МОРФОЛОГИЯ РАСТЕНИЙ - Т. А. Сауткина - 2012
ГЛАВА 4. РАЗВИТИЕ ФОРМЫ ТЕЛА У РАСТЕНИЙ
4.3. Изменение морфологического строения растений. Эволюция типов ветвления
Изменения морфологических особенностей высших растений связаны прежде всего с изменением и совершенствованием типа ветвления, а также с появлением особых специализированных органов фотосинтеза — листьев.
Изучение палеоботанического материала позволило установить, что исходным типом ветвления было дихотомическое. Именно к такому выводу пришел в 1912 г. немецкий ботаник Г. Потонье (1857—1913). При дихотомическом ветвлении образование боковых ветвей происходит благодаря разделению апикальной меристемы, поэтому его называют верхушечным. При дихотомическом ветвлении количество боковых ветвей увеличивается в геометрической прогрессии. На оси первого порядка образуется две оси второго порядка, на двух осях второго порядка — четыре оси третьего порядка и т. д. Таким образом, у растения возникает многовершинность, что приводит к разрастанию его в горизонтальной плоскости. Дихотомически ветвящиеся растения никогда не бывают высокорослыми.
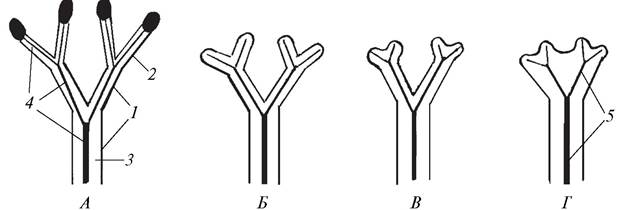
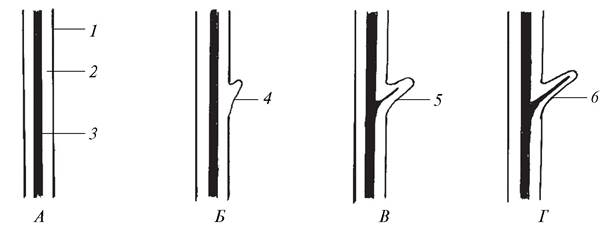
Различают два вида дихотомического ветвления — изотомное (от греч. isos — равный) и анизотомное (от греч. anisos — неравный). При изотомной дихотомии обе веточки, образовавшиеся при делении верхушки каждой оси, растут равномерно и имеют одинаковую длину. Такой тип ветвления характерен для растений, обитающих в относительно стабильной среде (в воде, в сильно загущенном травостое). Из современных высших растений по типу изотомной дихотомии ветвятся риччия плавающая (водный представитель печеночных мхов), представители родов дифазиаструм, баранец из отдела Плаунообразные. Под влиянием условий существования у высших растений сформировалась анизотомная дихотомия, при которой образующиеся веточки стали развиваться неравномерно, в результате чего одна перерастала в другую. Анизотомное дихотомическое ветвление было обнаружено уже у некоторых ископаемых растений. Оно может рассматриваться как начавшийся процесс перевершинивания, т. е. переход растения от многовершинности к появлению одной вершины, а, следовательно, к выделению одной главной оси. Из ныне живущих растений анизотомная дихотомия характерна для плауна булавовидного и ряда других представителей плаунообразных. Резкое отставание в росте одной из веточек привело к появлению дихоподиального (от греч. dicha — отдельно и pus (podos) — нога, здесь — ось) ветвления, при котором образуется главная зигзагообразная, вертикально расположенная ось растения. Дихоподиальное ветвление было присуще ископаемому растению астероксилону. Появление этого типа ветвления надо рассматривать как весьма важное событие в жизни наземных растений. У растений с дихоподиально ветвящимися стеблями спорангии оказались значительно приподнятыми над землей, что обеспечивало эффективное распространение спор (рис. 55).
Рис. 55. Схема верхушечных типов ветвления: А—дихотомическое изотомное; Б—дихотомическое анизотомное; В—дихоподиальное (цифрами обозначены последовательно формирующиеся побеги)
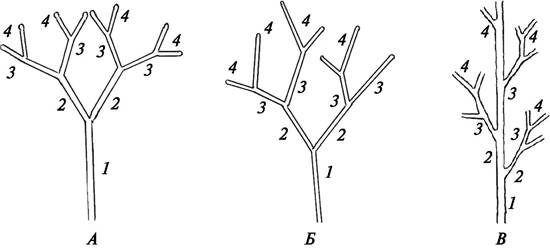
Начавшийся у древних растений процесс перевершинивания закрепил в конечном счете образование главной оси и привел к возникновению моноподиального (от греч. monos — один, единственный) ветвления. При моноподиальном ветвлении главная ось может довольно длительное время относительно неограниченно нарастать в длину, что способствовало увеличению высоты растения. В то же время при моноподиальном ветвлении изменился характер образования боковых ветвей, они стали возникать не из верхушечной меристемы, а из боковых (реже и из придаточных) почек, поэтому данный тип ветвления называют также боковым. Боковые ветви, как и главная ось, растут относительно неограниченно. В силу таких особенностей роста на осях разного порядка формируется неограниченное число олиственных боковых ветвей и, как следствие этого, сильно увеличивается фотосинтезирующая поверхность. Поскольку у моноподиально ветвящихся растений развитие боковых ветвей чаще всего происходит в акропетальном направлении (от основания к верхушке), у растений, особенно древесных, наиболее сильно развиты нижние боковые ветви и формируется пирамидальная крона. Такой тип моноподиального ветвления называется базитонным. У некоторых акаций более интенсивно при моноподиальном ветвлении развиваются верхние ветви, и крона принимает зонтиковидную форму. Такое моноподиальное ветвление называется акротонным. При мезотонном ветвлении наиболее сильные, длинные ветви образуются в средней части основного побега. При таком типе моноподиального ветвления у растений формируется колоновидная форма кроны. Моноподиальный тип ветвления довольно древний, он характерен для подавляющего большинства голосеменных растений (рис. 56).
Рис. 56. Схема различных видов моноподиального ветвления: А—акротонное ветвление; Б—базитонное ветвление; В—мезотонное ветвление (цифрами обозначены последовательно формирующиеся побеги)
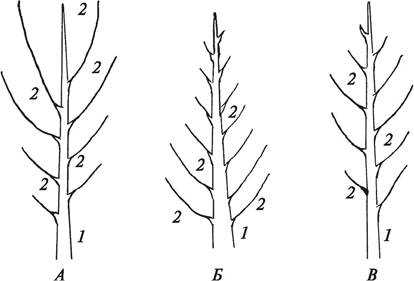
К боковому ветвлению относится и симподиальное (от греч. syn — вместе). При симподиальном ветвлении, как и при моноподиальном, образуется главная ось, но она состоит из осей разных порядков. Образование такой главной оси происходит следующим образом. Ось первого порядка начинает расти, но в какой-то момент верхушечный рост прекращается, точка роста отмирает или на верхушке формируется цветок. Как только главная ось (ось первого порядка) перестает нарастать в длину и прекращается апикальное доминирование, в рост трогается ближайшая к верхушке боковая почка, из которой формируется ось второго порядка. Сначала она располагается под некоторым углом к первоначальной оси, а затем принимает ее направление. И так происходит неоднократно, в результате чего главная ось представляет собой систему побегов. Такой характер ветвления приводит к тому, что при последовательном отмирании апикальных меристем осей разных порядков в рост трогается большое количество почек (боковые, придаточные), что приводит к сильному увеличению надземной скелетной части растения, а, следовательно, и увеличению фотосинтезирующей поверхности. Симподиальное ветвление считается вершиной эволюции типов ветвления растений. Оно характерно преимущественно для покрытосеменных растений. Однако в чистом виде симподиальное ветвление характерно только для травянистых растений. У древесных растений симподиальное ветвление сочетается с моноподиальным. Признаки симподиального ветвления в виде отмерших участков побега предыдущего порядка можно обнаружить у деревьев только на тронувшихся в рост молодых веточках. При наличии симподиального ветвления у растений образуется раскидистая крона (липа, береза, яблоня).
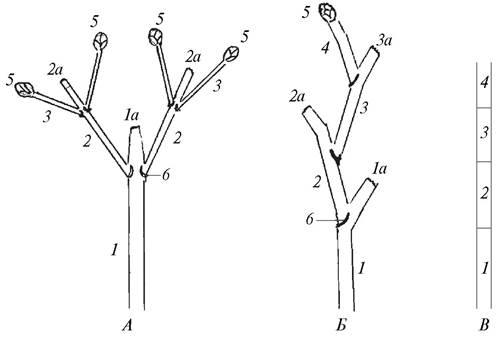
Боковым является также и ложнодихотомическое ветвление. При этом ниже верхушки исходного побега, которая отмирает или заканчивается цветоносом, из пазушных почек формируются два боковых побега следующего порядка. Такой тип ветвления характерен для многих представителей семейства Гвоздичные. Ложнодихотомическое ветвление встречается также у некоторых деревьев (конский каштан, клен) и кустарников (сирень, свидина). У древесных растений ложнодихотомическое ветвление так же, как и симподиальное, сочетается с моноподиальным (рис. 57).
Рис. 57. Схемы ложнодихотомического (А) и симподиального (Б) типов ветвления, строения основной оси (ствола) (В) у деревьев с симподиальным типом ветвления:
1—4—последовательно формирующиеся побеги; 1а—3а—отмирающие верхушки побегов; 5—верхушечные почки; 6—листовые рубцы
Появление главной оси позволило растениям приподняться над поверхностью суши и приблизиться к источнику энергии, используемой в процессе фотосинтеза.
4.4. Появление в процессе эволюции и развитие филлоидов и листьев
Ветвление было не единственным способом увеличения поверхности высших растений. Если у самых примитивных ископаемых растений (куксонии, ринии) процесс фотосинтеза осуществлялся осевыми частями тела, которые освещались неравномерно, то у более продвинутых появились специальные уплощенные боковые органы — листья. Они были ориентированы более или менее горизонтально, что способствовало лучшему улавливанию света и более эффективному процессу фотосинтеза. Возникновение листьев сильно увеличило надземную поверхность растений, что активизировало процессы ассимиляции, газообмена и транспирации и в то же время повысило уровень их организации.
Согласно существующим представлениям листья в ходе эволюции могли возникнуть двумя путями. Наличие небольших шипиков у псилофита и более крупных шиловидных образований у астероксилона свидетельствует о том, что они могли образоваться путем формирования поверхностных выростов (энациев) осевых частей. В такие выросты — филлоиды — от проводящей системы мезомов отходил проводящий пучок, по которому продукты ассимиляции передвигались от филлоида (листа) в мезомы и ризомоиды. По нему же вода с растворенными веществами доставлялась к филлоиду. Листья, возникшие энационным путем, мелкие, травянистые или чешуевидные. Они характерны для представителей микрофильной (от греч. micros — малый и phyllon — лист) линии эволюции. Из современных растений представителями микрофильной линии эволюции являются плаунообразные (рис. 58).
Рис. 58. Схема образования в ходе эволюции энационных (микрофильных) листьев (филлоидов): А—Г — последовательное формирование филлоидов: 1—фрагмент мезома псилофитов; 2—первичная кора; 3—проводящая система стели; 4—начало образования бокового выроста (энация); 5—начало отхождения проводящей системы из стели в энаций; 6—завершение образования энационного листа и его проводящей системы (одной жилки)
В отличие от плаунообразных у папоротникообразных и других таксонов высших споровых и семенных растений, согласно теломной теории, листья
образовались путем кладофикации (от греч. klados — ветвь), т. е. имеют осевое происхождение. Такие листья могли сформироваться в результате срастания, укорачивания и уплощения теломов, утративших спорангии. Для кладофикационных (синтеломных) листьев характерно нарастание верхушкой, что присуще осевым структурам. Листья, образовавшиеся путем кладофикации, имеют достаточно крупные размеры. Проводящая система листа объединяется с проводящей системой оси растения с образованием листовых прорывов (паренхимных участков, которые возникают в проводящей системе осевой структуры выше места впадения в нее листового пучка — листового следа). Кладофикационные листья характерны для представителей макрофильной (от греч. macros — большой) линии эволюции (рис. 59).
Рис. 59. Схема образования в ходе эволюции кладофикационных (синтеломных) листьев: А—фрагмент псилофита; Б—фрагмент псилофита с теломами, утратившими спорангии; В—укорочение и срастание стерильных теломов; Г—плоский лист: 1—фрагмент мезомов; 2—теломы; 3—первичная кора; 4—проводящая система мезомов и теломов; 5—проводящая система сформированного кладофикационного листа