БОТАНІКА. МОРФОЛОГІЯ РОСЛИН - О. А. Шевчук - 2014
МОРФОЛОГІЯ КВІТКИ
Будова квітки
Квітка — це вкорочений видозмінений листкостебловий пагін з обмеженим ростом, який виконує функцію утворення спор і гамет та статевого розмноження, в результаті якого утворюється насіння і плід. Процес утворення квіток — антогенез.
Типова квітка покритонасінних або квіткових рослин закінчує головний або бічний пагін. Зустрічаються також поодинокі піхвові квітки, однак частіше квітки зібрані в суцвіття. У деяких рослин (вовчі ягоди, обліпиха та ін.) квітки утворюються безпосередньо на стовбурах або на старих бічних гілках — кауліфлорія.
Оскільки квітка — листкостебловий пагін, то вона має стеблову і листкову частини. Стеблова частина представлена квітконіжкою (видовжена частина квіткового пагона) і квітколожем (вкорочена розширена вісь). Квітколоже буває ввігнутим (слива, шипшина, черемха), плоским (півонія), опуклим (жовтець, малина), конічним (гравілат, ожина, магнолія) тощо. Біля основи квітконіжки або суцвіття може знаходитися верхівковий, часто видозмінений, листок — приквіток або брактея (пересиріч, бузок, конвалія). Квітка, яка не має квітоніжки, називається сидячою.
Листкова частина (квітколистки) представлена чашолистками, пелюстками, тичинками і маточками, які утворені плодолистками. Найчастіше вони розташовуються на квітколожі колом. Такі квітки називаються циклічними. Звичайно буває 5, або 4 кіл; 1 коло — чашолистки, 1 коло — пелюстки; 2, або 1 коло тичинок; 1 коло маточок. Ізомерна квітка містить однакову кількість елементів в кожному колі. Квітки з різною кількістю елементів у кожному колі називаються гетероциклічними (губоцвіті, хрестоцвіті).
При спіральному розташуванні частин квітки, квітка називається ациклічною (магнолієві, жовтецеві). Проміжне положення займають квітки геміциклічні, у яких спостерігається колове розташування одних частин і циклічне — інших.
Чашечка — це сукупність чашолистків квітки, які її утворюють. В типовому випадку чашечка зеленого кольору і виконує функцію захисту внутрішніх частин квітки від висихання та впливу високих або низьких температур, особливо до її розкривання (вишня, горох, розові, мальвові та ін.). В окремих випадках, у зв'язку з приваблюванням комах, чашечка яскраво забарвлена (гравілат, живокіст, фуксія, сокирки, зозулині черевички). У деяких рослин (суниці, бавовник, гравілат, перстачі) чашечка складається з двох кіл листочків, нижній з яких утворює підчашу.
Підчаша утворюється з приквітків (мальвові) або прилистків (розові).
Чашечка буває вільнолистою (жовтець, редька) та зрослолистою (горох, картопля, кукіль). У зрослолистій чашечці виділяють трубочку чашечки та зубчики. В залежності від ступеня зростання чашолистків виділяють кілька типів чашечки: розсічена чашечка (чашолистки зростаються тільки основою); роздільна чашечка (чашолистки зростаються майже до половини); лопатева чашечка (чашолистки зростаються на 2/3 їх довжини); зубчаста чашечка (вільними залишаються тільки верхівки чашолистків). За кількістю зубчиків можна визначити кількість чашолистків які утворюють чашечку. Чашечка може мати трубчасту, двогубу та дзвоникувату будову.
За типом симетрії чашечка буває актиноморфна та зигоморфна.
Віночок — це сукупність пелюсток квітки. Він служить для приваблювання запилювачів. Різноманіття типів віночків дуже значне, їх розрізняють за кольором, інтенсивністю забарвлення, за кількістю членів, формою пелюсток, розміром, взаєморозташуванням, ступенем зростання тощо.
Віночок буває вільнопелюстковим (мак, тюльпан, роза, жовтець) та зрослопелюстковим (картопля, гарбуз, огірок, примула, дзвоники). У представників еволюційно просунутих родин з вільнопелстковим віночком, пелюстки диференційовані на вузьку нижню частину — нігтик і верхню розширену — пластинку. Пластинки пелюсток розташовані під прямим кутом до осі (гвоздичні, капустяні). Така пелюстка називається нігтиковою. Якщо основа пелюстки широка, вона називається сидячою (жовтецеві, розові).
Пелюстки можуть мати розгалуження в напрямку повздовжньої осі, які утворюють зубчастість, або надрізаність пелюсток (двозубчасті, багатозубчасті тощо). В напрямку перпендикулярному до поверхні, пелюстки часто утворюють різноманітні вирости (на межі нігтика та відгину): привіночок — коронка (нарцис, підсніжник); кільце волосків (глуха кропива пурпурова).
У зрослопелюстковому віночку виділяють зрослу частину пелюсток - трубочку, незрослу — відгин та місце переходу трубочки у відгин — зів. Відгин має певну кількість зубчиків або лопатей в залежності від кількості пелюсток, що його утворюють. Але, наприклад, 4-х лопатевий віночок представників роду вероніка утворений 5-ма пелюстками. В зіві віночка нерідко виникають різні лусочки і валики (особливо у шорстколистих), а переважно у роздільнопелюсткових віночків іноді виникають привіночки або коронки, що теж є виростами на пелюстках. Вони, очевидно, підсилюють привабливість віночка для запилювачів.
У багатьох випадках у таких родин, як руткові, жовтецеві, фіалкові, бальзамінові, зозулинцеві та інші (в зв'язку зі спеціалізованим запиленням) із пелюстки або листочка простої оцвітини формується шпорка. На початкових етапах розвитку квітки з'являється мішковидне випинання пелюстки або листочка оцвітини, яке пізніше видовжується і перетворюється на шпорку. Виникнення шпорок знаходиться в тісному зв'язку з утворенням нектару, який виділяється стінками самої шпорки або нектарниками, які знаходяться в її середині.
Актиноморфні (полісиметричні) зрослопелюсткові віночки класифікуються за довжиною трубочки, формою та величиною відгину: колесоподібний — трубочка мала або відсутня, відгин розвернутий у площину (картопля, вербозілля, незабудка); лійкоподібний — трубочка велика лійкоподібна, відгин порівняно невеликий (тютюн, кручені паничі, березка польова, дурман);
дзвоникоподібний — трубочка сферична, чашоподібна, поступово переходить у малопомітний відгин (дзвоники, конвалія); трубчастий — трубочка циліндрична без відгину або з коротким відгином (центральні квітки кошика соняшника, осоту, ромашки, нагідок); блюдцеподібний — трубочка циліндрична з широким відгином (бузок); ковпачковий — пелюстки зростаються верхівками (виноград).
Серед зигоморфних (моносиметричних) зрослопелюсткових віночків найбільш часто зустрічаються такі: двогубий — відгин складається з двох неоднакових губ — верхньої та нижньої (губоцвіті); язичковий — від трубочки відходять зрослі пелюстки, які мають вигляд язичка: справжньоязичковий (квітки кошика кульбаби, осоту); несправжньоязичковий (крайові квітки кошика соняшника, нагідок, ромашки, осоту польового); шпоркоподібний — пелюстки утворюють порожній виріст, який називається шпоркою, буває і у вільнопелюсткових зигоморфних і актиноморфних віночків (сокирки, льонок).
Асиметричний віночок мають канни, валеріана, орхідеї, каштан кінський та ін.
Чашечка і віночок разом утворюють оцвітину. Це стерильна (безплідна) частина квітки, що виконує захисну функцію, а також функцію приваблювання комах-запилювачів.
Походження віночка. Скоріше за все пелюстки виникли з видозмінених тичинок. Відмінною ілюстрацією перетворення тичинок в пелюстки можуть бути види латаття та глечики жовті. У цих видів спостерігаються всі перехідні ступені від крайніх зовнішніх пелюсток до самих внутрішніх тичинок. Внутрішні пелюстки у них можна розглядати як перехідні від тичинок. Органи, які втратили здатність до формування пилку, але ще зберегли деякі морфологічні риси тичинок, називають стамінодіями.
Оцвітина. Виділяють такі типи оцвітини: проста (перигоній), якщо вона складається з однакових листочків. Якщо оцвітина яскраво забарвлена і утворена тільки пелюстками то її називають простою віночковидною (тюльпан, гречка, півники, зірочки, підсніжник, конвалія, калюжниця); а якщо має зелене забарвлення і утворена тільки чашолистками — простою чашечковидною (лобода, буряк, щавель, коноплі). Квітка, яка має просту чашечковидну оцвітину називається безпелюстковою, або апетальною. Квітка з простою оцвітиною називається однопокривною.
Подвійна оцвітина утворена різними за зовнішнім виглядом та забарвленням чашечкою та віночком (троянда, вишня, яблуня, горох, квасоля, мак та ін.). Квітка, у якої оцвітина диференційована на чашечку та віночок називається гетерохламідною або дихламідною. У деяких рослин оцвітина плівчаста (верба, злаки). Якщо оцвітина у квітці відсутня то таку квітку називають голою, апохламідною або ахламідною (ясен, верба, образки). Квітки деяких видів тонконогових мають остюк — тонкий загострений виріст на нижній квітковій або на колосковій лусці та лодикули — плівчасті луски, що сприяють розкриванню квітки.
Оцвітина в основному відіграє захисну роль для тичинок і маточок та служить «рекламою», яка приваблює комах-запилювачів. Взаємопоєднана еволюція цих двох ліній (комах і квіткових рослин) розвитку органічного світу з різних царств давала обом групам відповідні взаємовигоду. Оскільки для великої групи комах квітка є основним постачальником продуктів їх харчування (нектар забезпечує в основному вуглеводну частину корму, а пилок - білкову та жирову), а рослина за це має гарантоване продовження свого роду у вигляді цілеспрямованого процесу запилення, а значить і запліднення. Крім приємного запаху та наявності поживного нектару чи пилку квітка в переважній більшості має яскраве забарвлення найрізноманітніших кольорів, що без сумніву є одним із факторів приваблення.
Найрізноманітніше забарвлення квіток а саме, їх пелюсток, зобов’язане, перш за все пігментам, найважливіше місце серед яких належить антоціану (грецького anthos — квітка і kianos блакить, лазур).
Квітки багатьох рослин мають жовте забарвлення, яким вони найчастіше зобов'язані пігментові антохлору (грецького anthos — квітка, hloros — зеленувато-жовтий), який маємо у квітках анемони жовтецевої, первоцвіту весняного, дивини, жовтеців та. інших видів рослин.
Білий колір пелюсток у багатьох рослин зумовлений відсутністю пігментів і наявністю у пелюстках дрібних повітряних порожнин, від яких відбиваються всі промені сонячного спектру. Немало випадків, коли у квітці поєднується кілька контрастуючих барв. Наприклад, великі чорні плями при основі пелюсток у маків і тюльпанів, або контрасність кольорів у квітках деяких лілій, багатьох орхідей, в тому числі і позатропічних. А в кошиках багатьох рослин з родини айстрових серединні квітки, як правило, мають жовтий колір, тоді як крайові, язичкові — білий, фіалковий чи синій. Інколи такі крайові квітки, наприклад, у суцвіттях багатьох айстрових, а також у суцвіттях калини при своїх розмірах і яскравості навіть стають стерильними, безплідними, ніби "жертвуючи" цим заради успіху запилення інших численних і менш помітних квіток цього ж суцвіття.
Останнім часом деякі ботаніки схиляються до думки, що різноманітний колір оцвітини в квітках служить не стільки засобом приваблення комах-запилювачів, скільки забезпечує і оптимізує тепловий баланс для повноцінного розвитку найбільш важливих для рослини елементів квітки — маточки, в зав'язі якої формуються насінні зачатки, а на приймочках відбувається проростання пилкових зерен. І в цьому випадку особлива роль відводиться антоціану, оскільки дослідженнями встановлено, що його наявність в листочках оцвітини взагалі, чи в пелюстках зокрема, підвищує температуру в квітці (чи в суцвітті), а отже захищає квітку від малоефективних відносно понижених температур, які скоріше за все є недостатніми для протікання відповідних фізіологічних і біохімічних процесів, які відбуваються при формуванні і розвитку насінних зачатків і пилку, та його подальшого розвитку на приймочці маточки і, особливо, процесу подвійного запліднення, що відбувається в кожному з насінних зачатків зав'язі. Окремо слід зауважити, що до такої думки нас приводить і той факт, що оцвітина квітки завдяки її конструкції часто діє як система своєрідних дзеркал, що концентрують у центральній її частині потік променів (в залежності від кольору і форми оцвітини) відповідної довжини, сили і інтенсивності, що формує не лише відповідний температурний, а й взагалі специфічний, енергетичний баланс таких важливих точок організму, квіткової рослини, де протікають чи не найважливіші для відтворення наступних поколінь процеси.
Махрові або повні квітки. Для них характерне значне збільшення кількості пелюсток, які виникають внаслідок перетворення у пелюстки тичинок, плодолистків, а іноді і інших частин квітки під дією різних факторів. Таким чином, вихідним матеріалом для селекції махрових форм є квітки з великою кількістю тичинок, як наприклад, у троянди або петунії.
Видозмінами листка у квітці є також тичинки і маточки. Це головні складові частини квітки як органу розмноження.
Андроцей — це сукупність тичинок квітки. Кількість тичинок може бути 1 (мономерний андроцей — канна) або 2 (бузок), дорівнювати кількості елементів оцвітини (лілія, підсніжник), бути в 2 або багато разів більшою (шипшина, вишня, груша, жовтець). Як правило, кількість тичинок постійна для роду. Він може бути вільним, якщо тичинки не зрослися між собою (тюльпан), або зрослим якщо тичинки зрослися між собою (бобові). Якщо зростаються всі тичинки, андроцей називають однобратнім (вербозілля, зіновать). Якщо одна тичинка залишається вільною — двобратнім (лядвенець, горох). Якщо тичинки зростаються в кілька груп — багатобратнім (у видів розових, пасльонових, лілійних та ін.).
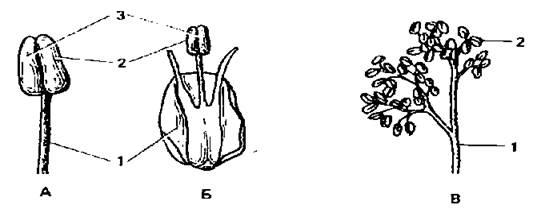
Рис.14. Будова тичинки: А - проста тичинкова нитка (троянда); Б - тичинкова нитка з зубцями (цибуля); В - гілляста тичинкова нитка (кліщовини); 1 - тичинкова нитка; 2 - пиляк; 3 - в’язальце.
Дуже рідко нитки тичинок зростаються з стовпчиком маточки, утворюючи так звану колонку, або гімностемій (зозулинцеві). У одних рослин зростаються тичинкові нитки — синандрія (горох, бавовник, види роду гарбуз), у інших — пиляки (кульбаба, соняшник, васильки), у багатьох рослин тичинки зростаються з пелюстками (цикорій, примула).
Тичинка — це видозмінений мікроспорофіл. Мікроспорофіл — це листок, який несе мікроспорангій. Мікроспорангій — вмістище мікроспор. З мікроспор утворюється пилок або чоловічий гаметофіт. Тичинка має тичинкову нитку, пиляк і в'язальце.
Тичинкові нитки у більшості рослин прості, не галузяться (троянда, тюльпан, вишня, пшениця та ін.). Однак, інколи вони мають бічні вирости різної форми (наприклад, тичинкові нитки цибулі мають зубчики) або галузяться (рицина). Розгалуження тичинкової нитки призводять до утворення складних тичинок (береза, ліщина). Якщо тичинкова нитка відсутня, то тичинки (пиляк) називають сидячими (фіалка).
На поперечному розрізі тичинкова нитка може бути округлої або овальної форми (шипшина, цибуля).
У типовому випадку пиляк складається з двох половинок — тек, у кожній з яких утворюється два пилкових гнізда. Тканина, що з’єднує між собою половинки пиляка, називається в'язальце. Пилкові гнізда встелені покриваючим шаром — тапетумом і в них знаходиться археспоріальна тканина з якої в результаті мікроспорогенезу утворюються мікроспори. Отже, пилкові гнізда — це видозмінені мікроспорангії. Спосіб прикріплення пиляка до тичинкової нитки у різних рослин різний. У злаків, лілій пиляк прикріплюється до тичинкової нитки серединою, у тюльпана, осок — основою. В’язальце може бути коротким (тонконогові), або довгим (фіалки, вороняче око).
Розгалужені (складні) тичинки несуть тільки половинну кількість пилкових гнізд, оскільки внаслідок розгалуження мають тільки одну теку. Інколи і проста тичинка може мати одну теку (барбарис). Монотекова тичинка може утворюватись внаслідок дегенерації перегородки або тканини в'язальця. Тичинка омели має до 50 пилкових гнізд. Тичинкова нитка, на якій міститься кілька пиляків, називається антерофор.
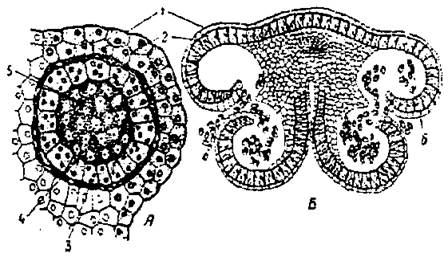
Рис. 15. Пиляк в поперечному розрізі: А - одне пилкове гніздо з археоспоріями; Б - пиляк : 1 - епідерма; 2 - фіброзний шар; 3 - дегенеруючий шар; 4 - тапетум; 5 - археоспорій (спорогенна тканина); 6 — пилок.
Тичинки, які втратили здатність утворювати пилок, але зберегли морфологічні ознаки тичинок називаються стамінодіями (льон). Іноді вони набувають вигляду яскраво забарвлених пелюстковидних пластинок (канна, гвоздики, махрові троянди). Стамінодії можуть перетворюватись на нектарники (купальниця).
За висотою відносно одна одної, тичинки можуть бути однаковими — односильними (тюльпан), або різними — двосильними, коли дві тичинки вище інших (губоцвіті); трисильними; чотирисильними (хрестоцвіті або капустяні).
Гінецей — сукупність плодолистків (мегаспорофілів) квітки, які утворюють одну або декілька маточок. Гінецей, який складається з однієї маточки, називається простим (сокирки), а з кількох — багатьох маточок — складним (магнолія, сусак).
Маточка — закрите вмістище для насінних зачатків, складається з приймочки (верхня розширена частина), стовпчика або стилодій (у апокарпній маточці) (середня циліндрична частина), зав'язі (розширена нижня частина). Форма приймочки може бути різноманітна, поверхня — липкою, на ній можуть міститися сосочки, волоски, що сприяє кращому затриманню пилку. Якщо стовпчик відсутній, приймочка називається сидячою. В зав'язі утворюються одна чи декілька порожнин (гнізд), в яких розвиваються насінні зачатки. Гінецей складається із стількох плодолистків (карпел), скільки окремих стовпчиків несе зав'язь, або скільки лопатей має приймочка, або швів має зав'язь. Для морфологічного аналізу слід використовувати всі ці ознаки. У конопель, деяких видів верби відбувається карпелізація тичинок — перетворення їх у плодолистки.
Зав’язь, по відношенню до інших частин квітки, може бути верхньою та нижньою. Верхня зав'язь розташовується вільно на плоскому, ввігнутому чи опуклому квітколожі і утворена тільки плодолистками (горох, вишня, жовтець), її можна легко відділити від квітколожа препарувальною голкою (тонконогові, горох, помідори та ін.). В утворенні нижньої зав'язі, крім плодолистків, беруть участь також інші частини квітки — основи чашолистків, пелюсток, тичинок, рідше квітколоже, з якими вона зростається (яблуня, огірок). Відділити препарувальною голкою таку зав'язь не можна. Виділяють також напівнижню зав'язь. Принаймні нижня частина такої зав'язі зростається з іншими частинами квітки (жимолость, бузина), а верхня залишається вільною.
Розширене бокаловидне квітколоже, з яким зростаються своїми основами листочки оцвітини та тичинки, називається гіпантієм. Гіпантій характерний для квіток, що мають напівнижню або верхню зав'язь (шипшина).
Виділяють одногнізду (якщо гнізда зав'язі сполучені між собою), двогнізду, тригнізду і багатогнізду зав'язі.
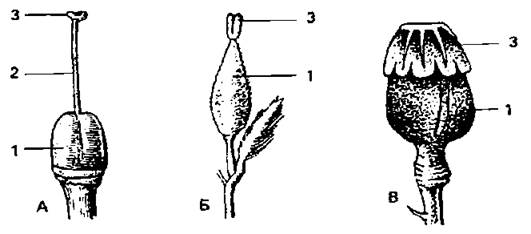
Рис. 16. Гінецей: А - моркви, Б - верби, В - маку: 1 - зав’язь, 2 - стовпчик, 3 - приймочка.
Типи плацентації. Порядок розміщення насінних зачатків на плодолистку називається плацентацією. В залежності від розташування насінних зачатків виділяють кілька типів плацентації. Пристінна плацентація — насінні зачатки прикріплюються до внутрішньої стінки зав'язі. До пристіної плацентації належать парієтальна плацептація, коли насінні зачатки розташовані уздовж швів плодолистків (види родин вербових, гарбузових), і медіанна плацентація, коли насінні зачатки розташовуються уздовж центральної жилки плодолистка (жовтецеві, ломикаменеві). Кутова (маргінальна) плацентація — насінні зачатки розташовуються по краях плодолистків, у центрі замкнутої багатогніздої зав'язі (лілійні). Колончаста плацентація — насінні зачатки знаходяться на колончастому утворенні, сформованому зрослими плацентами і розташованому в центрі зав'язі (гвоздичні). Сліди провідних пучків на плацентах мають характерне розташування, по якому після опадання насіння (по дозріванні) легко визначають плацентарну тканину, що демонструє тип плацентації даної рослини.
За характером зростання плодолистків розрізняють апокарпний та ценокарпний гінецеї.
Апокарпним називають гінецей, що складається з одного плодолистка і утворює одну маточку — простий гінецей (горох), або з декількох плодолистків (складний гінецей), які не зростаються між собою і утворюють багато маточок (сусак, жовтець). Складний гінецей завжди апокарпний тому, що кожна його маточка складається з одного плодолистка. В еволюційному відношенні — це найпримітивніший тип гінецею, який характерний для представників родин магнолієвих і жовтецевих. Ценокарпним називають гінецей, що складається з кількох зрослих плодолистків. За характером зростання плодолистків та числом гнізд зав'язі розрізняють три підтипи ценокарпного гінецею: синкарпний — утворений декількома глибоко зрослими плодолистками, що утворюють багатогнізду зав'язь (в результаті бічного зростання) і мають кутову (маргінальну) плацентацію (лілія, тюльпан); паракарпний — сформований декількома неглибоко зрослими плодолистками, які утворюють одногнізду завязь (мак, гарбуз, аґрус) і має пристінну плацентацію; лізикарпний — сформований декількома плодолистками, які утворюють одногнізду зав'язь. Для нього характерна колонка, що піднімається із дна зав'язі і нібито є продовженням квітколожа (гвоздика, смілка). Плацентація колончаста.
Наявність у квіток рослин одного виду стовпчиків маточки і тичинкових ниток однакової довжини називається гомостилією (рівностовпчастість). Гомостилія характерна для більшості квіткових рослин. Коли довжина стовпчиків маточок і тичинкових ниток різна, спостерігається різностовпчастість — гетеростилія (медунка, гречка, первоцвіт, бузок).
Типи квіток
За типом симетрії, особливо віночка, виділяють такі типи квіток: актиноморфні (полісиметричні); зигоморфні (моносиметричні); асиметричні. Квітка, через яку можна провести не менше двох площин симетрії, має радіальну симетрію (вишня, слива, конвалія, мак та ін.), і називається правильною або актиноморфною. Моносиметричні квітки, через які можна провести лише одну площину симетрії, називають неправильними або зигоморфними (бобові). Асиметричною називається квітка, через яку не можна провести площини симетрії (зозулинцеві, канни).
Іншою важливою ознакою квітки є її статевість. За цією ознакою розрізняють: двостатеві (гермафродитні) квітки, в яких є тичинки і маточки; одностатеві квітки, які мають або тичинки, або маточки. Відповідно до цього розрізняють тичинкові (чоловічі) і маточкові (жіночі) квітки. Рослини з різностатевими квітками на одній і тій же особині називають однодомними (кукурудза, дуб, береза). Рослини, що мають тичинкові та маточкові квітки на різних особинах, називають дводомними (тополя, верба). Рослини, в яких поряд з двостатевими квітками зустрічаються й одностатеві, називаються багатодомними, або полігамними. Для деяких видів рослин характерним є розвиток на одних особинах тільки тичинкових квіток, на інших — двостатевих. Таке явище називається андродієція або андроеція, а такі рослини — андродійокістами (щавель). Інколи тичинкові та двостатеві квітки утворюються на одній і тій же рослині (чемериця, підмаренник). Таке явище одержало назву андромонеція. Аналогічне явище, коли на одних особинах є двостатеві квітки, а на інших — лише жіночі, називається — гінодієція (жіноча дводомність). При наявності на одній рослині двостатевих та жіночих квіток спостерігається гіномонеція (жіноча однодомність). У конопель чоловіча особина, на якій утворюються лише тичинкові квітки називається плоскінь. Вона відрізняється від жіночої особини — матірки тоншим стеблом з меншою кількістю листків.
Не менш важливою ознакою квітки є положення зав’язі. Зав'язь може бути верхньою, нижньою, напівнижньою. Залежно від положення зав’язі, що до інших частин квітки, розрізняють квітку: підматочкову, якщо вона має верхню зав'язь, що розміщується вільно на плоскому, опуклому або ввігнутому квітколожі, її стінки утворені лише стінками плодолистків, а тичинки та інші частини квітки прикріплюються нижче маточки (жовтецеві, лілійні); напівнадматочкову, якщо вона має напівнижню зав'язь, що зростається з квітколожем знизу і вільна лише у верхній частині, а оцвітина відходить від середини зав'язі (бузина, калина); надматочкову, якщо вона має нижню зав'язь, в утворенні якої, крім плодолистків, беруть участь також інші частини квітки, здебільшого основа чашолистків і пелюсток, з якими вона зростається (яблуня, груша, гарбузові); навколоматочкову квітку, якщо вона має напівнижню зав'язь, що сидить на дні ввігнутого бокаловидного квітколожа — гіпантію (шипшина). Мономерна квітка не має оцвітини і містить або тільки маточки, або тільки тичинки (деякі види родів тополя, верба та ін.).
Формула та діаграма квітки
Формула квітки — це вираження будови квітки за допомогою символів, букв та цифр. Використовують такі позначення при написанні формули квітки:
✵ * або + — актиноморфна квітка;
✵ ↑ або ↓ — зигоморфна квітка;
✵ % - асиметрична квітка;
✵ ![]() - чоловіча квітка;
- чоловіча квітка;
✵ ![]() - жіноча квітка;
- жіноча квітка;
✵ ![]() двостатева квітка;
двостатева квітка;
✵ P (Perigonium) — покриви квітки або проста оцвітина;
✵ Са (Саlух — лат.), або К (Кеlch — нім.) — чашечка;
✵ Со або С (Соrоііа) — віночок;
✵ (Androeceum) — андроцей;
✵ G (Gynoeceum) — гінецей;
✵ + — наявність двох або декількох кіл, головним чином в оцвітині або андроцеї;
✵ ( ) — зростання;
✵ (-), (-), (-) — верхня, нижня та напівнижня зав'язь.
Кількість членів кожної частини квітки позначають цифрами. Велику (більше 12) та невизначену кількість членів квітки позначають знаком невизначеності — ∞. Дуже рідко кількість членів андроцея або гінецея буває невизначеною в межах 12 (глід). Їх кількість також позначається знаком невизначеності. У випадку зростання між собою членів квітки, їх кількість укладається в дужки. Якщо чашечка, віночок або тичинки розташовані кількома колами, то цифри, що позначають їх кількість в окремих колах поєднуються значком +. Формула повинна відображати кількість плодолистків, що утворюють гінецей. Якщо їх декілька, то слід зазначити зростаються вони між собою (ценокарпний гінецей) чи кожний плодолисток утворює окрему маточку (апокарпний гінецей), а також тип зав'язі: верхня чи нижня.
Наприклад, формула квітки вишні — *K5C5A∞G1. Характеристика такої квітки читається так: квітка правильна, двостатева, чашечка з 5 вільних чашолистків, віночок з 5 вільних пелюсток, тичинок багато, маточка 1, зав'язь верхня.
Діаграма квітки — це схематична проекція квітки на площину, перпендикулярну до осі квітки. Діаграма демонструє не тільки наявність частин квітки та кількість її членів, але і розташування їх відносно один одного. Тобто діаграма дає більш повне уявлення про будову квітки. Прийнято єдиний спосіб орієнтації діаграми: вісь квітки нагорі, а криючий листок знизу. Члени квітки позначаються завжди чітко визначеними значками. Чашолистки на діаграмі позначаються дужкою з кілем — {}; пелюстки — круглими дужками — (); тичинки — у вигляді розрізу через пиляк (якщо тичинок багато то у вигляді затушованого еліпса); гінецей у вигляді поперечного розрізу зав'язі або зав'язей з плацентацією та насінними зачатками. На діаграмі квітки позначаються також приквітки та вісь суцвіття у вигляді точок. У випадках, коли окремі члени квітки зростаються, знаки, якими вони позначаються на діаграмі, з'єднуються дугами чи прямими лініями.
Насінний зачаток
Видозмінені мегасинангії, які у більшості насінних рослин в результаті зростання мегаспорофілів потрапивши всередину зав'язі, називаються насінними зачатками, що мають короткі насінні ніжки, або фунікулус: якими вони прикріплюються в зав'язі до стінок останньої. Місце прикріплення насінних зачатків до тканин зав'язі називають плацентами. Місце з'єднання насінної ніжки з насінним зачатком називають рубчиком.
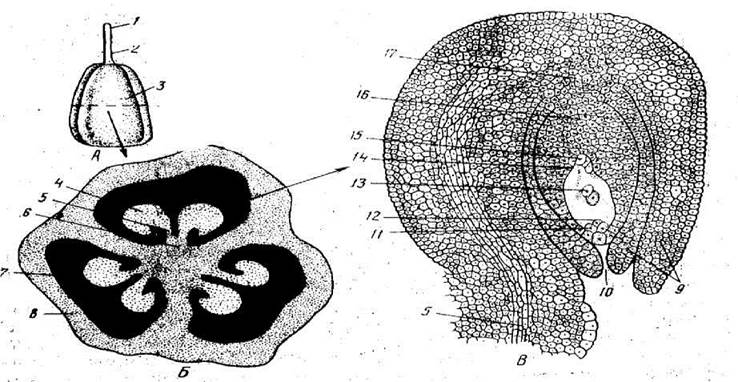
Рис. 17. Поперечний розріз насінного зачатка проліска: А - загальний вигляд, Б - поперечний розріз зав’язі, В - насінний зачаток: 1 - приймочка; 2 - стовпчик; 3 - зав’язь, 4 - насінний зачаток; 5 - фунікулус; 6 - плацента; 7 - гніздо; 8 - стінка зав’язі; 9 - інтегумент; 10 - мікропіле; 11 - синергіди; 12 - яйцеклітина; 13 - вторинне ядро центральної клітини; 14 - зародковий мішок; 15 - антиподи; 16 - нуцелус, 17 - халаза.
Насінний зачаток складається з нуцелуса (ядра в широкому розумінні), вкритого одним або двома шарам інтегументів (покривів), кінці яких з одного боку насінного зачатка, не зростаючись, утворюють вузькі канал (мікропіле), через який пилкова трубка проникає до зародкового мішка. Протилежну по відношенню до мікропіле частину насінного зачатка, де нуцелус і інтегументи зростаються між собою, називають халазою.
Розрізняють три основних типи насінних зачатків: прямі (ортотропні), обернені (анатропні), зігнуті (кампілотропні), а також гемітропний та амфітропний. У прямого насінного зачатка нуцелус є прямим продовженням фунікулуса (насінної ніжки), а спрямоване догори мікропіле знаходиться на одній осі з рубчиком, і з фунікулусом. Характерний для перцевих, гречкових, ароїдних. У оберненого насінного зачатка (при прямій насінній ніжці) мікропіле виявляється повернутим донизу (майже на 80°), так що останнє розміщується поряд з рубчиком і повернуте майже до плаценти, дуже поширений тип насінних зачатків і притаманний магноліїдам, таким чином, можливо є вихідним. Кампілотропний або односторонньо зігнутий насінний зачаток характеризується одностороннім розростанням і нуцелуса, і інтегументів. Зустрічається у бобових, лободових та інших. Гемітропний, або напівповертутий насінний зачаток, у якого нуцелус та інтегументи повернуті по відношенню до плаценти на 90°. Займає ніби проміжне положення між ортотропним та анатропним насінними зачатками (окремі представники ранникових, первоцвіті). У окремих представників тих родин і порядків, де є звичайними кампілотропні насінні зачатки, нуцелус має підковоподібне зігнуту форму і називається амфітропним, або двосторонньо зігнутим. Інтегументи, які входять до складу насінного зачатка у більшості родин подвійні (переважна більшість вільнопелюсткових дводольних і однодольні). Навпаки, зрослопелюсткові дводольні мають, як правило, один інтегумент.
В нуцелусі, або ядрі, насінного зачатка в подальшому відбуваються процеси, які ведуть до утворення насінини. За морфологічною природою нуцелус є мегаспорангієм, перетвореним в процесі еволюції з мегаспорангія далеких предків покритонасінних — палеозойських різноспорових папоротеподібних, які мали багатошарові спорангії. Еволюція нуцелуса покритонасінних пов'язана зі зменшенням товщини його стінок. Розрізняють товстостінні (крассинуцелятні) і тонкостінні (тенуінуцелятні) насінні зачатки.
Розвиток насінного зачатка і мегаспорогенез
Насінний зачаток виникає на плаценті у вигляді напівкулеподібного горбочка, який спочатку складається з однорідних меристематичних клітин. Через деякий час поблизу верхівки насінного зачатка з'являється одна або декілька археспоріальних клітин, які розвиваються з субепідермального шару. Приблизно в цей час біля основи насінного зачатка виникають інтегументи у вигляді одного або двох кільцевих валиків. Найчастіше в насінному зачатку під пилковходом одна із субепідермальних клітин нуцелуса — археспоріальна клітина починає збільшуватися, а потім поділяється способом мейозу. Виникає тетрада (четвірка) гаплоїдних (п) мегаспор, на цьому і закінчується мегаспорогенез. З чотирьох мегаспор виживає тільки одна, всі інші поступово зменшуються і зникають. Мегаспора, яка лишилася, починає рости. Ядро її тричі поділяється способом мітозу. Таким чином, з мегаспори утворюється восьмиядерний зародковий мішок(цитокінезу або формування клітинних оболонок під час поділу тут не буває). Вісім гаплоїдних ядер, які виникли, знаходяться спочатку по чотири в двох протилежних кінцях зародкового мішка (мікропілярному і халазальному). Між ними, звичайно, є вакуоля. Потім по одному ядру від кожної четвірки пересувається до центра клітини. Ці полярні ядра, з'єднавшись, утворюють диплоїдне ядро, яке називається центральним, або вторинним ядром зародкового мішка, в результаті чого цей мішок стає семиядерним. Центральне ядро покривається цитоплазмою і стає центральною клітиною зародкового мішка (іноді полярні ядра зливаються пізніше). Біля мікропілярного кінця зародкового мішка утворюється так званий яйцевий апарат з трьох клітин, які виникли з трьох ядер, навколо яких сконцентрувалася цитоплазма. Одна з цих трьох клітин стає яйцеклітиною, або жіночою гаметою, а дві інші — синергідами (допоміжними клітинами).
На халазальному кінці зародкового мішка формуються ще три клітини — антиподи. Утворений таким чином зародковий мішок із сімома клітинами тепер уже готовий до процесу запліднення. Такий тип утворення зародкового мішка трапляється найчастіше, але в природі існують і інші типи його розвитку. Порівняно з усіма голонасінними, у покритонасінних жіночий гаметофіт сформований дуже незначною кількістю клітин і тому відрізняється прискореним розвитком. Замість десяти або одинадцяти поділів, характерних для голонасінних, розвиток зародкового мішка покритонасінних відбувається в результаті лише трьох- п'яти поділів, завдяки чому дуже скорочується час, необхідний для розвитку зародкового мішка. Немає сумніву в тому, що швидкість розвитку жіночих і чоловічих гаметофітів покритонасінних входить до ознак, які забезпечили цьому відділу панування в рослинному світі в сучасну геологічну епоху. На відміну від переважної більшості голонасінних, насінні зачатки покритонасінних вже не мають архегоніїв.
Подвійне запліднення у покритонасінних рослин
Суть подвійного запліднення була з'ясована у 1898 році професором університету святого Володимира Сергієм Гавриловичем Навашиним у Києві.
Потрапивши на приймочку маточки, пилкове зерно починає проростати. Цьому сприяють речовини, які виділяють приймочки, відповідні температура і вологість та інші умови зовнішнього середовища. В процесі проростання вміст пилкового зерна випинається крізь пору екзини у вигляді тонкої трубочки, вкритої лише інтиною. Ця пилкова трубка починає видовжуватись і проникає в тканину стовпчика. Тут вона просувається по спеціальному каналу або по міжклітинниках; потім досягає зав'язі і прямує до мікропіле насінного зачатка. Ріст пилкової трубки пов'язаний з обміном речовин, що відбувається між нею та внутрішніми клітинами. У більшості рослин пилкова трубка проникає до насінного зачатка крізь мікропіле (порогамія), у берези, вільхи деяких інших — крізь халазу (халазогамія), у в'яза — крізь інтегументи (мезогамія). Потім пилкова трубка проникає до зародкового мішка. За період росту всередині маточки вміст пилкової трубки зазнає певних змін. Вегетативне ядро, яке спочатку знаходилось біля ростучої частини трубки і сприяло її видовженню, поступово розсмоктується. Генеративне ядро поділяється способом мітозу на дві чоловічі статеві клітини — спермії (щоправда у деяких рослин це відбувається значно раніше, ще у непророслій пилинці). При стиканні з зародковим мішком відбувається розчинення стінки пилкової трубки, а, увійшовши до зародкового мішка, вона розривається. З двох сперміїв, які звільнилися, один прямує до жіночої клітини — яйцеклітини і зливається з нею. З диплоїдної зиготи, що виникла, утворюється зародок. Другий спермій зливається з центральною диплоїдною клітиною зародкового мішка, і триплоїдна клітина, яка утворилася пізніше, сформує живильну тканину — ендосперм.
У цьому злитті двох сперміїв з двома клітинами зародкового мішка і полягає суть подвійного запліднення, притаманного лише покритонасінним рослинам. Після цього синергіди та антиподи повністю відмирають. Насінний зачаток розростається і перетворюється в насінину.
Інтегументи, видозмінившись, сформують шкірку насінини. Зав'язь (а іноді і квітка в цілому) перетвориться в плід. Із стінок зав'язі сформується оплодень. Якщо в клітинах нуцелуса накопичуються поживні речовини, тоді утворюється особлива тканина — перисперм. З часу потрапляння пилинки на приймочку маточки до процесу подвійного запліднення у різних рослин проходить від 20-30 хв. до кількох діб. В результаті подвійного запліднення, як уже відмічалося, із зав'язі формується плід, а з її стінок — оплодень. Насінина ж формується з насінного зачатку, шкірка насінини — з інтегументів, перисперм — з нуцелуса, ендосперм — з центральної клітини, зародок — з заплідненої яйцеклітини.
Різні плоди мають свою специфіку будови. Таку ж специфіку будови має і насіння різних рослин.