МОРФОЛОГИЯ РАСТЕНИЙ - Т. А. Сауткина - 2012
ГЛАВА 5. ВЕГЕТАТИВНЫЕ ОРГАНЫ РАСТЕНИЙ
Для подавляющего большинства высших растений характерно разделение тела на органы. Орган — это часть тела растения, которая имеет индвидуальное строение, занимает определенное место в теле растения и выполняет характерную функцию.
Органы, которые поддерживают индивидуальную жизнь растений, называются вегетативными (от лат. vegeto — оживляю, возбуждаю, в дальнейшем — расту). К вегетативным органам традиционно относят корень, стебель и лист, однако некоторые ботаники считают, что у растений существует два вегетативных органа — корень и побег (стебель с листьями). Побег как вегетативный орган выделяется на том основании, что формирование стебля и листа происходит в процессе индивидуального развития растения одновременно из верхушечной меристемы конуса нарастания стебля. В то же время по своим морфологическим, анатомическим и функциональным особенностям стебель и лист существенно различаются, что дает основания рассматривать их как самостоятельные органы.
Помимо вегетативных, у растений существуют репродуктивные (от лат. re — приставка, обозначающая возобновление, повторение и produco — произвожу, создаю) органы, или органы размножения. К репродуктивным органам относятся спорангии (органы, в которых образуются споры) и гаметангии (органы, в которых образуются гаметы, или половые клетки). Гаметангии высших растений подразделяются на мужские (антеридии) и женские (архегонии). Так как в гаметангиях образуются половые клетки, эти органы также называют половыми, или генеративными (от лат. genero — рождаю). Часто термины «репродуктивные» и «генеративные» органы употребляют как синонимы. Хотя внешне репродуктивные и вегетативные органы различаются, в функциональном отношении они могут быть сходными. При определенных условиях вегетативные органы могут выполнять функцию размножения растений.
5.1. Анатомо-морфологические функциональные особенности корня
Корень — это вегетативный орган, обладающий положительным геотропизмом (растет по направлению силы тяжести Земли), способный относительно неограниченно нарастать в длину и не несущий листьев, почек и цветков. Так определяют понятие «корень» исходя из его морфологических особенностей. Однако это определение нельзя считать абсолютно безупречным. Есть растения, у которых корни растут отрицательно геотропично (вверх, против силы тяжести Земли), такие корни характерны для растений мангровых зарослей. У некоторых, так называемых корнеотпрысковых растений, на корнях формируются придаточные почки, но никогда на корнях не образуются ни листья, ни цветки.
Как вегетативный орган корень выполняет три основные функции: удержание растения в субстрате (механическая функция), всасывание воды с растворенными в ней минеральными веществами (функция почвенного питания), выделение некоторых веществ в окружающую среду. Корни, которые выполняют типичные функции, имеют бечевковидную форму и обладают радиальной симметрией. Кроме основных, корень может выполнять ряд дополнительных функций: синтез некоторых органических веществ, накопление запасных питательных веществ, функцию фотосинтеза и др. У корней, выполняющих дополнительные функции, меняется не только внешний вид (они становятся реповидными, клубневидными, досковидными), но и внутреннее строение.
В процессе эволюции, по мнению ряда исследователей, корень мог возникнуть из ризомоидов первопоселенцев суши или в результате дифференцировки углубившихся в почву мезомов. Из современных высших растений не имеют корней только мохообразные и псилотообразные. Эти растения являются первично бескорневыми. Среди некоторых представителей других таксонов высших растений также есть виды, не имеющие корней. Исчез корень у ряда водных растений (сальвиния плавающая, роголистник — Ceratophyllum и др.), а также у растений-паразитов. Эти растения являются вторично бескорневыми. Корень у них исчез под влиянием условий существования в связи с потерей основных функций. Под действием экологических факторов могут редуцироваться и такие важные компоненты корня, как корневые волоски и корневой чехлик.
5.1.1. Морфологическая дифференцировка корня
Развитие корня в онтогенезе покрытосеменных растений начинается с формирования зародыша семени. При дифференцировке зародыша на ранних стадиях онтогенеза закладывается конус нарастания корня. Он расположен на стороне, противоположной конусу нарастания стебля. По мере развития зародыша семени в нем оформляются все зародышевые органы, в том числе и зародышевый корешок (лат. — radícula). У двудольных растений зародышевый корешок возникает экзогенно, т. е. из поверхностных слоев меристематических тканей. У некоторых однодольных, например, у злаков, зародышевый корешок образуется эндогенно (от греч. endo — внутри) и окружен чехлом поверхностной ткани — колеоризой, или корневым влагалищем.
При прорастании семени зародышевый корешок первым трогается в рост. Корень, который образуется из зародышевого корешка семени, называется главным. Главный корень, как правило, растет вертикально вниз. На молодом растущем корешке обнаруживается более или менее ясно выраженная морфологическая вертикальная зональность: зоны роста,всасывания и проведения, за которыми постепенно формируется зона ветвления. Верхушка корешка (апекс корня, апикальная зона), образованная меристематическими клетками, составляет зону роста. В этой зоне происходит активное деление меристематических клеток, рост образовавшихся клеток (растяжение) и начинается их дифференцировка, обеспечивающая формирование постоянных тканей. Таким образом, однородная по внешнему виду зона роста в анатомическом отношении оказывается состоящей из трех функционально различающихся участков: зоны деления, зоны растяжения и зоны дифференцировки.
Верхушка корешка прикрыта корневым чехликом (лат. — calyptra). Корневой чехлик выполняет защитную функцию, предохраняя верхушку растущего корня от повреждений при прохождении через почву, а также облегчает продвижение корня в субстрате и способствует переводу некоторых минеральных веществ в форму, доступную для растений. Выполнение этих функций обеспечивается благодаря строению клеток корневого чехлика, который образован живыми паренхимными клетками, содержащими большое количество крахмала. Есть данные, что, благодаря высокому содержанию в клетках крахмала, корневой чехлик играет гравитационную роль, т. е. контролирует направление роста корня. По мере роста корня периферические клетки корневого чехлика слущиваются и ослизняются. Слизь на поверхности отделившихся клеток корневого чехлика представляет собой высокогидратированный полисахарид, по-видимому пектиновые вещества. Согласно имеющимся данным, процесс ослизнения клеток корневого чехлика связан с функцией диктиосом, которые формируют крупные пектиновые пузырьки. Пектиновые пузырьки мигрируют в периферические части клетки, сливаются с плазмалеммой, расположенной вдоль наружных стенок клеток, а содержащаяся в них слизь накапливается между плазмалеммой и оболочкой клетки. Отсюда она постепенно выходит через оболочку на ее наружную поверхность. Слизь защищает кончик корня от высыхания, склеивает частицы почвы, в результате чего образуется своеобразный канал, по которому растет корень. Возможно, она в какой-то мере играет также роль абсорбирующей поверхности и принимает участие в процессах метаболизма. По мере слущивания периферических клеток корневого чехлика в его центральной части образуются новые клетки. Корневой чехлик отсутствует на корнях водных растений, в то же время у некоторых из них верхушка корня защищена крупным корневым кармашком.
За морфологической зоной роста располагается зона всасывания. Характерной особенностью этой зоны является наличие на поверхности корня корневых волосков, которые формируются как боковые выросты покровной ткани. Первичная покровная ткань в зоне всасывания — эпиблема — состоит из двух типов клеток: трихобластов, образующих корневые волоски, и атрихобластов, не способных к этой функции. У большинства растений корневые волоски не отделены от материнской клетки перегородкой и представляют собой одноклеточные структуры. Однако у некоторых покрытосеменных корневые волоски многоклеточные (ананас — Ananas sativus, марьянник луговой — Melampyrum pratense).Оболочка корневого волоска тонкая, имеет первичное строение и состоит из целлюлозы и пектиновых веществ. Благодаря наличию пектиновых веществ на поверхности клеточных волосков образуется слизь, которая склеивает частички почвы, а также, по-видимому, способствует избирательному поглощению ионов. Длина корневых волосков варьирует от 0,05 мм (у цитрусовых) до 10 мм (у сахарной свеклы). Число корневых волосков на 1 мм2 поверхности у растений различно (у проростков гороха более 200, у кукурузы более 400). Большое количество корневых волосков образуется у растений, растущих на хорошо увлажненной почве, однако у водных растений корневых волосков очень мало или их вообще нет. Подавлено развитие корневых волосков и у некоторых растений степей (шафран — Crocus, тюльпан — Tulipa).
Корневые волоски значительно увеличивают всасывающую поверхность корня. Они развиваются очень быстро, но недолговечны и через несколько дней отмирают, а вместо них формируются новые. Отмирание и развитие корневых волосков идет в акропетальном направлении (от основания к верхушке). В зоне всасывания осуществляется и проведение питательных веществ (рис. 61).
Рис. 61. Схема строения кончика корня: 1—корневой чехлик; 2—группа инициальных клеток; 3—калиптроген; 4—зона деления клеток; 5—зона роста клеток; 6—зона всасывания; 7—зона ветвления (проведения); 8—образующиеся боковые корни
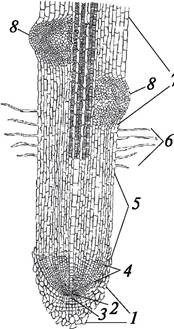
За зоной всасывания лежит зона ветвления, она характерна для более старого корня. По мере роста корня его зоны смещаются в акропетальном направлении.
Наличие зональности характерно не только для главного корня, но и для всех молодых корней растения.
5.1.2. Первичное анатомическое строение корня
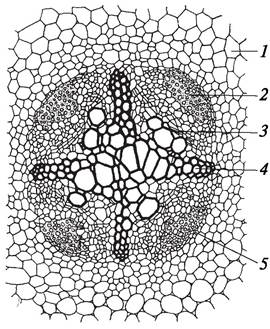
Одновременно с формированием морфологической структуры корня складывается и его первичная анатомическая структура. Как известно, верхушечная меристема конуса нарастания корня дифференцирована на дерматоген, периблему и плерому (см. рис. 13). Каждый из этих слоев имеет свои инициальные клетки, следовательно, у двудольных и однодольных растений инициальные клетки на конусе нарастания корня расположены в три этажа. Принципиально важным различием у двудольных и однодольных растений является функционирование инициальных клеток, занимающих разное положение. У однодольных из самой нижней группы инициальных клеток — калиптрогена — образуется корневой чехлик. Первичная покровная ткань (ризодерма, иногда употребляют термин «эпиблема» как синоним термина «ризодерма») и первичная кора возникают из среднего слоя (периблемы), а центральный цилиндр — из внутреннего (плеромы). У двудольных растений инициальные клетки нижнего этажа (дерматогена) делятся периклинально (параллельно поверхности органа) и дают начало корневому чехлику и первичной покровной ткани, поэтому этот слой называют дерматокалипрогеном. Внутренние клетки дерматокалипрогена делятся только в одном направлении — антиклинально (перпендикулярно поверхности органа) и образуют первичную покровную ткань корня — ризодерму. Меристематические клетки периблемы формируют первичную кору. Из клеток плеромы образуется центральный цилиндр. Таким образом, в молодом корешке четко выделяются две топографические анатомические зоны: первичная кора, расположенная под ризодермой, и центральный цилиндр (рис. 62).
Рис. 62. Поперечный срез корня ириса (Iris germanica): 1—ризодерма; 2—экзодерма; 3—основная паренхима (мезодерма); 4—эндодерма; 5—пропускная клетка эндодермы; 6—перицикл; 7—луч первичной ксилемы; 8—участок первичной флоэмы (2—5 — первичная кора; 6—8 — центральный цилиндр)
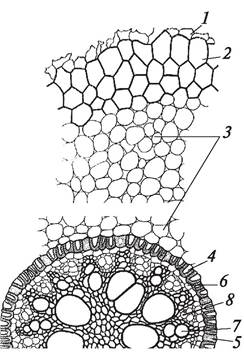
Первичная кора состоит из паренхимных клеток, которые отличаются размерами, строением клеточных оболочек и выполняют различные функции. Первичная кора дифференцирована на три слоя: экзодерму, мезодерму и эндодерму.
Экзодерма (от греч. exo — вне, снаружи) расположена непосредственно под ризодермой. Клетки ее имеют утолщенные клеточные оболочки. Экзодерма состоит из нескольких слоев клеток, имеющих многоугольные очертания и плотно прилегающих одна к другой. На ранних этапах развития корешка экзодерма выполняет механическую функцию, а также защищает корень от излишней потери воды, от проникновения в него фитопатогенных микроорганизмов. У некоторых растений оболочки клеток экзодермы могут пропитываться суберином и опробковевать. Это приводит впоследствии к отмиранию и слущиванию ризодермы, и тогда экзодерма становится покровной тканью. Особенно хорошо развита экзодерма у однодольных растений, у водных растений она отсутствует.
Мезодерма (от греч. mes — средний) состоит из различных по размеру и форме паренхимных клеток, между которыми образуются хорошо заметные межклетники. Наличие межклетников обеспечивает газообмен в корне. У водных и водно-болотных растений мезодерма представлена типичной аэренхимой. По паренхиме первичной коры к центральному цилиндру поступает из внешней среды вода с растворенными в ней минеральными веществами. Растворы питательных веществ, диффундирующие в корневой волосок, а также в периферические ткани первичной коры корня, передвигаются в горизонтальном направлении как по симпласту (через клетки по плазмодесмам), так и по апопласту (по межфибриллярным пространствам оболочек клеток и межклетникам).
Эндодерма (от греч. endo — внутри) — самый внутренний слой первичной коры. Она состоит из одного ряда клеток и прилегает непосредственно
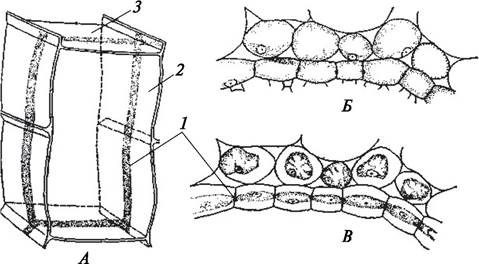
к центральному цилиндру. На ранних этапах онтогенеза клетки эндодермы живые, тонкостенные, вытянутые вдоль продольной оси корня. На поперечном разрезе они имеют правильную четырехугольную форму. В процессе развития первичной коры эндодерма проходит три стадии. На первой стадии в центральной зоне первичной оболочки клеток эндодермы на радиальных стенках появляются утолщения — пятна Каспари, названные по имени итальянского ботаника (J. X. R. Caspary, 1818—1887), описавшего эти образования. В состав утолщений входят целлюлоза и суберин. На второй стадии утолщение клеточной оболочки продолжается и распространяется на экваториальную часть радиальных и тангентальных стенок. Так в клетках эндодермы образуются пояски Каспари. На внутренней стороне этих утолщений откладывается тонкий слой суберина. Пояски соседних клеток вплотную прилегают друг к другу. Образование поясков Каспари наблюдается к моменту завершения формирования первичной коры корня. На третьей стадии развития, которая наиболее типично проявляется у однодольных растений, утолщения целиком охватывают радиальные и внутреннюю тангентальную стенки клеток. Поверх субериновой прослойки откладывается слой целлюлозы, который впоследствии пропитывается лигнином и одревесневает. Содержимое клеток отмирает, а первичная оболочка сохраняется только на наружной тангентальной стенке. Через эти оболочки (по апопласту) не могут передвигаться водные растворы минеральных веществ, поступающие из внешней среды к центральному цилиндру и из него к периферическим слоям первичной коры. Таким образом, эндодерма играет роль своеобразного физиологического барьера, регулирующего ток воды и веществ из первичной коры в центральный цилиндр и обратно. Она препятствует свободному перемещению ионов. Однако не все клетки эндодермы претерпевают вторичные и третичные изменения, некоторые клетки сохраняют первичное строение. Такие клетки расположены против элементов ксилемы и называются пропускными. Через них осуществляется передвижение растворов. Кроме барьерной функции эндодерма у некоторых растений (однодольных) выполняет и механическую функцию (рис. 63).
Рис. 63. Строение эндодермы: А—схематическое изображение клетки эндодермы с пояском Каспари на поперечных (тангентальных) и продольных (радиальных) стенках; Б—пояски Каспари на радиальных стенках; В—клетки эндодермы в состоянии плазмолиза (цитоплазма не отходит от оболочки вдоль всего пояска Каспари): 1—поясок Каспари; 2—радиальная стенка; 3—тангентальная стенка
Непосредственно к эндодерме примыкает перицикл — особая меристематическая ткань, которая ограничивает центральный цилиндр. Перицикл образуется из периферического слоя плеромы, следовательно, он является первичной образовательной тканью. Перицикл бывает однорядным или состоит из нескольких рядов клеток. Клетки его тонкостенные, слегка вытянутые. Перицикл длительное время сохраняет меристематические свойства и выполняет различные функции. При переходе корня от первичного анатомического строения к вторичному в перицикле закладываются камбий, феллоген. В перицикле также закладываются боковые корни, поэтому часто его называют корнеродным слоем.
Под перициклом некоторые клетки плеромы делятся вдоль и образуют прокамбиальные тяжи, которые почти вплотную подходят к зоне деления (рис. 64). Этот прокамбий дает начало первичной флоэме. Сначала под перициклом образуется протофлоэма, ситовидные трубки которой лишены клеток-спутниц. Метафлоэма возникает несколько позже и расположена ближе к центру корня.
Рис. 64. Схема формирования первичной проводящей системы в центральном цилиндре корня: 1—перицикл; 2—прокамбий, образующий первичную флоэму; 3— прокамбий, образующий первичную ксилему; 4—протофлоэма; 5—метафлоэма; 6—протоксилема; 7—метаксилема; 8—радиальный (тетрархный) сосудисто-волокнистый
пучок в молодом корне
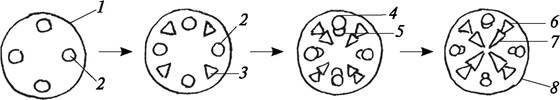
Таким образом, первичная флоэма закладывается экзархно и развивается центростремительно. Более раннее заложение первичной флоэмы имеет большое биологическое значение. Первичная флоэма подходит непосредственно к зоне деления. Именно по элементам флоэмы к верхушке корня поступают питательные вещества, которые необходимы для деятельности верхушечной меристемы.
Приблизительно на уровне зоны всасывания формируются под перициклом прокамбиальные тяжи, из которых образуется первичная ксилема. Ближе к перициклу возникает протоксилема, в состав которой входят кольчатые и спиральные элементы. За протоксилемой по направлению к центру корня образуется метаксилема. Она представлена лестничными и пористыми проводящими элементами. Таким образом, первичная ксилема, как и первичная флоэма, возникает экзархно и развивается центростремительно. Элементы первичной флоэмы и первичной ксилемы не соприкасаются друг с другом.
Они расположены по радиусам корня и разделены небольшими участками паренхимной ткани. Следовательно, в молодом корне возникает сосудистоволокнистый пучок радиального типа. В зависимости от числа ксилемных тяжей пучки могут быть диархными (с двумя участками ксилемы), триархными (с тремя участками) и т. д. Если число ксилемных тяжей более пяти, сосудисто-волокнистый пучок называется полиархным. Хотя первичная ксилема закладывается позже первичной флоэмы, она развивается быстрее и достигает центра корня, образуя звездообразную фигуру с различным числом лучей. В типичном случае в центре корня нет сердцевины из паренхимной ткани, однако в редких случаях она может формироваться (например, в корнях кукурузы). Расположение протоксилемы в непосредственной близости к перициклу имеет приспособительное значение. Вода с растворенными в ней минеральными веществами, поступающая из почвы в корень, быстрее попадает в водопроводящие элементы и транспортируется в вертикальном направлении к надземным частям растения.
У большинства однодольных растений корень в течение всей жизни сохраняет первичное анатомическое строение. Исключение составляют древесные растения (пальмы, юкки, драцены), однако у них усложнение строения корня происходит весьма своеобразно, не так, как у двудольных.
5.1.3. Вторичное анатомическое строение корня
Переход от первичного анатомического строения корня ко вторичному у двудольных растений происходит благодаря появлению камбия. Первоначально камбий закладывается в виде обособленных участков (рис. 65).
Рис. 65. Начальный этап перехода от первичного анатомического строения корня бобов (Faba vulgaris) к вторичному: 1—паренхима первичной коры; 2—первичная флоэма; 3—камбий; 4—первичная ксилема; 5—перицикл
Первые элементы камбия возникают под метафлоэмой из паренхимной ткани, у которой восстанавливается меристематическая активность. Новые элементы камбия закладываются в перицикле над протоксилемой.
Таким образом, на первых этапах развития участки камбия, имеющие разное происхождение, разобщены. Постепенно между ними из паренхимной ткани возникают камбиальные клетки и образуется сплошной слой камбия. Сначала он имеет извилистую форму, но по мере образования элементов постоянных тканей, возникающих из камбия, принимает форму окружности.
Из разных по происхождению участков камбия образуются различные постоянные ткани. Камбий, который образовался под метафлоэмой из паренхимной ткани, начинает откладывать элементы вторичных проводящих тканей. Центростремительно формируется вторичная ксилема, а центробежно — вторичная флоэма. Таким образом, в корне меняется тип сосудисто-волокнистого пучка, вместо пучка радиального типа появляются пучки коллатерального типа. Число развивающихся коллатеральных пучков равно числу тяжей первичной ксилемы, которые были сформированы в молодом корешке. Под действием разрастающихся коллатеральных пучков первичная флоэма оттесняется к перициклу и постепенно разрушается.
Камбий, начало которому дал перицикл, производит только клетки паренхимы. Они смещают первичную ксилему к центру корня, при этом протоксилема разрушается, а метаксилема образует в центральной части корня характерную фигуру из двух, четырех или более тяжей. От них, постепенно расширяясь, к периферии корня отходят тяжи горизонтальной паренхимы — первичные радиальные паренхимные лучи. По ним осуществляется передвижение веществ из проводящих тканей к живым периферическим тканям корня, а также осуществляется газообмен.
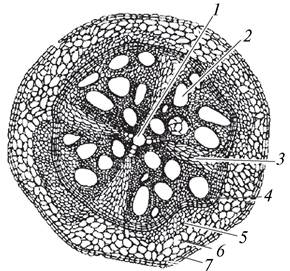
Вторичные изменения происходят не только в проводящей системе центрального цилиндра, но и в других структурах. Перицикл, кроме камбия, образует феллоген — пробковый камбий, в результате чего на корне появляется вторичная покровная ткань — перидерма. В перидерме корня у большинства растений чечевичек нет, что приводит к нарушению процессов газообмена растения с окружающей средой. Образование перидермы нарушает также транспорт растворов минеральных солей и ассимилятов. В результате этого вся первичная кора отмирает и сбрасывается. Защитную функцию в корне начинает выполнять феллема перидермы (рис. 66).
Рис. 66. Вторичное анатомическое строение корня тыквы (Cucurbita pepo): 1—остатки первичной ксилемы (метаксилемы); 2—вторичная ксилема; 3—паренхимный радиальный луч; 4—камбий; 5—вторичная флоэма; 6—паренхима первичной коры; 7—перидерма
5.1.4. Образование и строение корневой системы растений
Переход корня от первичного анатомического строения к вторичному связан с утолщением корня и формированием корневой системы растения. Под корневой системой следует понимать совокупность всех корней растения. Образование корневой системы происходит благодаря ветвлению корней. Для большинства растений характерно моноподиальное ветвление, однако у плаунообразных корни ветвятся дихотомически. Ветвление корня можно рассматривать как особый тип роста, связанный с образованием боковых и придаточных корней. Боковыми называются корни, образующиеся на корнях. Корни, которые возникают на других частях растения, являются придаточными, или адвентивными (от лат. adventicus — пришлый, чуждый).
Боковые и придаточные корни закладываются эндогенно (рис. 67). Корнеродной тканью является перицикл. Зачатки боковых корней могут возникать в различных зонах перицикла, но чаще всего корневые бугорки образуются против элементов ксилемы. При заложении бокового корня клетки перицикла начинают делиться тангентально (периклинально), в результате чего образуется меристематический бугорок. По мере увеличения числа клеток направление делений меняется, клетки делятся не только тангентально, но и антиклинально. Увеличивающийся корневой бугорок давит на эндодерму, но, так как оболочка клеток эндодермы очень прочная, она не разрывается, а охватывает образующийся корешок и выпячивается в сторону паренхимы первичной коры.
Рис. 67. Заложение бокового корня в перицикле главного корня: 1—развивающийся боковой корень; 2—центральный цилиндр; 3—перицикл; 4—первичная флоэма; 5— первичная ксилема; 6—паренхима центрального цилиндра; 7—камбий, образовавшийся из паренхимы центрального цилиндра; 8—эндодерма; 9—паренхима первичной коры; 10—эпиблема; 11—корневые волоски
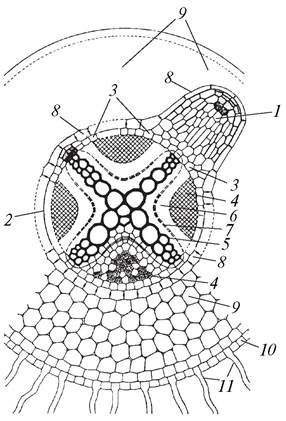
Клетки эндодермы делятся антиклинально, поэтому эндодерма остается однослойной и образует так называемый корневой кармашек — однослойный или многослойный. Он помогает формирующемуся корню проходить через паренхиму первичной коры и защищает его от повреждений.
По мере продвижения в первичной коре корневой кармашек или раздвигает клетки паренхимы, или, как полагают, благодаря выделению клетками ферментов способствует их разрушению. По мере роста в молодом корешке дифференцируется апекс, одетый корневым чехликом, и образуются центральный цилиндр и первичная кора. Растущий боковой корень проходит через первичную кору главного корня, прорывает ее и выходит наружу. При выходе на поверхность корневой кармашек разрывается и сбрасывается. С этого момента образовавшийся боковой корень начинает развиваться так же, как и главный. Главный корень с боковыми корнями связан посредством проводящей системы.
В зависимости от типа корней, которые входят в корневую систему растения, различают корневые системы стержневого и мочковатого типов (рис. 68).
Рис. 68. Типы корневых систем: А—стержневая (аллоризная); Б—мочковатая (гоморизная): 1—главный корень; 2—боковые корни; 3—придаточные корни
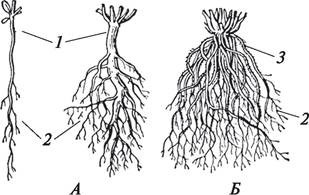
Стержневая корневая система состоит из хорошо выраженного главного корня и боковых корней, она всегда аллоризная (от греч. allos — другой). Кроме того, в состав такой корневой системы могут входить и придаточные корни. Она характерна преимущественно для двудольных.
Корневая система, в которой главный корень отсутствует или он не выделяется среди массы придаточных и боковых корней, называется мочковатой. Она преобладает у однодольных растений, но может формироваться и у двудольных. Мочковатая корневая система может быть аллоризной или гоморизной (от греч. homo — равный). Если гоморизная корневая система первична, тогда главный корень вообще не развивается (пшеница, рожь), если вторична, главный корень рано отмирает или не выделяется в массе придаточных корней (кукуруза).
Корневая система значительно увеличивает всасывающую поверхность корней и способствует более надежному закреплению растений в субстрате.
В пределах корневой системы наблюдается дифференцировка корней по выполняемым функциям. Особенно четко это проявляется у древесных растений. У них развиваются ростовые (скелетные) и сосущие (питающие) корни. Мощно развитые ростовые корни способны проникать на значительную глубину и расширять таким образом площадь питания растений. Они имеют вторичное строение и покрыты перидермой. Сосущие корни короткие и недолговечные. Эти корни ежегодно развиваются на скелетных корнях, имеют первичное строение, осуществляют всасывание воды с растворенными в ней минеральными веществами и к осени обычно отмирают. Ростовые корни могут проникать на значительную (максимальную) глубину, сосущие корни обычно располагаются на рабочей глубине, т. е. на глубине, на которой находится большая часть корней растения.
5.1.5. Симбиоз и его типы
Растение взаимодействует не только с абиотическими факторами внешней среды, но и с живыми организмами, населяющими почву. Различные формы совместного сосуществования организмов (комменсализм, паразитизм, мутуализм) немецкий миколог и анатом растений Антон де Бари (1831—1888) в 1879 г. назвал симбиозом (от греч. simbiosis — совместная жизнь). Впоследствии в 1906 г. зоолог О. Гертвиг (1849—1922) сузил границы данного термина и предложил называть симбиозом такие отношения, которые взаимовыгодны для обоих симбионтов. В таком понимании этот термин был воспринят русскими учеными и прочно укоренился в ботанической литературе. Симбиоз может быть облигатным и факультативным. При облигатном симбиозе один из партнеров оказывается в такой зависимости от другого, что существовать без него не может. Облигатный симбиоз характерен для представителей семейства Орхидные. При факультативном симбиозе каждый из симбионтов может нормально существовать при отсутствии другого партнера. Факультативный симбиоз присущ большинству древесных растений (ель, береза, осина, дуб и т. д.).
У высших растений наиболее широко распространен симбиоз с различными грибами и почвенными бактериями из рода Rhizobium. Относительно недавно установлено, что симбионтами некоторых растений могут быть актиномицеты.
Симбиоз мицелия грибов с корнями высших растений называется микоризой (от греч. mykes — гриб и rhiza — корень). Микориза — сложная симбиотическая система, в которой гифы гриба и корни растений взаимодействуют по- разному. Микориза может быть эктотрофной (эктомикоризой), эндотрофной (эндомикоризой) и эктоэндотрофной (эктоэндомикоризой) (рис. 69).
Рис. 69. Типы микоризы: А—эктотрофная микориза дуба обыкновенного (Quercus robur); Б, В—эндотрофная микориза в клетках корня пальчатокоренника мясо-красного (Dactylorhiza incarnata) на разных стадиях развития растения: 1—гифы гриба
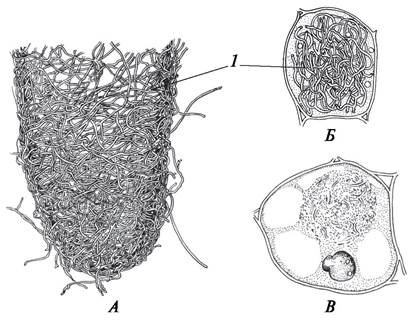
При эктотрофной микоризе гифы гриба оплетают кончики молодых корней. Эктомикоризные корни короткие, утолщенные, разветвленные наподобие кораллов. На таких корнях корневые волоски не образуются, а функцию всасывания воды с растворенными в ней минеральными веществами выполняют гифы гриба. Эктотрофная микориза характерна для большинства лиственных и хвойных деревьев.
У представителей семейства Орхидные гифы гриба проникают в клетки паренхимы первичной коры корня и образуют там клубковидные скопления. Такой тип микоризы называется эндотрофной. Участки корня, в которые проникли гифы гриба, имеют буровато-коричневую окраску, чем и отличаются от тех участков, где гиф нет. У орхидных гифы гриба обнаруживаются в клетках на ранних стадиях развития растения. После цветения растений гифы гриба начинают исчезать. Предполагают, что постепенно растение-хозяин «переваривает» своего симбионта. На этом основании эндотрофную микоризу орхидных некоторые ботаники рассматривают как случай контролируемого паразитизма.
Эктоэндотрофная микориза отличается от эндотрофной тем, что гифы гриба проникают не в клетки, а в межклетники паренхимы первичной коры. Такой тип микоризы встречается у вересковых (вереск, брусника, клюква).
Гифы гриба значительно увеличивают всасывающую поверхность корня и улучшают питание растений. Питание растений с помощью гиф гриба называется микотрофным. Особенности биологического взаимодействия симбионтов выяснены недостаточно. С помощью меченого углерода (C14) и фосфора (P32) установлено, что грибы, образующие микоризу с вереском и клюквой, могут транспортировать в растения соединения, меченые по фосфору. Соединения, меченые по углероду, из гиф гриба в корень растения не передаются. В то же время продукты фотосинтеза растений, меченые по углероду, обнаруживались в мицелии гриба. У орхидных происходит обмен витаминами или их компонентами. Гифы гриба не только улучшают питание растений, но и их водоснабжение. Микориза защищает также растение от фитопатогенных организмов. Микориза широко распространена в природе и характерна как для голосеменных, так и для покрытосеменных растений. Не отмечена микориза у представителей семейств Крестоцветные, Маковые, Крапивоцветные, а также у насекомоядных и паразитирующих растений.
Систематическое положение грибов-микоризообразователей различно. У орхидных микоризу образуют несовершенные грибы из рода Rhizoctonia, у вересковых — из рода Phoma. Микоризообразователями с древесными растениями являются многие представители базидиальных грибов, а с травянистыми преимущественно зигомицеты.
Классическим примером симбиоза с бактериями является образование клубеньков на корнях растений семейства Бобовые (рис. 70). Впер вые клубеньки на корнях бобовых были описаны известным русским микологом М. С. Ворониным (1838—1903) в 1866 г. Клубеньковые бактерии относятся к роду Rhizobium. Основной особенностью представителей этого рода является способность фиксировать молекулярный азот. При отсутствии бобовых растений клубеньковые бактерии могут жить в почве как сапротрофы. Различные виды рода Rhizobium обладают достаточно выраженной видоспецифичностью. Так, Rhizobium trifolii поселяется на корнях клеверов, Rh. japonicum — на корнях сои, Rh. melilotii — на корнях люцерны и донника, Rh. leguminosarum — на корнях гороха, вики, бобов. Однако не все бобовые могут вступать в симбиоз с клубеньковыми бактериями. Ученые считают, что из 13 тыс. бобовых растений к симбиозу способны всего около 10 %.
Рис. 70. Клубеньки на корнях люпина (Lupinus polyphullus): А—общий вид корневой системы; Б—поперечный срез корня с клубеньком: 1—клубеньки; 2—покровная ткань корня; 3—паренхима; 4—флоэма; 5—камбий; 6—радиальный паренхимный луч; 7—первичная ксилема; 8—вторичная ксилема; 9—бактероидная ткань; 10—проводящие ткани корня
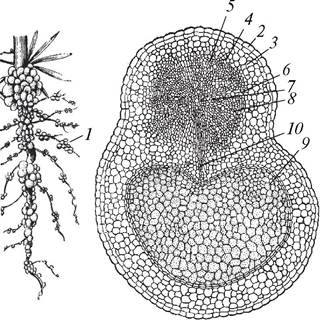
Клубеньковые бактерии проникают в корень через корневые волоски и внедряются в периферические клетки первичной коры. Растение реагирует на это чужеродное внедрение активным делением паренхимных клеток. В результате на поверхности корня образуются характерные для различных видов растений вздутия, которые называют клубеньками. В клубеньках бактерии меняют свою форму, превращаются в бактероиды, интенсивно размножаются и образуют так называемую бактероидную ткань. Бактероиды активно связывают молекулярный азот и переводят его в соединения, которые могут усваиваться растением. В то же время растение обеспечивает бактерии необходимыми питательными веществами. Фиксированный клубеньковыми бактериями азот накапливается в растениях, поэтому многие бобовые хорошо развиваются на почвах, бедных азотом. Кроме того, они способствуют накоплению азота в почве, поэтому бобовые культуры являются лучшими предшественниками для сельскохозяйственных и декоративных растений. С целью обогащения почвы азотом такие бобовые, как клевер, люпин используют в качестве сидерационных культур.
Сравнительно недавно ученым удалось обнаружить клубеньки на корнях некоторых деревьев и кустарников, относящихся к семействам Буковые, Березовые, Крушиноцветные, Казуариновые, Розоцветные и др.
У этих растений симбионтами, вызывающими образование клубеньков, являются актиномицеты. Как и клубеньковые бактерии, они способны фиксировать молекулярный азот и переводить его в доступные для растений соединения.
При симбиозе функция корней не меняется, а появляющиеся некоторые морфологические особенности не являются генетически обусловленными и не наследуются.
5.1.6. Метаморфозы корня
Под влиянием условий существования корни могут менять свои функции, что приводит к существенному изменению их морфологических и анатомических особенностей, т. е. к метаморфозам. Метаморфоз — это наследственно закрепленное видоизменение органа, происходящее под влиянием условий существования и связанное с изменением выполняемой функции.
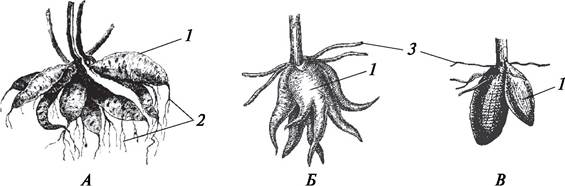
Метаморфозы корней довольно разнообразны. Если у корней преобладает запасающая функция, они сильно разрастаются за счет образования большого количества паренхимы. Утолщенные боковые или придаточные корни называют корневыми шишками. Корневые шишки формируются у георгины (сем. Сложноцветные), чистяка (сем. Лютиковые), пальчатокоренника (сем. Орхидные). При развитии корневых шишек паренхима в большом количестве образуется в первичной коре. Форма и размеры корневых шишек различны. Кроме запасающей функции корневые шишки растений могут выполнять функцию размножения. На самих корневых шишках почек возобновления нет, но они образуются у основания побегов, которые связаны с корневыми шишками (рис. 71).
Рис. 71. Внешний вид запасающих корней — корневых шишек разных растений: А—георгина культурная (Dahlia cultorum); Б—пальчатокоренник мясо-красный (Dactylorhiza incarnata); В—ятрышник мужской (Orchis mascula): 1—корневая шишка; 2—питающие корни; 3—типичные придаточные корни
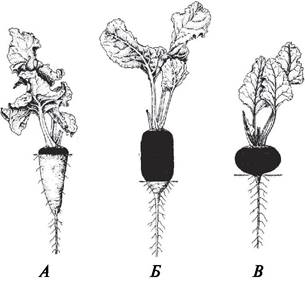
При разрастании главного корня и подземной части стебля образуются корнеплоды (рис. 72). Форма корнеплодов и их размеры различны. Корнеплод состоит из головки, которая представляет собой укороченный стебель и несет листья, шейки (гипокотиля) и собственно корня. Определить, какая часть корнеплода образовалась из корня, можно по наличию боковых корней.
Рис. 72. Внешний вид корнеплодов свеклы (Beta vulgaris): участие главного корня и гипокотиля (затемнен) в образовании корнеплода: А—сахарная свекла; Б—кормовая;
В—столовая
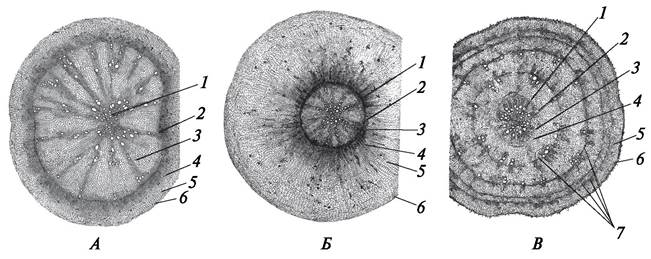
У различных растений при переходе корня от первичного анатомического строения к вторичному, корнеплоды формируются по-разному (рис. 73). У представителей семейства Крестоцветные (редька, редис, репа, турнепс) утолщение корнеплода происходит благодаря разрастанию паренхимы в области вторичной ксилемы. У зонтичных (морковь, петрушка, сельдерей, пастернак) паренхима в большом количестве образуется во вторичной флоэме. Особый тип корнеплода у свеклы. После формирования вторичного анатомического строения корень свеклы начинает приобретать третичное строение, благодаря чему развивается корнеплод. По мере заложения каждого нового листа в корнеплоде свеклы образуется кольцо добавочного камбия, который производит слабо развитые сосудисто-волокнистые пучки и большое количество паренхимы. Начало первому добавочному камбию дает камбий, образовавшийся из паренхимы перициклического происхождения, возникшей при формировании вторичного анатомического строения. Периферический слой первого добавочного камбия дает начало второму добавочному камбию и производит слабо развитые проводящие пучки коллатерального типа и паренхимную ткань. Часть клеток паренхимной ткани способна к делению, поэтому паренхимный слой быстро расширяется. Периферический слой второго добавочного камбия дает начало третьему добавочному камбию и т. д. Чем ближе к поверхности корнеплода, тем уже кольца паренхимы и тем более редуцированы сосудисто-волокнистые пучки. На поперечном разрезе через корнеплод хорошо заметны чередующиеся светлые (участки паренхимы) и более темные (участки добавочных камбиев и их производных) концентрические круги, количество их соответствует числу заложившихся слоев добавочных камбиев и числу образовавшихся на растении листьев.
Рис. 73. Анатомическое строение корнеплодов: А—редька посевная (Raphanus sativus); Б—морковь посевная (Daucus sativus); В—свекла обыкновенная (Beta vulgaris): 1 —остатки первичной ксилемы (метаксилемы); 2—камбий; 3—элементы вторичной ксилемы; 4—элементы вторичной флоэмы; 5—паренхима первичной коры; 6— перидерма; 7—добавочные слои камбия и сформированные ими элементы проводящих тканей и паренхимы
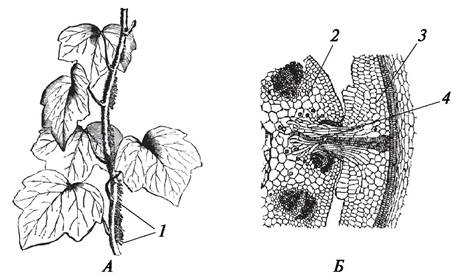
У некоторых лиан (плющ обыкновенный — Hedera helix, девичий виноград пятилисточковый — Parthenocissus quinquefolia) придаточные корни на стебле превращаются в корни-присоски, при помощи которых растение прикрепляется к опоре и поддерживает вертикальное положение.
Своеобразные корни-присоски образуются у растений-паразитов и полупаразитов. Такие корни-присоски называются гаусториями. Они проникают в ткани растения-хозяина, их проводящая система подходит к проводящей системе растения-хозяина и внедряется в нее. При помощи таких метаморфизированных корней растения-паразиты и полупаразиты получают питательные вещества из тканей растения-хозяина (рис. 74).
Рис. 74. Метаморфозы корня: А—плющ обыкновенный (Hedera helix); Б—повилика европейская (Cuscuta europaea) на стебле клевера: 1—корни-присоски (корни-прицепки); 2—поперечный разрез через стебель клевера; 3—продольный разрез через стебель повилики; 4—гаустория повилики в проводящей системе клевера
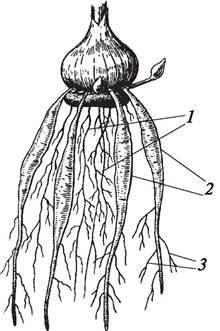
Контрактильные, или втягивающие, корни, которые развиваются у однодольных луковичных (тюльпаны, лилии, луки) (рис. 75) и клубнелуковичных растений (гладиолусы, крокусы) (рис. 76), также являются примером метаморфоза корней. В клетках контрактильных корней содержится большое количество сахаров, которые быстро расходуются в процессе роста растения. Благодаря «опустошению» клеток контрактильный корень приобретает поперечную морщинистость, сильно укорачивается и углубляет залегание луковиц и клубнелуковиц растений-эфемероидов. Это имеет большое биологическое значение, так как в условиях степей и полупустынь помогает растениям, впадающим в состояние покоя, успешно избежать недостатка влаги в засушливый период. Контрактильные корни могут также развиваться и у некоторых двудольных растений (пион — Paeonia).
Рис. 75. Контрактильные корни лилии саранки (Lilium martagón), ежегодно заглубляющие луковицу
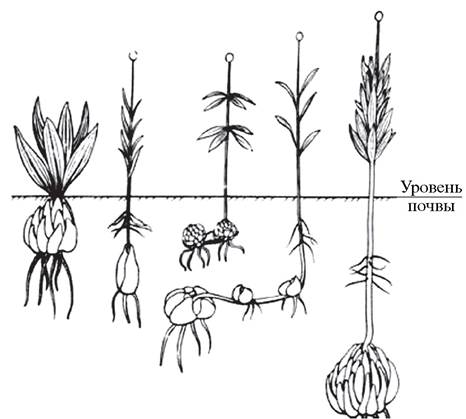
Рис. 76. Корневая система гладиолуса: 1—придаточные и боковые корни; 2—контрактильные (втягивающие) корни; 3—боковые корни
У тропических растений, растущих по побережьям океанов, формируются специальные дыхательные корни — пневматофоры. В таких корнях не только хорошо развита аэренхима, как у всех водно-болотных растений умеренной зоны, но они растут вертикально вверх, поднимаются над поверхностью воды и имеют систему особых отверстий, через которые воздух поступает в корень (рис. 77).
Рис. 77. Дыхательные корни (пневматофоры) южноамериканского водного растения жюссеи ползучей (Jusseia repens): 1—стебель; 2—придаточные корни; 3—пневматофоры
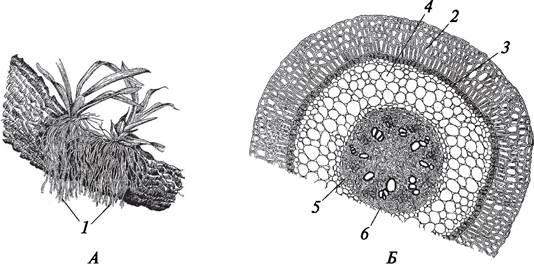
У многих тропических эпифитных орхидей развиваются воздушные корни (рис. 78). Такие корни способны выполнять функцию фотосинтеза и улавливать влагу, которая попадает на них в виде дождя или росы. Воздушные корни имеют зеленую окраску благодаря наличию в их клетках хлоропластов. На поверхности корней образуется особая многослойная мертвая ткань — веламен, которая абсорбирует влагу из воздуха. Под веламеном находится слой экзодермы, клетки которой утолщены так же, как и клетки эндодермы. Как и в эндодерме, в экзодерме есть пропускные клетки. Таким образом, у эпифитных орхидей экзодерма и эндодерма играют роль физиологических барьеров и регулируют поступление воды из веламена в центральный цилиндр, а из него в периферические зоны корня.
Рис. 78. Метаморфозы корня: А—эпифитные орхидеи на стволе дерева; Б—анатомическое строение воздушного корня орхидеи: 1—воздушные корни; 2—веламен; 3— экзодерма; 4—паренхима первичной коры; 5—эндодерма (3—5 — первичная кора); 6—центральный цилиндр
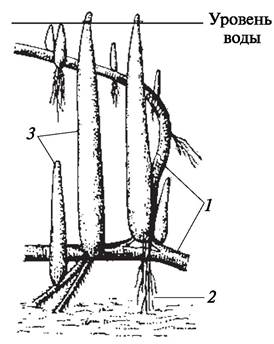
Своеобразными метаморфизированными корнями являются ходульные корни растений мангровых зарослей (ризофора — Rhizophora, авиценния — Avicennia) и корни-подпорки некоторых фикусов (баньян — Ficus bengalensis). По происхождению это придаточные корни. Придаточными корнями являются и уплощенные досковидные корни. Благодаря наличию большого количества таких корней растения могут поддерживать огромные кроны деревьев и нормально развиваться, находясь в крайне сложных условиях существования (рис. 79).
Рис. 79. Корни-подпорки баньяна — фикуса бенгальского (Ficus bengalensis)

Таким образом, типичные корни высших растений имеют различное происхождение, а также анатомическое и морфологическое строение, удерживают растение в почве и осуществляют почвенное питание растений: обеспечивают растения водой и растворенными в ней минеральными веществами. Строение и функции метаморфизированных и типичных корней существенно отличаются между собой, что позволяет растениям с метаморфизированными корнями наилучшим образом приспосабливаться к конкретным условиям существования.