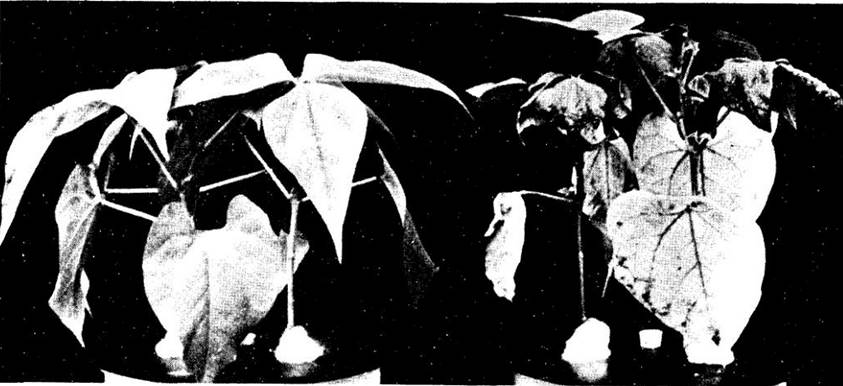
Недавно были проведены исследования по выведению фасоли (Phaseolus vulgaris), устойчивой к дефициту калия. Образцы фасоли были собраны по всему миру и выращивались на питательном растворе, в котором калия было меньше, чем требуется для оптимального роста. Остальные питательные вещества находились в оптимальных концентрациях. Крайние примеры реакции фасоли на стресс, созданный недостатком калия, показаны на рис. 26-18. Линия 58 (слева) растет почти нормально; линия 63 (справа) демонстрирует симптомы острой недостаточности калия. Выдающегося успеха в этой относительно новой области исследований достигли Э. Эпстейн и его сотрудники из Калифорнийского университета в Дейвисе, выделив линии ячменя (Hordeum sativum), устойчивые к высокой концентрации солей. Наиболее устойчивые росли достаточно хорошо даже при поливе морской водой.
Рис. 26-18. Сравнение симптомов недостаточности калия у опытных образцов фасоли (Phaseolus vulgaris). Слева линия, устойчивая к дефициту калия, справа — явно неустойчивая линия
Эффективность фиксации азота
Вмешательство в биологическую фиксацию азота таит огромные возможности повышения эффективности его использования. Одно из направлений этих исследований касается улучшения работы ассоциации Rhizobium-бобовые, например, за счет генетического отбора растений и бактерий для выявления таких комбинаций, которые увеличили бы фиксацию в определенных условиях среды. Этого можно было бы достичь, увеличив эффективность фотосинтеза бобовых, что привело бы к увеличению количества углеводов для бактериальной фиксации азота и роста. Необходимо учесть, однако, что фиксация азота требует много энергии и что любое увеличение фиксации пойдет за счет продуктивности побегов.
Другой подход к проблеме состоит в создании новых и более эффективных ассоциаций свободноживущих фиксирующих азот бактерий с высшими растениями. В Бразилии в начале 1970-х гг. были обнаружены несколько типов азотфиксирующих бактерий, живущих в ассоциации с корнями некоторых тропических трав. Например, злак Digitaria поддерживает существование популяций бактерий Azospirillum. Подобные ассоциации были найдены у важнейших сельскохозяйственных культур — кукурузы и сахарного тростника. Несмотря на то что практическая выгода могла быть огромной, эффективность ассоциаций азотфиксирующих бактерий с такими злаками, как кукуруза, все еще не выяснена.
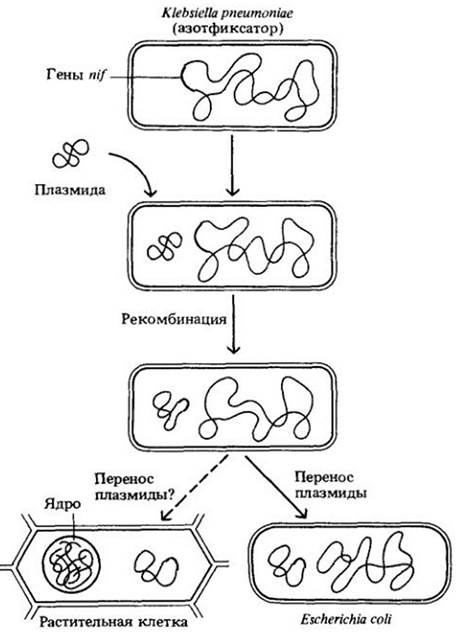
Среди исследовательских подходов, использующих наиболее изощренные методы молекулярной биологии, по-видимому, самым успешным является связанный с генетическими модификациями и переносом генов, ответственных за фиксацию азота, от одного организма к другому. Перенос соответствующих генов уже осуществлен. Кластер генов, обеспечивающих фиксацию азота, от бактерий Klebsiella pneumoniae был перенесен в Escherichia coli. Пересадка генов была произведена путем их включения в плазмиду (см. с. 248) и введения этой плазмиды в клетку Е. coli (рис. 26-19). Генетически измененная Е. coli фиксировала азот в определенных условиях.
Теоретически гены фиксации азота можно было бы пересадить нефиксирующим азот видам, например, кукурузе, но одновременно возникает множество проблем, таких, как защита фиксирующего азот фермента от кислорода, который подавляет фиксацию. Поэтому исследователи в этой области отнюдь не уверены, что эффективная операция подобного рода может быть легко осуществлена.
Рис. 26-19. Перенос ответственных за азотфиксацию генов (генов nif, от англ, nitrogen-fixation) от бактерии, фиксирующей азот, к другому организму. Гены nif от Klebsiella pneumoniaeбыли встроены в плазмиду и затем перенесены в Escherichia coli — бактерию, которая не может фиксировать азот. Несмотря на успешный первый шаг, перенос генов еще не гарантирует, что реципиент будет фиксировать азот. Например, генетически модифицированная Е. coli вырабатывала фермент нитрогеназу, необходимую для фиксации азота, однако могла фиксировать азот только в таких экспериментальных условиях, когда фермент был защищен от кислорода. Иными словами, среди «новых» способностей у бактерий отсутствовала способность защищать ключевой фермент от окисления. Теоретически азотфиксирующая способность может быть передана растениям, не способным к фиксации, с помощью плазмид
Влияние загрязнения
Токсические эффекты разнообразных неорганических агентов, загрязняющих среду, привлекают внимание современных исследователей. На сельскохозяйственные растения, например, могут неблагоприятно действовать тяжелые металлы, такие, как медь и кадмий. Особенно страдают водные экосистемы, куда сбрасываются промышленные и коммунальные отходы. Поступление в пресноводные экосистемы азота и фосфора — основных элементов эвтрофизации — приводит к бурному росту водорослей и водных цветковых растений, что значительно снижает ценность озер и рек как мест отдыха.
Широко распространены повреждения как наземных, так и в особенности водных экосистем, вызванные кислотным дождем. Кислотный дождь является результатом взаимодействия двуокиси серы и окислов азота — продуктов сгорания ископаемого топлива — с атмосферной влагой, что приводит к образованию серной и азотной кислот. Они и создают высокую степень кислотности дождя. В некоторых районах Скандинавии, на северо-востоке и Среднем Западе США, на юго-востоке Канады дождевая вода обычно имеет pH от 4,0 до 4,5, а иногда и ниже 4,0. В то же время pH дождевой воды,
находящейся в равновесии с углекислотой в незагрязненной атмосфере, равен приблизительно 5,6. Кислотный дождь неблагоприятно влияет на растения, на выветривание пород и минералов, на растворимость потенциально токсичных металлов и на здоровье людей. От него страдают сотни озер с мягкой водой в Скандинавии, Канаде и США. В этих озерах нет буферной системы, создаваемой бикарбонатами и карбонатами, которые нейтрализуют кислоты в озерах с жесткой водой. Мягкая вода характерна для озер высокогорий, например, гор Адирондак на северо-западе США и других мест, где тонкий слой почвы лежит на магматических породах. Увеличение кислотности воды, происходящее сейчас в таких озерах, может губительно сказаться на размножении рыбы (см. приложение к гл. 22).
Заключение
Всего 16 химических элементов требуется большинству растений для нормального роста. Из них углерод, водород и кислород поступают из воздуха и воды. Остальные поглощаются корнями в форме ионов. Эти 16 элементов делятся на макро- и микроэлементы в зависимости от того, в каком количестве требуются растению. К макроэлементам относятся углерод, кислород, водород, азот, калий, кальций, фосфор, магний и сера. Группу микроэлементов составляют железо, хлор, медь, марганец, цинк, молибден и бор. Некоторые элементы, такие, как натрий и кобальт, необходимы только отдельным организмам (см. табл. 26-1).
Неорганические питательные элементы выполняют важные функции в клетке. Они регулируют осмос и влияют на проницаемость. Некоторые из них являются структурными компонентами клетки, составными частями ключевых веществ метаболизма, активаторами и коферментами (см. табл. 26-2).
Химические и физические свойства почв определяют их способность поставлять неорганические питательные элементы, воду и создавать другие условия, необходимые для максимальной продуктивности сельскохозяйственных растений. В результате выветривания горных пород и минералов образуются частицы, составляющие неорганическую часть почвы. Все питательные элементы, за исключением азота, образуются в процессе выветривания. Кроме того, почва содержит органическое вещество и поры, заполненные водой и газами в различных соотношениях. На обрабатываемых землях азот, фосфор и калий чаще всего ограничивают рост растений и поэтому вносятся в почву с удобрениями.
Каждый необходимый питательный элемент циркулирует в сложном круговороте между организмами внутри экосистемы и между этими организмами и природными резервуарами элемента. Циркуляция азота через почву, растительные и животные организмы и снова в почву называется круговоротом азота. Азот попадает в почву в форме органических веществ растительного и животного происхождения. Эти соединения разлагаются почвенными организмами. Аммонификация — выделение ионов аммония (NH+4) из азотистых соединений — осуществляется почвенными бактериями и грибами. Нитрификация — окисление аммиака или ионов аммония до нитритов и нитратов. Одни бактерии окисляют аммиак до нитритов, другие ответственны за окисление нитритов до нитратов. Азот усваивается растениями почти исключительно в форме нитратов. Нитраты в растениях восстанавливаются до ионов аммония. Аминокислоты синтезируются в результате аминирования. приводящего к образованию глутаминовой кислоты, или в процессе переаминирования, в котором аминогруппа переносится от аминокислоты на кетокислоту, превращая се в другую аминокислоту. В конечном счете эти органические вещества возвращаются в почву, завершая цикл азота.
Почва теряет азот в результате уборки урожая, эрозии, пожаров, вымывания и в результате активности денитрифицирующих бактерий. Его запасы в почве пополняются за счет фиксации азота, приводящей к включению молекулярного азота в органические соединения. Биологическая фиксация азота осуществляется только бактериями. К ним относятся бактерии рода Rhizobium, живущие в симбиозе с растениями семейства бобовых; свободноживущие бактерии и актиномицеты, вступающие в симбиотические ассоциации с растениями других семейств. В результате сельскохозяйственных работ растения удаляются из почвы, и вследствие этого азот и другие элементы выключаются из естественного круговорота и должны вноситься в почву в форме органических или минеральных соединений.
Приложение 1. Круговорот воды
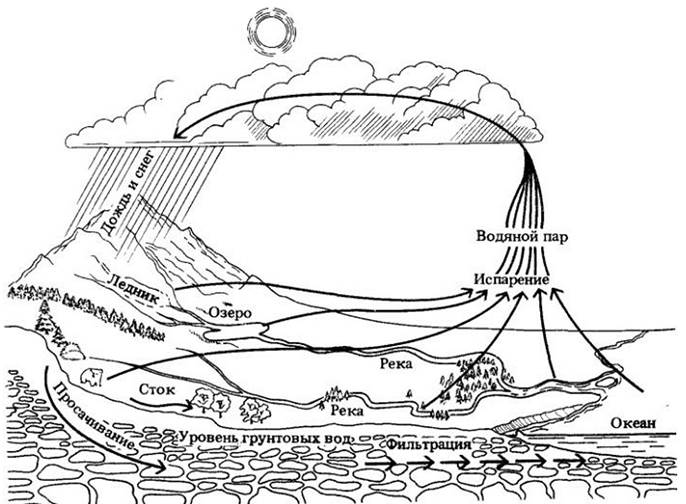
Запас воды на Земле постоянен и используется циклически. Подавляющее количество воды (98%) находится в океанах, озерах и водотоках. Оставшиеся 2% составляют полярные льды и ледники, вода почвы, водяные пары атмосферы и вода, входящая в состав живых организмов.
Под солнечными лучами вода испаряется из океанов, озер, рек и ручьев, с влажной поверхности почвы, из тел живых организмов, поступает в атмосферу и снова возвращается на Землю в виде осадков. Это постоянное перемещение воды с Земли в атмосферу и обратно называется круговоротом воды. Он происходит за счет солнечной энергии.
Часть воды, выпадающей на земную поверхность, просачивается сквозь почву вниз до зоны насыщения. В этой зоне все углубления и трещины в горных породах заполнены водой. Ниже зоны насыщения залегают сплошные горные породы, сквозь которые вода не может проникнуть. Верхняя поверхность зоны насыщения обозначается как уровень грунтовых вод.
Приложение 2. Микориза и питание растений
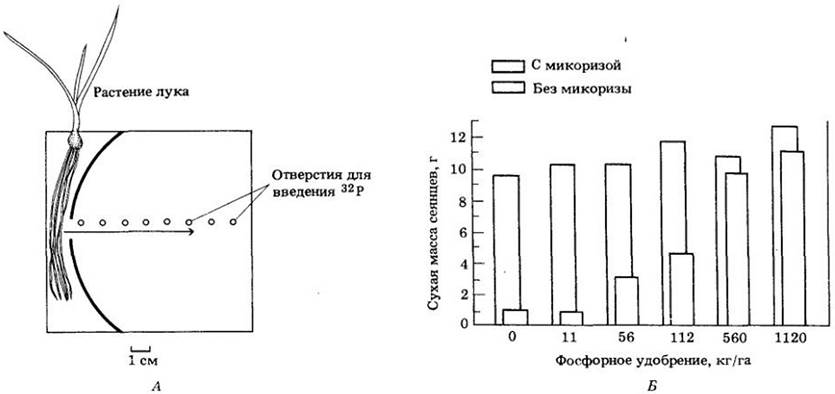
У большинства высших растений поглощение питательных веществ из почвы значительно усиливается, когда на их корнях поселяются грибы-микоризообразователи (см. гл. 16). Особенно велика роль микоризы в поглощении и переносе фосфора. Доказано также, что она способствует поглощению растениями цинка, марганца и меди. Эти элементы в почве слабоподвижны, и поэтому вокруг корневых волосков и корней быстро образуются обедненные ими зоны. Сеть гиф микоризных грибов распространяется на несколько сантиметров от каждого заселенного ими корня и увеличивает таким образом объем эффективно используемой почвы. Способность микоризы поглощать и транспортировать фосфор, находящийся в почве, была показана в опытах с радиоактивным 32Р (А). Помимо увеличения поглощающей поверхности корней значение микоризных грибов заключается и в том. что они могут извлекать фосфор из более разбавленного почвенного раствора и использовать те его источники, которые обычно недоступны растениям.
Увеличение поглощения фосфора с помощью микоризы можно показать, выращивая зараженные и не зараженные ею растения при различном содержании этого элемента в почве (Б). В приведенном в качестве примера опыте проростки цитрусовых, имеющие микоризу, не получая фосфорных удобрений, достигли размера проростков, не имеющих микоризы, но получавших удобрения из расчета 560 кг фосфора на гектар. В этом случае микориза заменила собой фосфорные удобрения, что существенно уменьшило энергетические затраты на получение продукции и ее стоимость.
А. Перемещение 32Р микоризой. Растения лука, зараженные микоризой, выращивались в специально сконструированных камерах, имеющих маленькие отверстия (показаны стрелками), которые позволяют прорастать в отдельный отсек с почвой только грибным гифам, но не корням растения. Затем в почву на разных расстояниях от корня вносили 32Р. Его обнаруживали в корнях и надземных частях растений, имеющих микоризу. В растения лука без микоризы или при нарушении связей с гифами 32Р не проникал. Подавляющее большинство гиф микоризных грибов обнаруживалось на расстоянии 2 — 5 мм от несущего их корня. Б. Ростовая реакция зараженных и не зараженных микоризой сеянцев цитрусовых на внесение разного количества фосфора
Разные виды растений в различной степени зависят от микоризных грибов. У некоторых растений, таких, как цитрусовые, снабжение фосфором в такой степени определяется микоризой, что без нее они могут расти только при очень высоком содержании фосфора в почве (Б). Для многих видов определен процент увеличения роста в присутствии микоризы. Показательны цифры, полученные для пшеницы (220%), кукурузы (122%), лука (3155%), земляники (250%) и туи (962%). При очень высоком содержании растворимого фосфора, когда в почву вносится большое количество фосфорных удобрений, микориза уже не усиливает рост растений. Болес того, обилие удобрений часто подавляет развитие микоризы.
Микоризные грибы присутствуют в большинстве растительных сообществ, однако их количество может уменьшиться или они вообще могут быть уничтожены в результате грубого нарушения почвенного покрова, например, вследствие горных работ. Беспорядочное использование в сельском или лесном хозяйстве фунгицидов против патогенных грибов также может привести к гибели микоризы и как следствие — к торможению роста и гибели растений. Поэтому желательно обработанные ядохимикатами почвы вновь инокулировать микоризными грибами. Эктомикоризные грибы можно культивировать и полученный вегетативный мицелий добавлять в почвенные смеси или в гряды. В юго-восточных районах США и на северо-западном побережье Тихого океана сеянцы хвойных часто заражают особыми видами эктомикоризных грибов для улучшения роста растений на неплодородных почвах. Везикулярно-арбускулярные эндомикоризные грибы пока не удастся выращивать в чистой культуре, поэтому производство их грибницы требует применения «горшечных культур» растения-хозяина. Споры гриба или корни колонизированного грибом растения, полученные из горшечных культур, используют для инокуляции почвы в оранжереях и питомниках. Из-за сложности массового выращивания таких грибов их коммерческое использование весьма ограниченно. При восстановлении растительности на местах горных работ, растения обитавших здесь ранее видов перед посадкой инокулируются в питомниках для улучшения роста и выживания. Заражение эндомико- ризными грибами применяется и в питомниках цитрусовых в южной Калифорнии, и в питомниках лиственных пород на юго-востоке США. При выращивании большинства сельскохозяйственных растений на крупных площадях инокуляция микоризными грибами невыгодна. Однако в производстве наиболее ценных культур и особенно растений, выращиваемых в питомниках, микоризные грибы могут быть очень полезны для улучшения роста без внесения дорогостоящих фосфорных удобрений.
Помимо хорошо доказанного усиления поглощения фосфора на бедных почвах микориза благотворно влияет на рост растений и но другим причинам. К ним относятся повышение устойчивости к почвенным патогенным организмам, крайне кислым почвам и засухе.
Приложение 3. Насекомоядные растения
Насекомоядные растения получают минеральные вещества, включая фиксированный азот, от пойманных животных. Некоторые из них, например, жирянка Pinguicula grandiflora, имеют листья, к поверхности которых приклеиваются мухи и другие мелкие насекомые (А). Ловля насекомых происходит с помощью многочисленных столбчатых желез, расположенных по всему листу. Под сканирующим электронным микроскопом видно, что каждая железа несет округлую капельку слизистого секрета (Б). Это делает лист клейким, как липучка. Когда насекомые прикасаются к каплям секрета, капли вытягиваются и образуют прочные путы, которые можно видеть на снимке вместе с попавшим в них муравьем В). Чем больше бьется насекомое, тем больше желез оно задевает и тем плотнее приклеивается к листу. Среди столбчатых желез имеются и сидячие (Б), которые ничего не выделяют, пока пойманная жертва не дотронется до них. После такой стимуляции сидячие железы выделяют секрет, содержащий ферменты, который быстро образует лужицу вокруг насекомого. Ферменты переваривают жертву, а продукты переваривания накапливаются в жидком секрете. После того как переваривание завершается, жидкость поглощается листом и продукты переваривания распределяются между растущими частями растения.
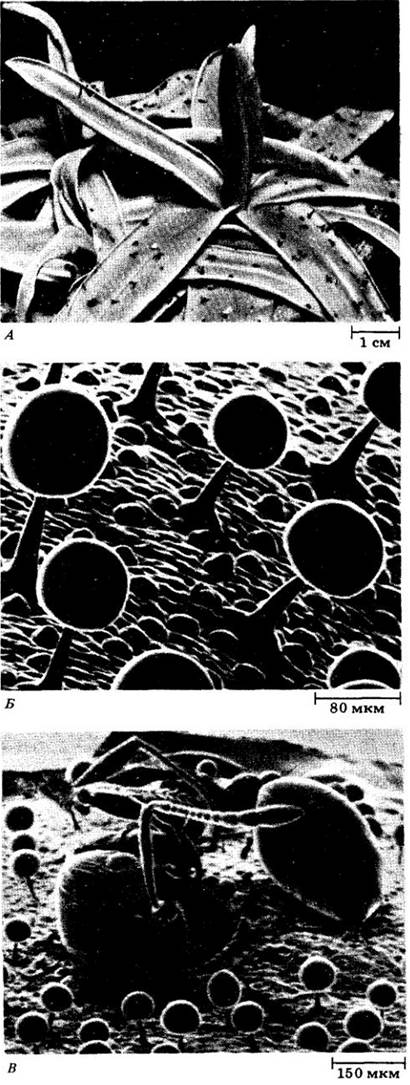

Пищеварительные ферменты синтезируются в сидячих железах. Пока нет стимуляции, ферменты находятся в увеличенных вакуолях и в клеточных оболочках, где их можно обнаружить с помощью соответствующих цитохимических методов. Например, локализация фермента кислой фосфатазы в оболочках клеток головки железы выявляется по распределению темного продукта реакции (Г). Выделение ферментов происходит в результате выброса воды через головку железы. Этот процесс идет за счет выкачивания ионов хлора из нижележащей клетки-резервуара. «Ионные насосы» встроены в мембраны промежуточной клетки, похожей на эндодермальную; они неактивны в бездействующей железе, но быстро включаются стимулом, идущим от жертвы.
Сходные секреторные механизмы обнаружены у других насекомоядных растений, в том числе у венериной мухоловки (Dionaea muscipula) (см. рис. 25-22) и у росянки (Drosera) (см. рис. 26-9).
Приложение 4. Компост
Компостирование — прием старый, как само земледелие, — привлекает повышенный интерес как способ использования органических остатков путем превращения их в удобрение. Исходный продукт — некоторое количество органического материала — листьев, кухонных отходов, навоза, соломы, травы, осадка сточных вод, опилок — плюс популяция бактерий и других обычных микроорганизмов. Единственное требование к условиям — наличие кислорода и влажность. Измельчение органического материала необязательно, но оно увеличивает поверхность, расширяя поле деятельности микроорганизмов, и этим ускоряет процесс.
В компостной куче микроорганизмы быстро растут, генерируя тепло, значительная часть которого сохраняется, поскольку верхние слои органического материала служат изоляцией. Внутри крупной кучи (например, 2 x 2 х 1,5 м) температура поднимается до 70° С, небольшой — обычно до 40°С. С увеличением температуры меняется популяция деструкторов — термофильные и термотолерантные формы сменяют обычные. По мере отмирания исходных форм микроорганизмов их органическое вещество становится частью образующегося продукта. Побочным эффектом подъема температуры служит гибель обычных патогенных бактерий (которые могут присутствовать, например, в осадке сточных вод), а также цист, яиц, и других незрелых форм паразитов растений и животных.
С течением времени в компостной куче изменяется и pH. Его начальное значение около 6, что сравнимо со слабокислой жидкой средой большей части растительного материала. Образование органических кислот на ранних стадиях разложения вызывает дальнейшее подкисление, pH падает до 4.5 — 5,0. Однако с подъемом температуры увеличивается и значение pH, и конечный продукт уже слабощелочной (pH 7.5 — 8,5).
Важным фактором в образовании компоста (как и в любом биологическом процессе, связанном с ростом) является отношение углерода к азоту. Отношение 30 к 1 (по массе) оптимально. Если углерода больше, микробный рост ослабевает. Если больше азота, выделяется аммиак. Если компост слишком кислый, к нему можно добавить известняк (карбонат кальция) для повышения pH, однако его избыток приведет к потере азота.
Исследование городских компостных куч в Беркли (Калифорния) показало, что при увлажнении и аэрировании больших куч образование компоста может быть завершено уже за 2 недели. Обычно на завершение процесса уходит три или более зимних месяцев. Если компост внести в почву прежде, чем процесс его образования закончен, это может временно обеднить почву растворимым азотом.
Из-за резкого сокращения объема растительных остатков при компостировании оно может быть очень удобным способом уничтожения отходов. В Скарсдейле (шт. Нью- Йорк), например, листья, компостированные на городской свалке, уменьшились до 1/5 своего первоначального объема. К тому же из них образовалась добавка к почве, улучшающая ее аэрацию и водоудерживающую способность. Химический анализ показывает, что богатый компост содержит в пересчете на сухую массу только около 1,5 — 3,5% азота, 0,5 — 1,0% фосфора и 1,0 — 2,0% калия, что значительно меньше, чем в искусственных удобрениях. Однако в отличие от этих удобрений компост может служить источником почти всех известных элементов, необходимых растениям. Компост обеспечивает длительное и сбалансированное поступление питательных веществ, которые постепенно образуются по мерс его разложения в почве.
В то время как минеральные удобрения становятся все дороже и труднодоступнее, а водоемы все сильнее загрязняются ими и органическими остатками, компостирование становится все более привлекательной альтернативой.
Каждая половина листа венериной мухоловки снабжена тремя чувствительными волосками. Если насекомое попадает на один из листьев, привлеченное нектаром, выделяемым на его поверхность, оно задевает волоски и запускает таким образом механизм закрывания листа, действующий подобно ловушке. Зубчатые края смыкаются, половинки листа постепенно сжимаются, и насекомое оказывается прижатым к переваривающим железкам, расположенным на поверхности ловушки (рис. 25-22).
Рис. 25-23. А. Листья люпина (Lupinus arizonicus), поворачивающиеся вслед за солнцем. Это явление называют гелиотропизмом. Б. Гелиотропизм у подсолнечника
