БОТАНІКА. МОРФОЛОГІЯ РОСЛИН - О. А. Шевчук - 2014
МОРФОЛОГІЯ КОРЕНЯ
Визначення і функції кореня.
Корінь (radix) - здебільшого ортотропний, вегетативний, радіально- симетричний орган, з позитивним геотропізмом, негативним геліотропізмом, який закріплює рослину в ґрунті (субстраті), здатний до галуження та тривалого верхівкового росту.
Корені виконують різні фізіологічні і механічні функції:
✵ ґрунтового живлення рослин (поглинають з ґрунту і транспортують і мінеральні речовини);
✵ механічну або якірну (закріплюють рослини в субстраті);
✵ запасаючу (накопичують поживні речовини - морква, буряк, жоржини тощо);
✵ провідну (забезпечується провідними тканинами осьового циліндра);
✵ синтетичну (синтезують деякі органічні сполуки - амінокислоти, гормони росту, алкалоїди тощо);
✵ видільну (виділяють у навколишнє середовище коліна - слиз, органічні кислоти, амінокислоти, вуглекислий газ тощо, які сприяють розвитку мікрофлори та засвоєнню важкорозчинних сполук);
✵ вегетативного розмноження рослин (забезпечують вегетативне поновлення у разі наявності додаткових бруньок);
✵ симбіотичну (забезпечують взаємодію з коренями інших рослин, грибами та мікроорганізмами, за допомогою гормонів та фізіологічноактивних речовин);
✵ ґрунтоутворюючу (беруть участь у ґрунтоутворенні).
У процесі еволюції корінь виник як необхідне пристосування рослин до умов наземного існування. Філогенетично корінь найбільш молодий орган. Як підземний орган корінь сформувався в процесі еволюції пізніше за пагін у плауноподібних, однак лише у спорофіта. Найбільш ймовірно, що корінь виник від ризомоїдів, покритих ризоїдами, перших наземних рослин - риніофітів.
Коренів не було у псилофітів - предків більшості надземних рослин. Їх немає у мохоподібних.
До виникнення коренів функцію всмоктування виконували ризоїди. Це тонкі коренеподібні, безбарвні утвори, за допомогою яких рослини прикріплюються до субстрату і поглинають з нього воду та поживні речовини. Ризоїди утворюються у сучасних мохоподібних, лишайників, деяких водоростей тощо. Не дивлячись на відносну філогенетичну молодість, корінь - високоспеціалізований орган, який добре пристосований до умов існування і виконання своїх функцій. Найвищої диференціації і спеціалізації досягають справжні корені, властиві голонасінним і квітковим рослинам.
Корінь властивий всім сучасним вищим рослинам за винятком мохів, рослин-паразитів (повитиця, петрів хрест), напівпаразитів (омела) та деяких вищих рослин, пристосованих до життя у воді (пухирник, сальвінія).
Корінь відрізняється від пагона тим, що немає листків, вузлів і меживузлів; верхівка кореня (апекс) захищена кореневим чохликом; інтеркалярний ріст відсутній.
Це осьовий орган. Разом із стеблом він утворює єдину вісь рослини. Корінь характеризується здатністю до галуження, завдяки чому при відносно невеликому об’ємі він має велику зовнішню поверхню. Це збільшує контакт кореня з ґрунтом і полегшує процес всмоктування води. Загальна поверхня кореня, як правило, набагато перевищує зовнішню поверхню надземних органів. Завдяки швидкому рості і галуженні корені переміщуються в глибину ґрунту, захоплюючи і освоюючи його нові ділянки.
Морфолого-генетичні зони молодого кореня
Якщо прослідкувати будову молодого кореня по всій його довжині від кінчика до основи, то можна помітити, що вона на різних рівнях змінюється, як змінюються і її функції. У будові молодого кореня виділяють такі морфолого-генетичні зони: зона ембріонального росту або поділу клітин, яка покрита кореневим чохликом; зона росту і розтягування; зона кореневих волосків або зона всмоктування (зона поглинання або диференціації); зона бічних коренів або зона проведення.
Зона ембріонального росту (зона поділу клітин) кореня займає кінцеве положення. Її довжина становить приблизно 1 мм. Має жовтуватий колір, тому що клітини меристеми заповнені цитоплазмою і не мають постійних вакуолей. Складається з тонкостінних паренхіматичних клітин первинної меристеми - ініціалей. Довгий час вважали, що постійно діляться саме ініціалі. Однак, пізніше було з’ясовано, що велика кількість поділів відбувається на деякій відстані від ініціалей, які утворюють відносно неактивну зону - спочиваючий центр. Спочиваючий центр здатний відновлювати периферійні зони меристеми при їх пошкодженні.
Зовні точка росту кореня захищена кореневим чохликом.
Кореневий чохлик (calyptra) - ковпачкоподібне утворення з живих, пухких, тонкостінних паренхіматичних клітин, що вкриває молодий кінчик кореня (точку росту) і захищає його від механічних пошкоджень під час просування вглиб ґрунту. Наявність кореневого чохлика - одна із морфологічних ознак кореня як підземного органа рослини.
Довжина кореневого чохлика не перевищує 1 мм.
Паранхіматичні клітини кореневого чохлика продовгуватої форми, слабко зв’язані одна з одною. Така характеристика клітин чохлика ніби суперечить його захисній функції. Проте в основу захисної функції чохлика входить те, що його клітини постійно оновлюються, а зовнішні клітини, які доторкаються до ґрунту, відмирають і злущуються і ослизнюються. Слиз обгортає корінь і забезпечує просування ростучого кінчика кореня в ґрунті. Вважають, що слизиста речовина - це гідратований полісахарид (пектинової природи), що виділяється як секрет зовнішніми клітинами чохлика і на поверхні клітинної оболонки утворює дрібні краплини. Залежно від довжини чохлика та виду рослини, від утворення кожної клітини чохлика до її злущування проходить 4-9 днів.
Слід відмітити, що кореневий чохлик є структурою, яка сприймає дію сили тяжіння. Підтвердженням цього є наявність у центральних клітинах чохлика крохмальних зерен, які так зберігаються постійно.
Відновлення кореневого чохлика здійснюється групою клітин верхівкової меристеми, які лежать у самої основи точки росту кореня. Група ініціальних клітин, що дають початок кореневому чохлику, називається каліптрогеном.
Багато рослин не мають кореневого чохлика. Зокрема, він відсутній у рослин-паразитів (повитиця, вовчки), напівпаразитів (омела), водних рослин. Проте в ряски та інших водних рослин кінчик кореня замість чохлика має спеціальну кишеньку, яка схожа на наперсток. Існує гіпотеза, що це пристосування виникло, мабуть, для захисту коренів від вилуговування під дією води та поїдання мешканцями водойм.
Над зоною поділу розміщується зона розтягування. Зовнішню, світлу частину цієї зони називають периблемою, а центральну, темну - плеромою. З плероми утворюється центральний циліндр, а розростання і диференціація периблеми дає початок первинній корі. Поверхневий шар клітин (дерматоген) перетворюється на епіблему - одношарова поверхнева покривна тканина наступної зони кореня. Саме в зоні розтягування клітин збільшується довжина кореня. Клітини цієї зони утворюють великі вакуолі, а їх ядра займають бічне положення.
Зона поглинання розміщується за зоною розтягування. Її довжина становить 1,5-2 см. В цій зоні формується постійна тканина - епіблема, з клітин якої утворюються численні кореневі волоски. Вони мають прозенхімну форму і вбирають з ґрунту воду і розчинені в ній мінеральні речовини. Кореневі волоски виникають екзогенно і густо вкривають корінець на відстані 0,1-10,0 мм від його кінчика.
Довжина кореневих волосків варіює від 0,05 мм до 10,0 мм. У деревних рослин волоски коротші, а у трав’янистих - довші. Вони дуже швидко утворюються, проте функціонують недовго і на межі всисної та провідної зон руйнуються і відмирають. Нові кореневі волоски формуються на межі із зоною росту, тому складається враження, що всисна зона весь час переміщується і завжди знаходиться поблизу кінчика кореня.
Залежно від умов існування довжина, товщина і форма кореневих волосків, а також їх кількість дуже варіюють. Встановлено, що при зменшенні вологи, однак до певної межі, довжина та кількість кореневих волосків збільшується.
Поглинаюча поверхня коренів збільшується від 3 до 40 разів, саме завдяки кореневим волоскам.
Кореневі волоски дають опору верхівці кореня і полегшують закріплення кореневої системи, тобто виконують механічну функцію.
Слід відмітити, що не у всіх рослин на коренях утворюються кореневі волоски. Їх немає у деяких водних та прибережних рослин (сусак, латаття біле тощо). Вони слабо розвинені або зовсім відсутні на коренях мікоризних рослин (бук, береза тощо). У цих рослин поглинання води та розчинів здійснюють гіфи гриба, що покривають корінь зовні.
У зоні поглинання знаходиться зона диференціації клітин, де клітини після розтягування спеціалізуються на клітини окремих тканин у зв’язку з виконанням ними певних функцій.
Зона проведення розміщується під зоною поглинання. Ця зона займає більшу частину кореня і простягається аж до кореневої шийки. Кореневих волосків в ній немає. У зоні проведення на поверхні кореня знаходиться покривна тканина. Її ще називають зоною бічних коренів, оскільки в ній ендогенно, за рахунок первинної твірної тканини - перициклу, закладаються бічні корені. В ній сформовані провідні тканини ксилеми, по яких вода і мінеральні речовини потрапляють до стебла і листків, а до кореня надходять органічні речовини.
Типи коренів. В процесі онтогенезу корінь розвивається вже в зародку насінини, де він представлений зародковим корінцем.
За походженням виділяють такі типи коренів: головний, бічні та додаткові.
Головний корінь розвивається із зародкового корінця під час проростання насінини і росте донизу (позитивний геотропізм).
Бічні корені утворюються на головному, бічному або додатковому коренях в результаті їх галуження. Вони виникають ендогенно із перициклу, на деякій відстані від апекса в акропетальному порядку (в напрямку від основи кореня до його верхівки). Бічні корені більшості рослин формують ортостихи - правильні поздовжні ряди вздовж головного кореня, оскільки закладаються певним чином по відношенню до провідних тканини головного кореня (найчастіше - напроти груп ксилеми). Слід відмітити, що у плаунів верхівкове галуження кореня і має дихотомічний характер.
Бічні корені, що утворюються на головному корені, називаються бічними коренями першого порядку, від бічних коренів першого порядку відходять бічні корені другого порядку і т.д.
У головного кореня виявлений позитивний геотропізм і негативний фототропізм. Бічні корені другого порядку ростуть більше у горизонтальному напрямку, тобто вони поперечно (трансверзально) геотропічні, а бічні корені третього порядку ростуть в усіх напрямках, але з тяжінням до землі і не мають чітко виявленої геотропічності.
На стеблах, листках та їх метаморфозах із перициклу, паренхіми чи камбію можуть утворюватися додаткові (адвентивні) корені, яким теж властивий позитивний геотропізм. Адвентивні корені не кореневого походження, вони виникають на інших вегетативних органах. Однак, іноді можуть утворюватися на старих коренях. Закладаються вони ендогенно, але на відміну від бічних коренів не виявляють суворого акропетального порядку виникнення.
Додаткові корені обов’язково утворюються на підземних видозмінах пагона - бульбах, цибулинах, кореневищах. У однодольних рослин головний корінь рано припиняє свій ріст і вся коренева система складається із додаткових коренів. У багатьох рослин, як однодольних, так і дводольних, додаткові корені розвиваються із підсім’ядольного коліна і нижньої частини стебла. Цьому сприяє підгортання рослин ґрунтом. Додаткові корені забезпечують краще живлення та підвищують стійкість стебел. За їх участю забезпечується вегетативне розмноження. Таким чином, у більшості однодольних і дводольних рослин розвиваються додаткові корені, проте є рослини - дводольні однорічники (лобода біла, грицики), хвойні (сосна) та листопадні (дуб) дерева, у яких за нормальних умов такі корені не розвиваються взагалі.
За формою корені найчастіше циліндричні, довгі, товсті (шнуроподібні) чи тонкі (ниткоподібні), рідше - конічні, веретеновидні (морква, петрушка), конусовидні, ріповидні (ріпа, редиска), шишкуваті, бульбочковидні або іншої форми. В поперечнику корені мають округлу форму.
У рослин, які зростають в посушливих умовах, на час короткочасних дощів утворюються недовговічні ефемероїдні додаткові корені. Під час пересихання ґрунту вони гинуть.
У коренів рослин іноді спостерігається явище окорковіння. Метакутинізація - припинення росту коренів дерев чи трав, яке супроводжується окорковінням кореневих закінчень. При цьому утворюються захисні футляри, які навесні розриваються і звільняють апекси коренів.
У деяких рослин, які мешкають у більш-менш екстремальних умовах існування з сухим жарким кліматом або суворою зимою, зустрічаються скоротливі, або контрактильні корені. Найчастіше вони зустрічаються у багаторічних трав’янистих дводольних рослин (наприклад, конюшина, гречка, морква, цукровий буряк тощо). Такі корені втягують у ґрунт гіпокопиль і базальну частину пагонів з бруньками відновлення.
По відношенню до субстрату, у якому корені розміщуються і з якого всмоктують воду і мінеральні речовини, виділяють чотири типи коренів: підземні, водяні або плаваючі, повітряні та гаусторії (присоски рослин-паразитів).
Підземні - корені, які цілком або частково знаходяться у ґрунті. Такі корені мають біля 70 % рослин.
Водяні корені завжди розміщені у товщі води і ніколи не досягають дна водойми (наприклад, ряска тощо).
Повітряні корені формуються в атмосфері і ніколи не досягають ґрунту. Вони характерні для рослин-епіфітів і мешканців тропічних вологих лісів.
Гаусторії (корені-присоски) - одноклітинні або багатоклітинні утвори, за допомогою яких рослини-паразити та напівпаразити всмоктують поживні речовини з рослини-хазяїна (наприклад, повитиця, омела).
Типи кореневих систем.
Кореневою системою називають сукупність усіх коренів рослини. Тип кореневої системи є спадково закріпленою ознакою.
Виділяють три типи кореневих систем рослин за морфологічною будовою: система головного кореня або стрижнева, система додаткових коренів або мичкувата, змішана коренева система.
Стрижнева коренева система - це система, в якої головний корінь добре виділяється серед маси інших коренів за довжиною і товщиною та потужним розвитком. Вона утворена системою головного кореня. Стрижнева коренева система притаманна більшості дводольним і голонасінним рослинам. Слід відмітити, що у деяких дводольних рослин (конюшина, перстач) додаткові корені на своїх горизонтальних пагонах потовщуються, галузяться і здатні утворювати вторинно-стрижневу кореневу систему.
Мичкувата коренева система - це система, в якої головний корінь швидко відмирає або слаборозвинений і не виділяється своїм розвитком і потужністю серед великої кількості додаткових коренів, які сформувалися на нижній частині стебла. Вона утворена системою додаткових та бічних коренів. Мичкувата коренева система характерна зазвичай однодольним рослинам, проте у деяких дводольних рослин також зустрічається такий тип кореневої системи (наприклад, у жовтецевих та подорожникових).
У деяких рослин виділяють ще змішану кореневу систему. Така коренева система має добре розвинений головний корінь з численними бічними та додатковими коренями, що розвиваються на нижній частині стебла. Змішана коренева система властива деяким дводольним рослинам (наприклад, помідор, капуста, перець, огірки, квасоля, смородина тощо).
За походженням виділяють кілька типів кореневих систем: первинно-гоморизна, вторинно-гоморизна, алоризна.
У первинно-гоморизної кореневої системи головний корінь не утворюється, а формуються виключно додаткові корені. Це генетично первинна, найбільш примітивна система, яка характерна для плаунів, хвощів, папоротей. Оскільки ці рослини не мають насіння, а отже і зародкового корінця, вся їх коренева система утворена тільки додатковими коренями.
Вторинно-гоморизна коренева система також утворена тільки додатковими коренями, однак на відмінну від первинно-гоморизної кореневої системи, головний корінь у проростка утворюється, проте рано відмирає або взагалі не розвивається. Така коренева система характерна для однодольних (наприклад, злаки)та дводольних рослин, які розмножуються вегетативним шляхом (наприклад, картопля, суниці, матий мачуха).
Алоризна коренева система утворена головним, бічними і додатковими коренями. Така коренева система характерна для дводольних рослин (наприклад, помідор, капуста тощо).
На характер росту кореневої системи впливає тип ґрунту, його механічний склад, ступінь зволоження, вміст поживних речовин тощо. За напрямком росту розрізняють три типи кореневої системи: поверхневу горизонтальну, глибинну вертикальну, універсальну. Формування цих кореневих систем є відображенням еколого-фізіологічного пристосування рослин до умов ґрунтового забезпечення.
При поверхнево горизонтальному типу кореневої системи, головний корінь відмирає, а бічні і додаткові ростуть паралельно поверхні ґрунту (наприклад, ялина, опунція, тюльпан тощо).
При глибинно вертикальному типу кореневої системи, всі корені рослини ростуть вглиб ґрунту (наприклад, верблюжа колючка, саксаул тощо).
При універсальному типу кореневої системи, корені рівномірно розподіляються по горизонтах ґрунту, тобто розростаються у всі боки.
У межах кореневої системи однієї рослини (особливо багаторічної) гетероризація - морфологічна і функціональна диференціація коренів.
У листяних та плодових дерев утворюються скелетні та напівскелетні корені, на яких виникають недовговічні обростаючі кореневі мички. Кореневі мички формуються близько до поверхні ґрунту, що пов’язано із сезонними зволоженнями чи поливом.
Скелетні корені І-го порядку та напівскелетні корені ІІ-го та ІІІ-го порядків видовжені, довговічні, мають вторинну будову і виконують провідну функцію. Корені IV-го та наступних порядків - обростаючі кореневі мички. Частина з них - ростові корені - видовжені, довговічні, мають вторинну будову, сприяють розширенню площі живлення. На їх верхівках утворюються бічні вкорочені всмоктувальні корені, які мають первинну будову, поглинають поживні речовини, наростають обмежено, швидко відмирають і замінюються новими. Перехідні кореневі мички можуть мати первинну чи вторинну будову.
Значення кореневих систем полягає в тому, що вони подрібнюють субстрат, збільшують площу мінерального живлення, змінюють його хімічний склад, збагачують ґрунт органічними речовинами після свого відмирання, закріплюють ґрунти від ерозії тощо.
Видозміна і спеціалізація коренів
Нормальне середовище існування типових коренів - ґрунт. Умови існування у ньому відносно однотипні, тому і порівняно мала різновидність у будові коренів. Однак, якщо змінюються умови існування, корені проявляють високу пластичність. Вони виконують інші функції і набувають своєрідної будови. Зміна форми й будови органів рослин, які виникають в процесі історичного розвитку (філогенезу) в зв’язку із зміною функцій або умов функціонування і які передаються по спадковості називають метаморфозом. Вчення про метаморфоз сформулював Гете.
Метаморфоз кореня може бути пов’язаний з явищем симбіозу і паразитизму, з розвитком кореня у воді або повітряному середовищі. Метаморфізовані корені можуть виконувати опорну, дихальну, асимілюючу, запасаючу, симбіотичну, живильну та інші функції.
До метаморфозів кореня, які виконують функції запасу та вегетативного розмноження, належать коренеплоди, коренебульби і стеблокоренеплоди. Вони потовщені, у зв’язку із значним розвитком паренхімної тканини. У запасаючій паренхімі первинної кори, деревини або серцевини відкладаються такі запасаючі поживні речовини, як вуглеводи, особливо крохмаль і цукри.
Коренеплоди і стеблокоренеплоди складаються з трьох частин різного походження і форми: головки, шийки і власне кореня. Головка - вкорочений епікотиль (надсім’ядольне коліно), яке несе прикореневу розетку листків і пазушні бруньки. Шийка, або гіпокотиль (підсім’ядольне коліно) розміщується під головкою. На шийці можуть відростати додаткові корені. Власне корінь - нижня частина коренеплоду. У різних рослин кожна із складових частин коренеплоду вкорочується або потовщується в більшій чи меншій мірі і набуває своєрідної форми. Це залежить від того, яка частина найбільш інтенсивно розростається та яка тканина (флоема чи ксилема) резервує поживні речовини.
Кореневі шишки, або коренебульби - потовщення бічних чи додаткових коренів, а іноді й певної частини стебла (наприклад, жоржини, любка, батат) або стеблових бруньок (зозулинці, пшінка). Досить часто на коренебульбах закладаються додаткові бруньки, що забезпечує рослині вегетативне розмноження.
Часто додаткові корені виконують не одну, а декілька функцій. Наприклад, опорні корені поєднують опорну та живильну функції і утворюються у різних рослин певним чином. Так, ходульнікорені розвиваються на нижніх вузлах трав’янистого стебла (наприклад, у кукурудзи) або ростуть косо вниз від стовбура і гілок дерев, підтримуючи велику крону (наприклад, у тропічних мангрових дерев, пандануси, деякі пальми). Опорні додаткові корені, що розвиваються на надземних органах звисають у повітрі, називаються повітряними. Вони можуть бути тонкими (наприклад, монстера, філодендрон, епіфітні орхідеї) або досягати значної товщини (наприклад, стовпоподібні корені індійського баньяна або якірні корені сосни). Повітряні корені покриває особлива багатошарова всисна тканина, яка утворюється з протодерми - веламен.
Веламен вбирає вологу з повітря капілярним шляхом, захищає кору і навіть здатний фотосинтезувати.
До опорних коренів належать і бічні дошкоподібні корені, які відростають від основи стовбура косо в грунт у вигляді плескатих 1-3-метрових завширшки перегородок (наприклад, в’яз, тополя, бук та деякі дерева вологих тропічних лісів). Дошкоподібні опорні корені характерні для дерев найвищого ярусу тропічного дощового лісу. Це бічні корені, а не додаткові.
Дихальні корені, або пневматофори - корені деяких болотних і водних рослин, які піднімаються своїми верхівками над поверхнею ґрунту або води, забезпечуючи підземні органи киснем (наприклад, у кипариса болотяного, авіценії тощо). За походженням це додаткові корені. Крізь сочевички, які розташовані на них, повітря надходить до повітряних порожнин кореня. Дихальні корені мають добре розвинену аеренхіму, між клітинами якої знаходяться ці повітряні порожнини. Вони завжди заповнені повітрям, яке необхідне для дихання занурених у воду частин рослин. Близько до поверхні ґрунту на дихальних коренях утворюються тонкі корені, саме вони є основною поглинаючою частиною кореневої системи багатьох мангрових рослин. Дихальні корені іноді мають отвори на кінчиках коренів - дихальні пори, через які здійснюється аерація.
Втягуючі або контрактильні (скоротливі) корені - бокові або додаткові корені, які у процесі розвитку здатні скорочуватися у поздовжньому напрямку, завдяки чому надземна частина рослини, тобто розетка, втягується у ґрунт. Такі корені характерні для багаторічних трав’янистих рослин, наприклад, деяких видів амарилісових, лілійних тощо. Поздовжні скорочення контрактильних коренів здійснюється завдяки певним анатомічним та фізіологічним особливостям. Це здійснює заглиблення в ґрунт цибулин, кореневищ, бульб або притиснення до ґрунту листкових розеток, що цим самим захищає зимуючі підземні органи від різних температурних коливань і сприяє формуванню додаткових коренів.
Чіпкі ліани, а також деякі лазячі рослини (наприклад, іпомеї, фікуси, плющі, дикий виноград тощо) мають додаткові корені-причіпки. Такі корені присмоктуються до поверхні стовбурів, стін будівель, скель тощо, цим самим піднімають рослину і закріплюють на них. Корені-причіпки властиві рослинам, що мають слабке, витке стебло. Дрібні додаткові корені формуються на нижній стороні стебла, проникають у тріщини опори і утримують стебло з листками на вертикальних опорах.
Зеленуваті асиміляційні корені утворюються у деяких епіфітів, у окремих гідрофітів (наприклад, водяний горіх) та напівпаразитів (наприклад, омела). Асиміляційні корені - корені, в тканинах яких міститься хлорофіл і які виконують функцію фотосинтезу. У епіфіт них орхідей такі корені плоскі, стрічковидні. Їх нижня сторона вкрита волосками і поглинає розчини, а верхня - фотосинтезує. Слід відмітити, що асиміляційні корені закладаються на підсім’ядольному коліні і на відміну від звичайних коренів мають вигляд розсічених утворів.
Корені-присоски або гаусторії - це видозмінені корені, які розвиваються у тканинах вищої рослини-господаря і висмоктують з них поживні речовини. Такі метаморфози кореня характерні рослинам паразитичним (наприклад, петрів хрест, вовчок, повитиця) і напівпаразитичним (наприклад, омела, дзвінець, філодендрон, перестріч). Гаусторії на кінцях цих коренів переходять у присоски - довгі волосковидні ряди клітин, які проникають до деревини або до судин пучка рослини-господаря.
Кореневі паростки. У деяких рослин на коренях утворюються додаткові бруньки, які здатні утворювати надземні пагони - кореневі паростки. Вони властиві таким рослинам, як бузок, малина, смородина, слива, вишня, осот польовий, осот жовтий, суріпиця звичайна, барбарис, осика, берізка польова тощо. Такі рослини називають коренепаростковими. Ці пагони служать для вегетативного розмноження. Кореневі паростки особливо характерні для бур’янів, які швидко розмножуються.
Мікориза. Зміна зовнішньої і внутрішньої будови може бути викликана симбіозом. Симбіоз — співжиття організмів різних видів, з якого вони мають взаємну вигоду. Для коренів вищих рослин характерний симбіоз з грибами і бактеріями. Мікориза (грибокорінь) - співжиття міцелію гриба з коренями деяких вищих рослин. Мікоризу утворюють гриби з класів зигоміцети, аскоміцети та базидіоміцети. Відповідно до глибини проникнення гіфів гриба до тканин кореня розрізняють ектотрофну, ендотрофну і екто-ендотрофну мікоризи.
Гриби - це гетеротрофні організми, тобто вони живляться готовими органічними речовинами, всмоктуючи їх всією поверхнею тіла із зовнішнього середовища. Тіло гриба - грибниця складається з тонких довгих переплетених ниток - гіфів. У випадку ектотрофної мікоризи гіфи гриба обплітають кінчики молодих бічних коренів. На коренях вони утворюють щільний футляр або чохол, який обволікає кореневі закінчення. Будова таких коренів спрощується: кореневий чохлик відсутній або дуже слабо розвинений, не розвиваються кореневі волоски, не відбувається вторинний ріст і в зв’язку з цим зберігається первинна кора. Грибниця, яка розвивається на коренях, стимулює процес галуження. Ектотрофна мікориза характерна для багатьох дерев та кущів. Наприклад, для сосни, берези, осики, клена, липи, ліщини тощо. Такі мікоризи утворюють гриби з класу базидіоміцети (гіменоміцети та гастероміцети) і деякі аскоміцети.
При ендотрофній мікоризі грибниця розвивається всередині клітин первинної кори і забезпечує поглинання поживних речовин. Кореневі волоски при цьому не відмирають. Така мікориза характерна для орхідей, цибулі, півників, конюшини, суниць, буркуна, яблуні, груші, томатів тощо. В утворенні цієї мікоризи беруть участь гриби з класів ооміцети та зигоміцети. Найбільш високо спеціалізованою формою ендотрофної мікоризи є мікориза зозуленцевих.
Зустрічається мікориза змішаного типу - екто-ендотрофна. Такий тип характерний для вільхи, дуба. Гіфи грибів як частково проникають в клітини корової паренхіми, так і можуть знаходитися на поверхні кореня.
При мікоризі між грибами і вищими рослинами виникають складні біологічні відносини. Гриби знаходять на коренях і в коренях сприятливе середовище для розвитку і джерело органічного живлення. В свою чергу для вищих рослин ектотрофна мікориза виконує роль кореневих волосків.
За допомогою грибних ниток рослина всмоктує з ґрунту поживні речовини і в першу чергу азотисті сполуки. У випадку ендотрофної мікоризи грибниця бере участь, у первинній переробці деяких речовин, які надходять з ґрунту.
Слід відмітити, що мікоризні шапкові гриби (наприклад, боровики, дубовики, підберезовики, грузді, рижики тощо) не можуть рости поза кореневою системою дерев, але й дерева здебільшого гинуть, якщо в них на коренях не утворюється мікориза.
Рослини, які одержують поживні речовини за участю міцелію гриба, називають мікотрофними (мікотрофи). Розрізняють облігатні та факультативні мікотрофи. Облігатними мікотрофними рослинами називають вищі рослини, які нормально розвиваються лише при зараженні грибом-симбіонтом. Наприклад, види родини зозуленцевих, дуб, бук, сосна, граб, ялина тощо. Факультативні мікотрофні рослини - рослини, які розвиваються при зараженні грибом-симбіонтом, але можуть нормально жити без нього.
Бактеріориза — співжиття коренів вищих рослин з бактеріями, які зв’язують атмосферний азот і переводять його у розчинні сполуки, збагачуючи при цьому ґрунт. Між бактеріями та клітинами кореня існує тісна взаємодія, внаслідок якої у вищих рослин синтезуються органічні речовини, такі як вітаміни, ферменти тощо, а бактерії використовують органічні речовини кореня.
Симбіоз кореня з бактеріями характерний для рослин родини бобових. Бактерії проникають у кору кореня з ґрунту, розмножуються і викликають посилене розмноження клітин кори. В результаті такого інтенсивного розростання кори на поверхні кореня утворюється клубочок. Всередині нього знаходиться бактеріальне гніздо. До цього гнізда зі сторони центрального циліндру кореня підходять розгалуження провідних пуків, що забезпечує надходження до нього поживних речовин і води. Отже, бульбочка складається з бактеріального гнізда, зв’язаного провідними тканинами з флоемою і ксилемою кореня. Скупчення азотфіксуючих бактерій на коренях вищих рослин характерні для бобових, розоцвітих, гінкгових, березових, хрестоцвітих, жостерових, маслинкових та злаків.
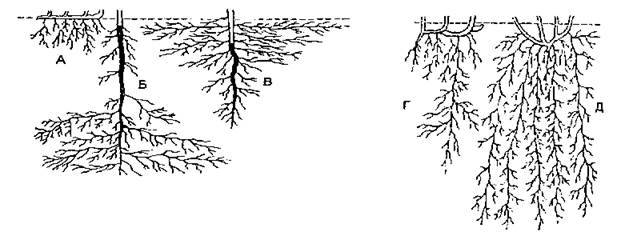
Рис. 1. Кореневі системи: А - первинно-гоморизна; Б — Г - алоризні; Д - вторинно-гоморизна; Б - В - стрижневі; А,Г,Д - мичкуваті; Б - глибинна; А,В - поверхневі; Д - універсальна. Головний корінь затемнений.