БОТАНІКА. МОРФОЛОГІЯ РОСЛИН - О. А. Шевчук - 2014
ЧЕРГУВАННЯ ПОКОЛІНЬ ТА ЦИКЛИ ВІДТВОРЕННЯ У РОСЛИН
Чергування поколінь
Для всіх організмів, які розмножуються статевим шляхом, характерне чергування ядерних фаз — диплоїдної та гаплоїдної. При статевому процесі відбувається подвоєння числа хромосом у ядрі зиготи. Щоб запобігти прогресуючому подвоєнню числа хромосом у ряді наступних поколінь, у результаті тривалої еволюції виробився протилежний процес — мейоз (редукційний поділ), внаслідок якого утворюються клітини з одинарним (гаплоїдним) набором хромосом в ядрі. Отже, запліднення дає початок диплоїдній фазі, мейоз — гаплоїдній. Ці два процеси тісно пов'язані між собою і фактично є частинами одного життєвого циклу. Організм, що утворився у результаті запліднення з диплоїдної зиготи (це диплоїдна ядерна фаза, або диплофаза), називається диплобіонтом. Організм, що утворився після мейозу і подальшого мітозу (це гаплоїдна ядерна фаза, або гаплофаза), називається гаплобіонтом.
На відміну від вищих тварин і людини, в статевих клітинах яких мейоз відбувається безпосередньо перед утворенням гамет, у різних груп рослин мейоз відбувається в різні періоди їхнього життя, а також існує багато варіантів зміни ядерних фаз.
У бурої водорості фукуса, наприклад, кожна особина є диплобіонтом, тобто всі її клітини мають подвійний набір хромосом. У статевозрілих особин утворюються оогонії та антеридії, в яких у результаті мейозу виникають гамети — гаплоїдні яйцеклітини та сперматозоїди. Після запліднення у воді диплоїдні зиготи покриваються оболонкою і дають початок новим диплобіонтам.
У багатьох водоростей (зелена водорость улотрикс) кожна особа є гаплобіонтом, тобто всі її клітини, на відміну від фукуса, мають одинарний (не парний) набір хромосом. Спеціалізованих гаметангіїв в улотрикса немає, тому в кожній клітині нитчастого тіла статевозрілих особин у результаті мітозу можуть утворюватись ізогамети, що попарно зливаються у воді. Диплоїдна зигота покривається оболонкою і після періоду спокою мейотично ділиться, утворюючи чотири мейоспори. Отже, з диплоїдної зиготи утворюється не одна диплоїдна особина, як це відбувається у фукуса, а чотири гаплоїдних. Крім мейоспор, в клітинах тіла улотрикса можуть утворюватись і мітоспори. В улотрикса диплофаза представлена лише зиготою, тобто і в нього змінюються ядерні фази, але інакше, ніж у фукуса. Перехід від галоїдної фази до диплофази відбувається при заплідненні, а від диплофази до гаплофази — при утворенні мейоспор, а не гамет.
Таким чином, в обох варіантах зміни ядерних фаз є або диплобіонт (всі особини фукуса), або гаплобіонт (всі особини улотрикса). В першому варіанті відсутній гаплобіонт, а є лише гаплофаза у вигляді гамет; у другому — відсутній диплобіонт, а є лише диплофаза у вигляді зиготи.
У багатьох рослин в життєвому циклі може утворюватись і диплобіонт, і гаплобіонт, які представлені різними поколіннями, що чергуються. Чергуванням поколінь називають закономірну зміну в життєвому циклі генерацій (поколінь, або біонтів), котрі відрізняються способом розмноження. Диплобіонт — це нестатеве (спорове) покоління, спорофіт, який розвивається із зиготи в результаті злиття двох гамет і продукує спори. Гаплобіонт — це статеве покоління, гаметофіт, який розвивається із мегаспори і продукує гамети. Органи нестатевого розмноження (спорангії, зооспорангії) розвиваються на спорофіті; в результаті мейозу в них утворюються гаплоїдні спори, котрі проростають у нові статеві покоління. Статеві органи утворюються на гаметофіті, причому він може бути одностатевим (політрихум, маршанція, сальвінія) або двостатевим (плаун, орляк, чоловіча папороть). Гаметофіт та спорофіт можуть бути однакові морфологічно і за віком (ізоморфне чергування поколінь) або різко відрізнятися (гетероморфне чергування поколінь).
У водоростей зустрічаються обидві форми чергування поколінь. Ізоморфна зміна поколінь притаманна більшості червоних, деяким бурим та деяким зеленим водоростям. У гаплоїдній фазі такі водорості розмножуються статевим шляхом, тобто утворюють гамети, котрі при заплідненні зливаються в зиготу. При проростанні зигота дає нову водорість з диплоїдним числом хромосом, яка може бути цілком схожа на гаплобіонт або більш-менш (у деяких навіть дуже різко) різнитися від нього. Ця диплоїдна форма розмножується нестатевим шляхом, спорами, при утворенні яких відбувається редукційний поділ. Зі спор розвивається гаплоїдна водорость, котра продукує гамети; зигота знову дає диплоїдну особину, яка розмножується нестатевим шляхом тощо. Таким чином, тут має місце не лише зміна гаплоїдної та диплоїдної фаз, а й відповідне чергування статевого і нестатевого поколінь, котрі розмножуються статевим і споровим способами. Така зміна поколінь притаманна батрахоспермуму з червоних водоростей, ектокарпусу з бурих, кладофорі скупченій із зелених.
При гетероморфній зміні поколінь обидва покоління розвиваються або незалежно одне від одного (ламінарія, рівноспорові папороті, плауни, хвощі), або одне з поколінь, яке не може самостійно розвиватися, існує за рахунок іншого (мохи і всі насінні рослини). При цьому в циклі розвитку переважає лише одне з поколінь (гаметофіт або спорофіт). У вищих рослин до гаметофітної лінії еволюції відносяться лише мохоподібні, в яких спорофіт (спорогон) розвивається на гаметофіті, тобто на зеленій рослині, а до спорофітної — решта вищих рослин. Спорофіт (він же диплобіонт) - це листкостеблова рослина, на якій розвиваються спорангії. Гаметофіт (заросток) розвинутий слабше, недовговічний (за винятком плаунів) і представлений двостатевим або одностатевим таломом, який живе самостійно (папороті, плауни, хвощі). В голонасінних і квіткових рослин гаметофіти являють собою дрібні мікроскопічні утвори, котрі розвиваються частково або повністю на спорофіті, причому за його рахунок.
Цикли відтворення
Поняття про цикли відтворення. В часі життя кожного виду відбувається у формі безперервної зміни послідовних поколінь особин, обумовленої здатністю кожної особини давати потомство. Потомство виникає різними шляхами.
Статевий процес та мейоз закономірно чергуються, спостерігається правильна зміна ядерних фаз, процесів утворення спорангіїв, гаметангіїв і гамет, сингамія, зміна морфологічних та фізіологічних станів особини. Інколи все це повторюється в кожному поколінні. Якщо особини всіх поколінь однакові (лише диплобіонти чи лише гаплобіонти), відтворення здійснюється без зміни поколінь. У папороті, плауна, хвоща чергуються два різних покоління, між якими розподілені функції утворення гамет і мейоспор. Дочірні особини при цьому не подібні до батьківських: поява їх не є відтворенням, воно здійснюється лише через одне покоління. Спорофіт утворює лише гаметофіти (зі спор), гаметофіт утворює лише спорофіт (через статевий процес).
Отже, в житті виду (послідовній зміні поколінь особин) чітко виділяються цикли відтворення, які часто називають циклами розвитку. Цикл відтворення — це відрізок життя виду, обмежений двома однойменними етапами: від гаплобіонта до гаплобіонта, від спорофіта до спорофіта, від зиготи до зиготи, від мейоспори до мейоспори. В ході циклу відтворення обов'язково відбувається зміна ядерних фаз.
У природі існує багато циклів відтворення, особливо вони різноманітні в нижчих рослин. Подібні між собою цикли відтворення у хвощів, плаунів, рівноспорових папоротей. Дещо відмінне, але явне чергування поколінь мають різноспорові папоротеподібні (сальвінії) та різноспорові плауноподібні (плаунки). І зовсім специфічним чергуванням поколінь, навіть прихованим, а не явним, характеризуються насінні рослини, в яких гаметофіти дуже редуковані.
Мохоподібні. Розглянемо на прикладі рунянки звичайної чергування поколінь у мохоподібних.

Рис. 7. Зозулин льон (Polytrichum commune): А — жіноча рослина зі спорогоном (з ковпачком і без нього); Б — чоловіча рослина з розеткою на верхівці; В — поперечний зріз листка з асиміляторами; Г — верхівка чоловічої рослини з антеридіями і парафізами; Д — антеридій: ніжка, оболонка і спермагенна тканина; Е — форми парафіз; Ж — верхівка жіночої рослини: архегонії оточені листками; З — архегоній: у його шийці канальцеві клітини, в черевці — яйцеклітина, над нею черевцева канальцева клітина; І — протонема моху з ризоїдами і бруньками; К — поздовжній зріз спорогона: всередині спорангій, що оточує колонку, нагорі кришечка; Л — перистом з епіфрагмою.
На верхівках стебла цієї рослини та багатьох інших мохів утворюються чоловічі (антеридії) та жіночі (архегонії) органи статевого розмноження. Вони оточені листками і розміщені на різних особинах. Антеридій має вигляд продовгуватого або округлого мішечка з одношаровою стінкою, що оточує дрібні сперматогенні клітини. Із кожної сперматогенної клітини утворюються один або два спірально зігнутих сперматозоїди (сперматозоони) з двома довгими джгутиками на передньому кінці.
Архегоній завжди багатоклітинний і схожий на колбу з довгим горлом. У нижній здутій частині архегонія (в черевці) розташовується не вкрита оболонкою жіноча статева клітина (яйцеклітина), а над нею — маленька черевцева канальцева клітина. У вузькій верхній частині архегонія (шийці) утворюється канал, який містить дрібні канальцеві клітини. Ці клітини розпливаються в слиз, який заповнює канал шийки і частково виходить назовні крізь розкриту на верхівці шийку. Одночасно на верхівці розкриваються антеридії, і звільняються численні сперматозоїди. Рухаючись у воді (без якої запліднення неможливе), сперматозоїди за допомогою джгутиків направляються до архегоніїв, оскільки їх приваблюють речовини, що містяться в слизі каналу шийки архегоніїв. Один із сперматозоїдів проникає в шийку архегонія, направляється до яйцеклітини і зливається з нею. Запліднена яйцеклітина покривається оболонкою, одразу починає ділитись і утворює спорофіт, який у мохоподібних називають спорогоном. Він дуже редукований і являє собою коробочку на ніжці, яка закінчується гаусторієм у стеблі моху. Всі клітини спорогонія утворились із зиготи і мають подвійний набір хромосом. У коробочці з клітин археспорія після редукційного поділу їхніх клітинних ядер утворюються мікроскопічне дрібні одноклітинні гаплоїдні спори, котрі служать для нестатевого розмноження , бо виникли нестатевим шляхом.
У вищих спорових рослин утворенню спор передує редукційний поділ, і зі спор розвивається інше покоління (гаметофіт) з іншим числом хромосом, відмінне від рослини, яка утворила спори.
Після дозрівання спор у коробочці з неї опадає кришечка, спори висіваються, розносяться вітром і, потрапивши в сприятливі умови, проростають. Із проростаючої спори утворюється протонема— дихотомічно розгалужена нитка, на якій формуються бруньки. З них розвиваються листкостеблові різностатеві рослини моху, які далі утворюють на верхівках статеві органи — антеридії та архегонії. Клітини протонеми і утворені на них типові рослини моху із статевими органами мають ядра з гаплоїдним числом хромосом. Диплоїдна фаза починається із запліднення і закінчується під час поділу клітин археспорія перед утворенням спор. Отже, коробочка на ніжці з гаусторієм (спорогон) є диплоїдом.
Таким чином, у циклі розвитку мохів відбувається правильне чергування статевого і нестатевого способів розмноження. Органи статевого розмноження утворюються на листкостебловій або таломній рослині, яка разом з протонемою є статевим поколінням, або гаметофітом, адже його клітини мають ядра з гаплоїдним числом хромосом. Клітини, які служать для нестатевого розмноження (спори), утворюються в спорогоні, котрий є нестатевим поколінням, або спорофітом; його клітини, за винятком спор, мають ядра з диплоїдним числом хромосом. Спорофіт, хоча й відмежований морфологічно й анатомічно від гаметофіта, ніколи не відокремлюється від нього і живиться або повністю за рахунок останнього, або, маючи хлорофіл, утворює органічні речовини і бере з гаметофіта воду та неорганічні солі. Після розсіювання спор спорофіт відмирає.
Отже, в циклі розвитку мохів чергуються два покоління, причому домінує статеве покоління (гаметофіт), а нестатеве (спорофіт) — має менший вік і повністю залежить від гаметофіта.
Рівноспорові плауни. Розглянемо на прикладі плауна булавовидного чергування поколінь у плаунів. На верхівках вертикальних пагонів рослин утворюються спороносні колоски (стробіли), до осі яких прикріплюються спорофіли з нирковидними спорангіями. Зі спорогенної тканини в спорангіях шляхом мейозу утворюються однакові мейоспори. Після дозрівання їх спорангії розкриваються поперечною щілиною, спори висипаються, підхоплюються вітром і розносяться.
За сприятливих умов спори дуже повільно проростають у безбарвний підземний заросток (гаметофіт), що веде сапрофітний спосіб життя. У плауна булавовидного гаметофіт двостатевий, на ньому утворюються архегонії та антеридії типової будови. Запліднення можливе лише при наявності води, в якій рухаються дводжгутикові сперматозоїди від антеридія до шийки архегонія.
Після злиття гамет, утворених шляхом мітозу, виникає диплоїдна зигота, з якої бере початок диплобіонт, тобто нестатеве покоління (спорофіт). Спершу утворюється зародок, котрий живиться за рахунок гаметофіта, а далі, з появою листків і кореня, він переходить до самостійного життя. Отже, в плауна чергуються два незалежних самостійних покоління, які виконують різні функції.
Спорофіт — це багаторічна рослина з добре розвинутими вегетативними органами, пристосована до умов наземного життя, а гаметофіт — маленька дзигоподібна рослина, яка здійснює статевий процес. Зміну поколінь плауна булавовидного називають гетероморфною і антитетичною. Спорофіт тут утворює мільйони спор, які забезпечують розмноження виду.
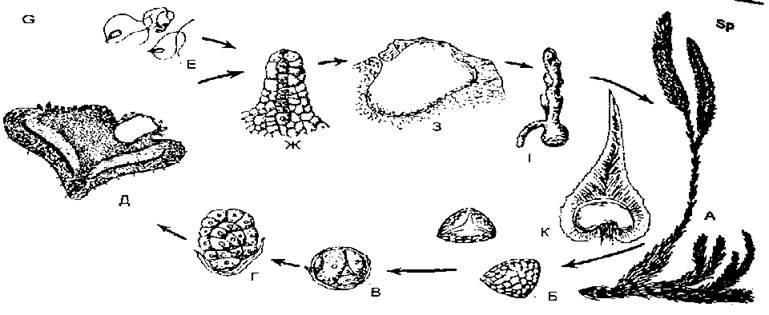
Рис. 8. Цикл розвитку плауна булавовидного (Lycopodium davatum): А — загальний вигляд рослини (спорофіт); Б - Г — розвиток заростка із спори; Д — заросток (гаметофіт); Е — сперматозоїди; Ж — архегоній: 3 — зародок; І — молода рослина; К — спорофіл із спорангієм.
Хвощеподібні. Розглянемо на прикладі хвоща польового чергування поколінь хвощеподібних. На верхівках спороносних пагонів цієї рослини утворюються стробіли, котрі складаються з особливих шестикутних спороносних щитків (спорангієфорів), які прикріплені центральною ніжкою до стробіла. На внутрішньому боці щитка, повернутому до стебла, розташовуються 8-15 спорангіїв, які мають вигляд мішків. У археспоріальній тканині спорангіїв відбувається редукційний поділ клітинних ядер і утворюються численні однакові кулясті зелені спори, котрі служать для розмноження та розселення рослин. Кожна спора має тришарову оболонку. Зовнішній шар її розривається на дві спірально закручені гігроскопічні стрічки (елатери), котрі у вологому середовищі щільно прилягають до спор, а при висиханні розкручуються. Такі стрічки скріплюють спори в нещільні клубочки, які розносяться вітром або водою при висіванні із спорангіїв. За сприятливих умов спори проростають, формуючи заростки (гаметофіт). Заростки мають вигляд маленьких зелених пластинок, розсічених на стрічковидні лопаті. На кінцях лопатей розвиваються антеридії, а в них багатоджгутикові сперматозоїди. Колбовидні архегонії розвиваються в основі стрічковидних лопатей гаметофіта і черевцем занурені в його тканину. Рухаючись в краплях води, сперматозоїди проникають крізь шийку архегонію до яйцеклітини, і один з них зливається з нею. Після запліднення із зиготи розвивається нове нестатеве покоління, що представляє власне хвощ.
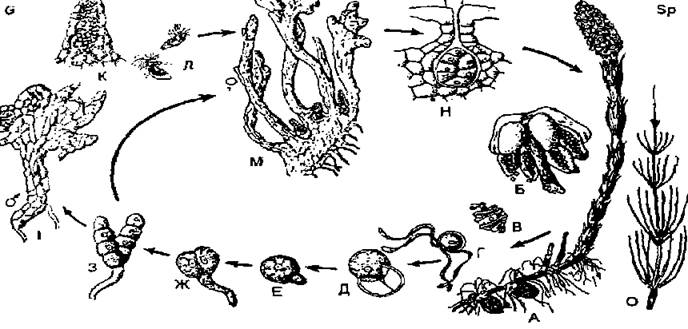
Рис. 9. Цикл розвитку хвоща польового (Eqrnsetum arvense): А — спороносний пагін; Б — спорофіл із спорангіями; В - 3 — розвиток заростка (гаметофіта) зі спори; І — заросток з антеридіями; К — антеридій; Л — сперматозоїди; М — заросток з архегоніями; Н — архегоній; О — асимілюючий пагін.
Отже, власне хвощ — це диплобіонт, спорофіт, нестатеве покоління. Гаплобіонт — це заростки хвоща, гаметофіт, статеве покоління, незалежне у своєму живленні від диплобіонта. Г аметофіти можуть бути одностатеві (чоловічі та жіночі) і двостатеві (тобто такі, що несуть одночасно архегонії і антеридії).
Таким чином, у циклі розвитку хвощів відбувається таке ж, як і в плаунів, чергування поколінь; диплобіонта (спорофіта) і гаплобіонта (гаметофіта), так само за ступенем морфологічного розвитку і за тривалістю життя домінує спорофіт. Як і в плаунів, тут відбувається гетероморфна й антитетична зміна поколінь. Кожна зигота так само виростає у великий спорофіт, який утворює багато мейоспор.
Рівноспорові папороті. Розглянемо на прикладі щитника чоловічого, або чоловічої папороті чергування поколінь у рівноспорових папоротей. На листках цієї рослини з нижнього боку утворюються соруси (зібрання спорангіїв). Спороносні листки, на яких формуються спорангії, називають спорофілами.
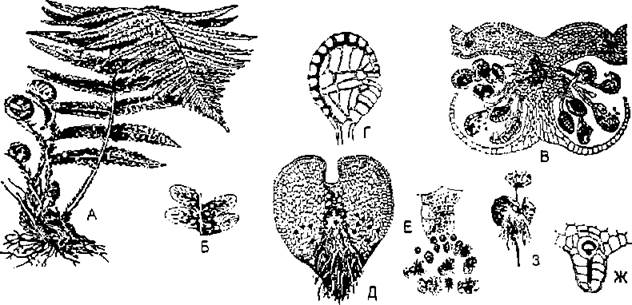
Рис. 10. Щитник чоловічий (Dryopteris filix-mas): А — загальний вигляд; Б — частинки листка з сорусами на нижньому боці; В — розріз листка та соруса: плацента, індузій, спорангії; Г — спорангій з кільцем; Д — заросток (гаметофіт); Е — антеридії в момент виходу сперматозоїдів; Ж—архегоній з яйцеклітиною; З — молодий спорофіт на гаметофіті.
Кожен спорангій закладається на нижньому боці листка як меристематичний горбик. Зовнішні його клітини утворюють стінку спорангія, а внутрішня клітина при поділі — археспоріальну тканину. Із частини її клітин утворюється вистилаючий шар, або тапетум, а з решти після кількох мітотичних поділів формується спорогенна тканина, клітини якої є материнськими клітинами спор. Вони діляться шляхом мейозу і дають тетради гаплоїдних мейоспор. Клітини тапетума використовуються для живлення спор і формування їхньої зовнішньої оболонки. Дозрілі спори покриті подвійною оболонкою. Зовнішня (екзина) — товста, горбкувата, добре захищає вміст спори від висихання. Після розкриття спорангія спори висипаються і розносяться вітром.
Таким чином, забезпечується розмноження і розселення папороті. Це нестатеве покоління щитника чоловічого. За сприятливих умов спори проростають і в результаті мітотичного поділу клітин утворюють заросток. Він є гаплобіонтом, який формою та розмірами дуже відрізняється від папороті — диплобіонта. Заросток являє собою зелену багатоклітинну пластинку серцевидної форми завдовжки до 1 см, не розчленований на стебло та листки, не має коренів і схожий на пластинчасті водорості. Ця пластинка дуже чутлива до вологи; на її нижньому боці є ризоїди — однорядні нитки, якими вона прикріплюється до ґрунту. Всі клітини пластинки зелені, хлорофілоносні, епідерма не виражена. На нижньому боці заростка виникають гаметангії, причому спершу антеридії, а пізніше архегонії, в яких мітотичним шляхом утворюються гамети — сперматозоїди та яйцеклітини. Отже, заросток виконує статеву функцію: формує гамети, а тому є гаметофітом, або статевим поколінням. Антеридії папороті мають одношарову стінку і заповнені сперматогенними клітинами, які дають багато-джгутикові сперматозоїди штопороподібної форми. При наявності води між нижнім боком заростка і грунтом антеридії відкриваються, сперматозоїди виходять у воду і підпливають до архегоніїв. Архегонії колбовидної форми з розширеним черевцем, яким вони занурюються в тканину заростка, та вузькою шийкою. В черевці формується яйцеклітина і черевцева канальцева клітина, в шийці — кілька шийкових канальцевих клітин. При дозріванні архегонія шийка відкривається, всі канальцеві клітини ослизнюються, слиз виходить у воду. Речовини, які утворилися в слизу, приваблюють сперматозоїди, і один із них запліднює яйцеклітину. Зигота в архегонії ділиться шляхом мітозу і утворює зародок нового спорофіта. Спочатку він розвивається за рахунок заростка, а пізніше, коли утворюються листки і додатковий корінь, переходить до самостійного живлення. Заросток, виконавши свою функцію, гине.
Отже, в папороті закономірно змінюються не лише ядерні фази, межами яких є утворення мейоспор (перехід від диплофази до гаплофази) і статевий процес (перехід від гаплофази до диплофази), а й два різних за габітусом і функціями самостійних покоління рослин: нестатеве
(диплоїдний спорофіт) і статеве (гаплоїдний гаметофіт). Відтворення собі подібних відбувається через одне покоління.
Спорофіт папороті — це багаторічна рослина з добре розвинутими органами, яка пристосувалась до умов наземного життя; гаметофіт — маленька недовговічна рослина зі слабко диференційованим таломом, пристосована здійснювати статевий процес у приземному шарі води, без якої не відбувається запліднення. Спорофіт дає потомство через мейоспори, гаметофіт — через гамети. Таку зміну поколінь називають гетероморфною і антитетичною. На відміну від улотрикса, де зигота дає лише чотири мейоспори, а не організм (диплобіонт), у папороті кожна зигота виростає в спорофіт, який утворює багато мейоспор.
Різноспорові плауни. Серед вищих спорових є такі рослини, в яких утворюються два типи спор і відповідно два типи заростків. Розглянемо цикл відтворення селагінели (плаунка). Листкостеблова рослина плаунка — це спорофіт, диплобіонт. На кінцях пагонів у плаунка формуються спороносні колоски, або стробіли, утворені віссю і сидячими на ній лусковидними спорофілами. Спорангії прикріплені короткою ніжкою до основи верхнього боку листка — спорофіла. Спорангії неоднакові: в одному стробілі є мікроспорангії з мікроспорами та мегаспорангії з мегаспорами. В спорангіях є стінка, вистилаючий шар і спорогенна тканина. В мікроспорангіях всі спорогенні клітини діляться шляхом мейозу і утворюють дуже багато мікроспор. У мегаспорангіях материнською клітиною мегаспор стає лише одна клітина спорогенної тканини, яка розростається і займає майже всю порожнину спорангія, а інші клітини відмирають і використовуються на її живлення. Материнська клітина ділиться шляхом мейозу і утворює тетраду мегаспор, покритих товстою горбкуватою чорною оболонкою. Ці спори добре помітні навіть неозброєним оком. При дозріванні спори обох типів висипаються зі спорангіїв або ж, як у деяких видів плаунків, мегаспори залишаються в мегаспорангіях. За сприятливих умов спори проростають і утворюють заростки різних типів. З мікроспор утворюються дуже маленькі редуковані чоловічі заростки, з мегаспор — дещо більшого розміру, жіночі. Мікро-або мегаспори залишаються в оболонці. В деяких видів плаунків проростання спор починається всередині спорангіїв.
Чоловічий заросток (гаметофіт) утворений однією вегетативною клітиною (її називають ризоїдальною) і одним антеридієм, в якому формуються дводжгутикові сперматозоїди. За наявності води антеридій розкривається, сперматозоїди виходять з нього і підпливають до жіночих заростків. Після виходу сперматозоїдів чоловічий гаметофіт, відігравши свою роль в статевому процесі, гине.
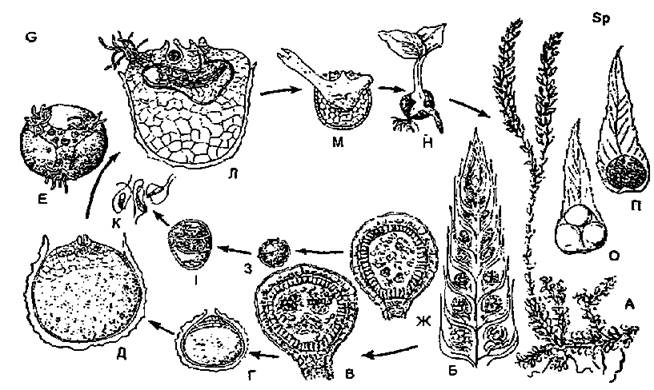
Рис. 11. Цикл розвитку селагінели (8еіа§іпеііа Бр.): А — загальний вигляд рослини (спорофіт); Б — стробіл; В — мегаспорангій; Г - Е — розвиток жіночого заростка з мегаспори; Ж — мікроспорангій: З, І — розвиток мікроспори; К — сперматозоїди; Л — жіночий заросток із зародком; М - Н — проростаюча рослина; О — спорофіл з мегаспорангієм; П — спорофіл з мікроспорангієм.
Дозрілий жіночий заросток (гаметофіт) утворений багатьма клітинами в оболонці мегаспори і частково висувається назовні крізь трипроменеву тріщину в оболонці. Тут утворюються пучки ризоїдів і формується кілька архегоніїв, занурених у тканину заростка. Після проникнення сперматозоїдів в архегоній і запліднення яйцеклітини зигота в одному з архегоніїв починає розвиватися в заросток нового спорофіта, який незабаром розвиває зелений пагін і додатковий корінь. Жіночий гаметофіт після цього гине.
Біологічне значення різноспоровості полягає в тому, що чоловічий та жіночий гаметофіти і молодий заросток спорофіта краще захищені від висихання порівняно з гаметофітами чоловічої папороті, хвощів, плаунів (у них гаметофіти залишаються в оболонках мікро- та мегаспори). Утворення гаметофітів усередині оболонки спор супроводжується зменшенням розмірів гаметофітів, їх редукцією, особливо в чоловічого заростка, який потрібний лише для утворення сперматозоїдів. Малі розміри гаметофітів компенсуються великою кількістю численних мікроспор. Різностатевість заростків забезпечує перехресне запліднення, а рання фіксація різностатевості виражається в різноспоровості.
Отже, на прикладі селагінели можна простежити важливу тенденцію в еволюції зелених наземних рослин: сховати і краще захистити вологолюбні гаметофіти від небезпеки наземного життя. Однак, у різноспорових рослин (селагінел) спорофіт і гаметофіт представлені самостійними особинами, які живуть на субстраті, хоча й з редукованою здатністю до самостійного живлення в гаметофіта, особливо чоловічого. Статевий процес залишається повністю залежним від наявності води. Функцію розмноження і розселення виконують спори, причому мегаспори відіграють основну роль, бо забезпечують захист і живлення заростка і зародка.
Різноспорові папороті. До різноспорових рослин відноситься також водяна папороть — сальвінія плаваюча, яка зрідка зустрічається на поверхні водойм. Ця маленька (завдовжки 5-15 см) рослина занесена до Червоної книги України. Біля основи підводних листків у неї утворюються зібрання дрібних мікроспорангіїв і більших мегаспорангіїв. На одній рослині утворюються окремі соруси з мікро- та мегаспорангіями. Соруси оточені з усіх боків індузієм — оболонкою, котра захищає спорангії від загнивання у воді. В мікроспорангіях зі спорогенної тканини, шляхом мейозу, утворюється багато мікроспор. В мегаспорангіях з однієї материнської клітини мегаспор утворюється чотири мегаспори, однак, розвивається лише одна, а три — рано відмирають. Мікро- і мегаспори, не висипаючись зі спорангіїв, проростають і після відмирання рослини розносяться водою. З мікроспор весною розвиваються чоловічі заростки (гаметофіти), які проривають стінку мікроспорангія і висуваються назовні. Чоловічий заросток (гаплобіонт) безбарвний, коротенький (до 0,5 мм). Вегетативна частина його має дві клітини. Крім них, у верхній частині є два антеридії, які мають стінки з двох клітин. У кожному антеридії є чотири злегка заокруглених, багатоджгутикових сперматозоїди. Мегаспора проростає в жіночий заросток (гаплобіонт), який має вигляд трикутної зеленої пластинки завбільшки з булавочну голівку, яка плаває на поверхні води. В тканину гаметофіта занурені кілька архегоніїв з редукованою короткою шийкою. Після запліднення яйцеклітини із зиготи розвивається нова рослина сальвінії — спорофіт (диплобіонт), представлений спочатку зародком у стінці архегонія і захищений стінкою мегаспори. Отже, в сальвінії, як і в плаунка, спорангії та спори різняться за розміром. Спори проростають усередині спорангіїв. У сальвінії спостерігається редукція одностатевих гаметофітів і нездатність чоловічого безбарвного гаметофіта до самостійного життя: він живе за рахунок запасів поживних речовин, що були в мікроспорі.
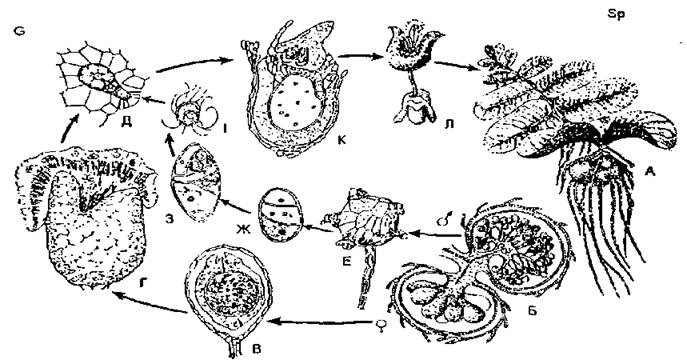
Рис. 12. Цикл розвитку сальвінії плаваючої (Salvinia natans): А — загальний вигляд (спорофіт); Б — мега- і мікроспорангієсоруси; В, Г— розвиток жіночого гаметофіта; Д — архегоній; Е-З — розвиток чоловічого гаметофіта; І — сперматозоїд; К — зародок; Л — молода рослина.
У сальвінії відбувається чітке чергування поколінь — диплоїдного (спорофіта) та гаплоїдного (гаметофіта), хоча й має місце поступова редукція гаметофітів.
Різноспоровість — прогресивне явище в еволюції рослин, яке спричинило в кінцевому результаті появу насінних рослин. Різноспоровість виникала в процесі еволюції неодноразово, і редукція гаметофітів здійснювалась у різних груп рослин самостійно.
Голонасінні. У ході подальшої еволюції виникла група рослин, які розмножуються не спорами, а насінням. Так з'явилися голонасінні та покритонасінні. Розглянемо, як відбувається розмноження та чергування поколінь у голонасінних.
Виникнення різноспоровості було важливим етапом еволюцїі рослинного світу. З'явилося два типи спор, два типи гаметофітів і одночасно відбувалася редукція останніх, які поступово втрачали здатність до самостійного способу живлення і все більше починали залежати від спорофіта. При вивченні голонасінних дуже важливо чітко усвідомити, що мегаспори, які після редукції трьох лише по одній утворюються в мегаспорангіях, взагалі не висипаються з них, а залишаються на материнській рослині і проростають, даючи гаметофіт всередині спорангія. Тут же відбувається сингамія, незалежно від того, є вода чи ні.
Отже, головною відміною насінних рослин від вищих спорових є те, що гаметофіти (особливо жіночий) втратили здатність до самостійного існування і живуть тільки за рахунок спорофіта. При цьому виникло запилення — надзвичайно важливе пристосування до наземного життя, яке дало величезні переваги насінним рослинам перед споровими. Насінним рослинам уже не була потрібна вода для руху гамет, тобто стався повний відрив від водного середовища при заплідненні, а чоловічі гаметофіти стали переноситися рухом повітря.
Насінні рослини різноспорові. На рослині-спорофіті в них, як і в спорових, утворюються мікро- та мегаспорангії з мейоспорами, котрі, на відміну від спорових, проростають тут же під покривами спорангіїв на материнських рослинах, не висипаючись назовні. Отже, розвиток і чоловічих, і жіночих гаметофітів відбувається всередині мікро- і мегаспорангіїв.
У результаті статевого процесу з'являється справжній шедевр еволюції — насінина, яка служить для відтворення, розмноження та розселення насінних рослин. Насіння утворюється у великій кількості на материнській рослині, потім опадає з неї, даючи початок новим рослинам, подібним до материнської. Чергування поколінь при цьому замасковане, але при детальному вивченні процесів формування насіння досить добре розпізнається. Насіння має ряд переваг перед спорами. В ньому є більший чи менший запас поживних речовин, необхідний для розвитку зародка та при проростанні насіння. Важливим також є більший чи менший період спокою насіння і добра захищеність його шкіркою, що дозволяє переносити несприятливі зовнішні умови.
Саговникові. Однією з найбільш примітивних груп голонасінних є саговники, що живуть в тропіках та субтропіках. Це дводомні рослини (диплобіонти), які утворюють на одних екземплярах зібрання мікроспорофілів — мікростробілів (їх називають не зовсім точно чоловічими шишками). В представників роду саговник типові мегастробіли не утворюються. На мегаспорофілах саговників утворюється від двох до восьми видозмінених мегаспорангіїв, які в голонасінних називаються насінними зачатками. Всередині насінного зачатка є мегаспорангій (нуцелюс) — центральна багатоклітинна частина, оточена зовні покривом, який виростає з основи нуцелюса, так званої халази. Покрив або інтегумент, обгортає нуцелюс поступово, знизу вгору, не змикаючись на вершині і залишаючи отвір для входження пилку — пилковхід, або мікропіле. Палеоботанічні знахідки підтвердили гіпотезу про синангіальне походження інтегумента. В предків сучасних голонасінних по кілька мегаспорангіїв зростались, утворюючи синангій. В подальшому всі периферійні мегаспорангії стали безплідними, редукувались і утворили захисний покрив (інтегумент) навколо одного центрального мегаспорангія. Так утворився насінний зачаток.
В нуцелюсі (мегаспорангії) утворюється одна мегаспора. Одна з клітин спорогенної тканини ділиться двічі з редукцією числа хромосом, формуючи тетраду гаплоїдних мегаспор. Одна з них, розростаючись, витісняє інші. Вона проростає в насінному зачатку, даючи безбарвний багатоклітинний заросток — первинний ендосперм. Лише на верхівці заросток трохи висувається з оболонки мегаспори, і в ньому утворюються два (інколи більше) архегонії завдовжки до 4 мм. Кожен з них має велику яйцеклітину, черевцеву канальцеву клітину і коротку шийку. Частина клітин нуцелюса над жіночим заростком розчиняється, з’являється заглиблення — пилкова камера, наповнена слизистою цукристою рідиною, що виступає з камери назовні. Весь заросток (ендосперм) разом із архегоніями утворює жіночий гаметофіт.
Зібрані в мікростробіли мікроспорофіли на нижньому боці несуть в сорусах купки мікроспорангіїв, в яких шляхом мейозу з клітин спорогенної тканини розвиваються мікроспори. Мікроспорангії при підсиханні розкриваються завдяки нерівномірним потовщенням на оболонках клітин зовнішнього шару їхньої стінки. Мікроспори ще в мікроспорангіях починають проростати, утворюючи три клітини. Одна маленька стає вегетативною клітиною чоловічого заростка; із середньої в подальшому в пилковій камері утворюється два великих сперматозоїди з численними джгутиками, розміщеними по спіралі; з третьої клітини мікроспори розвивається гаусторій, який заглиблюється у тканину нуцелюса і служить для прикріплення пилинки та її живлення за рахунок останнього. Його можна гомологізувати з вегетативними клітинами біля типових чоловічих гаметофітів. Пророслі мікроспори називають пилинками, або пилковими зернами. Вони розносяться вітром, і деякі потрапляють у рідину, що виступає з пилкової камери насінного зачатка. При підсиханні рідини пилинки втягуються в пилкову камеру, де лопається їхня оболонка (екзина). Сперматозоїди плавають у рідині і направляються до архегоніїв. Один із них проникає в яйцеклітину і зливається з нею. Зигота починає ділитись, утворює зародок, який на підвіску вростає в первинний ендосперм (заросток) і розвивається за рахунок його запасних поживних речовин. У зародку формуються дві сім'ядолі, між ними — точка росту стебла, під ними — первинне стебло (підсім'ядольне коліно), яке переходить у корінь. Навколо зародка в заростку формується тканина — ендосперм, що в подальшому використовується ним при проростанні насіння. З залишку нуцелюса та з покриву (інтегумента) насінного зачатка формується насінна шкірка. Отже, мегаспорангій у саговників перетворюється в насіння з одним зародком. У багатьох саговників насінні зачатки опадають після запилення, ще до запліднення. Злиття гамет може відбуватись уже в опалих насінних зачатках — на поверхні ґрунту. Тут же формується зародок та насінина.
Отже, у саговників, які розмножуються насінням, а не спорами, при детальному аналізі теж можна констатувати чергування двох поколінь— нестатевого (спорофіта, диплобіонта) та статевого (гаметофіта, гаплобіонта). Однак, спорофіт (багаторічна рослина з добре розвинутими вегетативними органами) значно домінує над редукованим різностатевим гаметофітом. Чоловічий гаметофіт — мікроскопічна пилинка, що служить лише для перенесення та утворення чоловічих гамет, редукований настільки, що навіть не утворює антеридія. Жіночий гаметофіт — ендосперм з двома архегоніями — не покидає материнської рослини, розвивається всередині мегаспорангія і теж значно редукований, не здатний самостійно існувати.
Хвойні. Хвойні — найбільша група голонасінних, представлена багатьма видами дерев та чагарників у помірному кліматі.
Розглянемо особливості відтворення, формування та розсіювання насіння хвойних на прикладі сосни звичайної. Сосна — спорофіт, на якому утворюються органи спороношення: мікро- та мегастробіли, які умовно називають чоловічими та жіночими шишками. Це зібрання мікро- та мегаспорофілів.
Жіноча шишка складається з укороченої осі та плівчастих покривних лусок. В їх пазухах розвиваються потовщені насінні луски, які згодом дерев'яніють і зростаються з покривними. Вони мають складну будову і є гомологічними цілому пазушному пагону, в склад якого входять і мегаспорофіли, зрослі з віссю пагона. Тому жіноча шишка — видозмінена система пагонів. На верхньому боці насінної луски утворюється два насінних зачатки. Насінні зачатки мають таку ж будову, як у саговників. Мегаспорангій (нуцелюс) так само одержує додатковий захист і додаткове живлення за рахунок інтегумента. З чотирьох мегаспор лише одна проростає і утворює в нуцелюсі жіночий гаметофіт з двома архегоніями — первинний ендосперм, що складається з запасаючої паренхіми з великою кількістю поживних речовин, переважно олій. Архегонії так само мають яйцеклітину, над нею — черевцеву канальцеву клітину, в шийці — кілька шийкових канальцевих клітин.
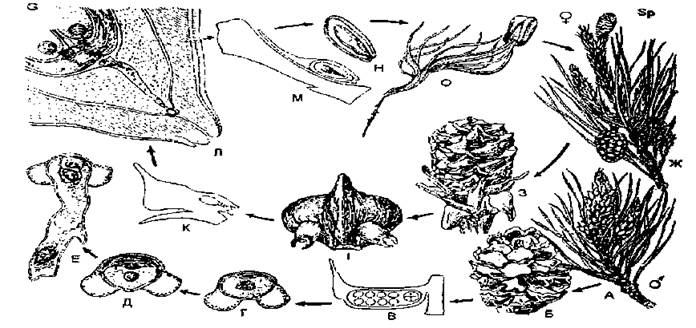
Рис. 13. Цикл розвитку сосни звичайної (Pinus sylvestris): А — гілка з чоловічими шишками; Б — чоловіча шишка; В — розріз мікроспорангія; Г-Е — проростання пилку; Ж — гілка з жіночими шишками; З — жіноча шишка; І — мегаспорофіл з двома насінними зачатками; К — мегаспорофіл збоку: Л — верхівка насінного зачатка; М, Н — насінина в розрізі; О — проросла насінина.
Чоловіча шишка на короткій осі має багато мікроспорофілів — тонких плівчастих лусок із двома мікроспорангіями на нижньому боці. В мікроспорангіях з археспорія формуються тапетум та спорогенна тканина. В результаті мейозу з клітин спорогенної тканини утворюються численні мікроспори, які вкриваються екзиною за рахунок тапетума. Екзина відстає від інтини, і з'являються два повітряних міхури, які забезпечують перенесення спори повітрям. Ще в мікроспорангії спори проростають, даючи дуже редуковані чоловічі гаметофіти, котрі не виходять за межі оболонки спори. Спершу відчленовуються дві маленькі протапіальні клітини, які швидко руйнуються, — вегетативні клітини заростка. Велика клітина ще раз ділиться, утворюючи антеридіальну (заміняє антеридій) і сифоногенну клітину пилкової трубки, яку неправильно називають вегетативною клітиною чоловічого заростка. Тепер спори перетворились на чоловічі гаметофіти (пилок) і висипаються з розкритого мікроспорангія в травні. Такі двоклітинні пилинки потрапляють на жіночі шишки відбувається запилення. Сифоногенна клітина на поверхні нуцелюса починає рости, утворюючи пилкову трубку, яка спершу лише прикріплює чоловічий заросток до нуцелюса.
У насінних зачатках жіночих шишок першого року життя спочатку сформована лише мегаспора, а жіночий заросток з архегоніями ще не утворився. В пилинці ще відсутні сформовані чоловічі гамети. Отже, запліднення не може відбуватися відразу після запилення. Луски в шишці після запилення щільно змикаються, і подальший розвиток заростків відбувається всередині шишки. Запліднення відбувається лише наступної весни, в зеленій шишці.
Перед заплідненням антеридіальна клітина пилинки ділиться і дає дві чоловічі гамети без джгутиків. Кінець пилкової трубки проникає в архегоній, спермії виходять з неї, і один з них запліднює яйцеклітину. Після запліднення насінний зачаток перетворюється в насінину. Із зиготи утворюється зародок нового спорофіта, який за допомогою підвіска заглиблюється в тканину жіночого заростка. Сам заросток розростається, заповнюється поживними речовинами і стає ендоспермом насіння. Таким чином, у жіночого гаметофіта сосни спостерігається зміна функцій: спершу статева, потім запасаюча. Заросток хвойних живе тільки за рахунок спорофіта. Отже, ендосперм голонасінних — це гаплоїдна тканина, яка утворюється задовго до запліднення.
У процесі розвитку зародка та ендосперму нуцелюс поступово руйнується, і в зрілому насінні від нього залишається лише плівочка. Покриви насінного зачатка, як і в саговників, перетворюються в щільну і шкірясту насіннєву шкірку. З прилягаючих тканин насінної луски утворюється прозоре крило, яке сприяє переносу стиглого насіння вітром. Отже, насіння голонасінних — це складний утвір: диплоїдна оболонка утворилася з мегаспорангія, гаплоїдний ендосперм — це заросток; диплоїдний зародок — дочірній спорофіт, який утворився із зиготи після запліднення.
Сформований зародок у стиглому насінні має добре виражені зачаткові вегетативні органи: гіпокотиль із зародковим корінцем, повернутим до мікропіле, і мутовку сім'ядольних листків-хвоїнок, які оточують верхівкову бруньку. Насіння в голонасінних відкрито сидить на насінній лусці, але під час дозрівання захищене щільно зімкнутими насінними лусками в шишках.
Отже, в сосни, як і в саговників, при детальному вивченні формування насіння виявляється замаскована антитетична зміна поколінь із значним переважанням спорофіта. Гаплоїдні гаметофіти різностатеві, дуже редуковані, живуть за рахунок спорофіта. Самостійно існує лише чоловічий гаметофіт — пилинка — у період від висипання з пиляка до моменту прикріплення до нуцелюса. Саме насіння є органом відтворення, розмноження та розселення рослин.
Підбиваючи підсумки, відзначимо, що в голонасінних спори втратили здатність до розселення і залишаються всередині спорангіїв. У процесі розвитку насіння змінюються функції мегаспорангія: всередині його розвивається гаметофіт і відбувається статевий процес. Отже, насіння із зародком — результат статевого процесу, а насіннєве розмноження слід вважати особливим типом статевого розмноження.