МОРФОЛОГИЯ РАСТЕНИЙ - Т. А. Сауткина - 2012
ГЛАВА 4. РАЗВИТИЕ ФОРМЫ ТЕЛА У РАСТЕНИЙ
4.5. Усложнение в ходе эволюции анатомического строения растений. Эволюция типов стели
По мере усложнения внешнего строения растений совершенствуется и их внутреннее строение. В первую очередь следует отметить усложнение строения центрального цилиндра, или стели.
Основы стелярной теории заложил французский ботаник Ф. Ван-Тигем в конце XIX в. Он определил понятие «стель» (стела) (от лат. stela, греч. stele — столб, колонна) как совокупность первичных проводящих тканей и перицикла. Ф. Ван-Тигему удалось показать, что стель является для сосудистых растений универсальным элементом стебля и корня, но у представителей различных таксонов она имеет разное строение. Дальнейшее развитие стелярная теория получила в работах ряда ученых. Особенно большой вклад в развитие стелярной теории внесли немецкий ученый В. Циммерман и российские ботаники К. И. Мейер и А. Л. Тахтаджян, которые показали, что строение стели изменяется у разных таксонов и в ходе эволюции, и в процессе онтогенеза.
Самым примитивным типом стели считается протостель (от греч. protos — первый), которая была выявлена у ринии. Она представляла собой тяж проводящих элементов, в центре которого находилась ксилема, окруженная флоэмой. Часто протостель, имеющую на поперечном разрезе округлое очертание, называют гаплостелью (от греч. haplos — одиночный). Протостель защищена первичной корой. У современных растений протостелическое строение центрального цилиндра можно обнаружить в похожих на ризомоиды корневищах псилота и в органах молодых примитивных папоротников — гроздовника, ужовника, гельминтостахиса.
Усложнение типов стели связано с развитием листьев и боковых ветвей, что потребовало усовершенствования проводящей системы. Английский ботаник Ф. О. Боуэр (1855—1948) указывал, что передача веществ пропорциональна поверхности соприкосновения живых и мертвых проводящих тканей. На основании изучения строения центрального цилиндра у ископаемых и современных растений удалось выявить типы стелей у различных таксонов и установить их эволюционное взаимоотношение. Согласно имеющимся данным усложнение типов стели происходило благодаря увеличению площади соприкосновения ксилемы и флоэмы, а также путем витализации (от лат. vitalis — живой) — появления в центральном цилиндре большого количества живой паренхимной ткани.
Изменение площади соприкосновения ксилемы и флоэмы привело к образованию актиностели (от греч. aktis — луч), для которой характерно лопастное или звездчатое очертание ксилемы, со всех сторон окруженной флоэмой. Такой тип стели был обнаружен у астероксилона, а также встречается в молодых побегах некоторых современных плаунообразных. В старых побегах плаунов центральный цилиндр состоит из ксилемы, разбитой на обособленные ленты, вокруг которых расположена флоэма. Перицикл почти не отличается от внутреннего слоя первичной коры — эндодермы. Такой тип стели называется плектостелью (от греч. plectos — сплетенный). Актиностель и плектостель часто рассматривают как разновидности протостели. Протостель, актиностель и плектостель характерны для представителей микрофильной линии эволюции.
У представителей макрофильной линии эволюции усложнение строения стели связано с процессом витализации. От протостели могли возникнуть два типа сифоностели (от греч. siphon — трубка) — эктофлойная и амфифлойная, или соленостель (от греч. solon — трубка). В сифоностели в центре ксилемы появляется сердцевина из живой паренхимной ткани, которая начинает выполнять запасающую функцию. У эктофлойной сифоностели за ксилемой располагается флоэма, окруженная перициклом. Весь центральный цилиндр защищен первичной корой. Внутренний слой первичной коры — эндодерма — состоит из тонкостенных клеток. Перемещение ксилемы к периферии увеличило механическую прочность осевых органов. Эктофлойная сифоностель (или просто сифоностель) характерна для некоторых современных папоротников, у которых образуется небольшое количество относительно мелких листьев — гроздовник, марсилея. У некоторых древних папоротникообразных, имеющих крупные (от 2 до 6 м длины) листья (осмунда, мараттиевые), в молодых органах развивается соленостель. Она имеет специфические отличия от сифоностели. Сердцевина окружена внутренней эндодермой. За ней формируются проводящие ткани, включающие наружную и внутреннюю флоэму и заключенную между ними ксилему. К наружной флоэме примыкает перицикл, ограничивающий центральный цилиндр, за которым следует первичная кора.
Для большинства современных папоротникообразных характерна диктиостель (от греч. dictyon — сеть). Она формируется из соленостели, в которую впадает множество листовых следов (совокупность сосудисто-волокнистых пучков листа), что приводит к образованию большого числа листовых прорывов, заполненных паренхимой. Часть стели между двумя листовыми прорывами называется меристелью. Каждая меристель имеет вид концентрического пучка, внутри которого расположена ксилема, окруженная флоэмой, перициклом и хорошо выраженной эндодермой. Размеры и форма меристелей различны. Обычно они располагаются в виде кольца и погружены в паренхиму. У некоторых папоротников (орляк) меристели могут располагаться и в первичной коре, образуя там один или более циклов. Такие типы диктиостели называют соответственно дициклической или полициклической диктиостелью.
Для покрытосеменных растений с их сильно разветвленными осевыми органами и большим количеством листьев характерны два типа стели: эустель и атактостель. Эустельобразуется из сифоностели и представляет собой круговую систему расположенных в один ряд коллатеральных пучков, разделенных первичными сердцевинными лучами. Этот тип стели присущ двудольным растениям. На поперечных разрезах через молодые корень и стебель видны довольно существенные различия в строении эустели. В корне нет сердцевины, но хорошо развиты перицикл и эндодерма. В молодом стебле четко сформирована сердцевина, но перицикл и эндодерма выражены нечетко. Кроме того, образование ксилемы и флоэмы в корне и стебле происходит по-разному. У однодольных, центральный цилиндр содержит большое количество беспорядочно расположенных коллатеральных пучков закрытого типа. Такой тип стели называется атактостелью (от греч. atactos—беспорядочный). Он образуется благодаря тому, что из листа однодольных в стебель входит множество листовых следов, которые сохраняют в паренхиме центрального цилиндра свою самостоятельность (рис. 60). У многих однодольных перицикл очень часто модифицируется в механическую ткань — перициклическую склеренхиму, а кора нечетко отделена от центрального цилиндра.
Рис. 60. Схема эволюции типов стели: 1—протостель; 2—актиностель; 3—плектостель; 4—эктофлойная сифоностель; 5—артростель; 6—амфифлойная сифоностель (соленостель); 7—диктиостель; 8—эустель; 9—атактостель (заштрихована первичная ксилема, точками обозначена первичная флоэма)
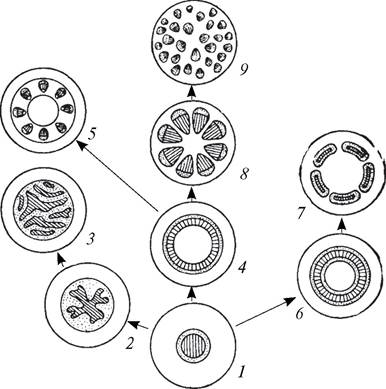
Все типы стели эволюционно связаны между собой, и каждый из них характерен для определенных таксонов высших растений. Следовательно, стелярная теория имеет большое значение не только для морфологии и систематики растений, но и для познания филогенеза в целом.
4.6. Направление эволюции. Формы тела у высших растений
Как было указано ранее, в процессе эволюции вегетативная сфера растений чаще всего развивалась в направлении усложнения внешнего и внутреннего строения органов и дифференциации их функций.
Современные высшие растения (за исключением мохообразных) — это сложно устроенные организмы с хорошо развитыми системами тканей и вегетативных органов, что позволяет им существовать в разнообразнейших экологических нишах.
Однако развитие высших растений в условиях суши шло не только по пути морфофизиологического прогресса. В связи с переходом к водному образу жизни, а также с явлениями сапротрофизма и паразитизма у некоторых высших растений произошло упрощение как морфологического, так и анатомического строения. Следовательно, в развитии формы тела у высших растений можно выявить три основных направления эволюции: ароморфоз, идиоадаптацию и дегенерацию.
Появление вегетативных органов, образование тканей, формирование стели, несомненно, представляют собой ароморфозы — качественные скачки, существенно повысившие уровень организации растений и позволившие им приспособиться к новым условиям жизни.
Различное строение листьев у растений разных местообитаний: гидрофитов (кувшинки, рдесты) и гигрофитов (калужница болотная) — растений переувлажненных мест обитаний; ксерофитов (вереск, очитки) — растений засушливых мест обитания; мезофитов — растений умеренно увлажненных мест — следует рассматривать как пример идиоадаптации, т. е. как возникновение частных приспособлений в строении и функции органов при сохранении общего плана организации предковых форм.
И, наконец, в растительном мире можно найти примеры дегенерации — упрощения строения органа, вплоть до полного исчезновения (абортирования). У растений-паразитов (повилика — Cuscuta, заразиха — Orobanche) исчезают листья и корень. У водных свободноплавающих покрытосеменных растений (ряска — Lemna) корень есть, но отсутствует корневой чехлик и не образуются корневые волоски. У водного папоротника сальвинии плавающей (Salvinia natans) корень вообще не образуется, его функцию выполняет сильно рассеченный и густо опушенный подводный лист. Кроме того, у сальвинии упрощается тип стели: вместо диктиостелического центрального цилиндра у нее формируется протостелический. Дегенерация органов или каких-то других структур — своеобразная реакция растения на конкретные условия существования.