МОРФОЛОГИЯ РАСТЕНИЙ - Т. А. Сауткина - 2012
ГЛАВА 4. РАЗВИТИЕ ФОРМЫ ТЕЛА У РАСТЕНИЙ
4.1. Понятие об эволюции. Эволюция формы тела у примитивных организмов
Даже при самом поверхностном знакомстве растительный мир поражает своим разнообразием. Одноклеточные микроскопические водоросли и гиганты растительного мира секвойи, достигающие 100 и более метров; растения, обитающие в воде, и суккуленты пустынь; самое маленькое цветковое растение вольффия, не превышающее 1 мм, с цветком около 0,5 мм, и раффлезия туан-мудэ, цветки которой достигают до 1 м в диаметре и т. д. Такое сопоставление можно продолжать до бесконечности, ибо к настоящему времени ученые насчитали около 40 000 видов водорослей и 300 000 видов высших растений и все еще продолжают открывать новые виды.
Как могло возникнуть такое разнообразие — вот один из первых проблемных вопросов, на который должна ответить морфология растений. На этот вопрос наука отвечает однозначно: все многообразие растительного мира могло возникнуть в процессе эволюции.
Что следует понимать под эволюцией? Вальтер Циммерман (1953) дал следующее определение понятия «эволюция»: «Преобразование формы и образа жизни организмов, в результате чего потомки приобретают другие черты, отличающие их от предков». Это определение фактически показывает конечный результат процесса эволюции, но не раскрывает причины этого явления. Более детальное определение принадлежит советскому зоологу С. С. Шварцу (1974): «Эволюция — это историческое развитие живых существ, которое сопровождается прогрессирующим совершенствованием морфофизиологической организации. Эволюция — это в конечном счете процесс приспособления к среде». Эта последняя фраза подчеркивает и раскрывает причину тех преобразований, которые происходили и у обитателей водной среды, и у растений, вышедших на сушу.
Согласно существующим представлениям, эукариотические организмы появились около 1—1,5 млрд лет тому назад в водной среде. Предполагают, что они были шаровидными и питались гетеротрофно, всасывая питательные вещества всей поверхностью тела. Среди современных одноклеточных водорослей мож-
но встретить немало представителей, имеющих шаровидную форму тела (хлорелла, эремосфера, голенкиния и др.). Постепенно форма тела водорослей под влиянием условий существования изменялась. Поскольку всасывание пищи из внешней среды осуществлялось всей поверхностью тела, в лучшем положении оказывались те организмы, у которых поверхность тела была большей.
Таким образом, способ питания надо рассматривать как первый движущий фактор эволюции, который обусловил изменение формы тела древних водорослей. Основной принцип эволюции формы тела растений состоял в увеличении их поверхности. Так как подавляющее большинство растений — автотрофные организмы, эволюция на завершающем этапе обеспечила увеличение фотосинтезирующей поверхности растения.
На первых этапах эволюции шаровидных организмов, очевидно, происходило увеличение поверхности, но сохранялось геометрическое подобие тела, т. е. организмы оставались шаровидными. Однако шаровидные организмы не могли увеличиваться до бесконечности. При сохранении геометрического подобия объем и поверхность шаровидных организмов растут непропорционально. Поверхность тела увеличивается в арифметической прогрессии, а объем тела — в геометрической. Это приводит к нарушению обменных реакций между одноклеточным организмом и средой, создаются неблагоприятные условия для существования организма, что может повлечь за собой его гибель. Чтобы избежать гибели и успешно развиваться, шаровидное тело растений должно было каким-то образом измениться. На основании анализа строения тела у современных водорослей можно предположить, что в процессе приспособительной эволюции объем тела и его поверхность могли прийти в соответствие двумя путями. У одних организмов увеличение объема тела привело к изменению шаровидной формы тела на иную. У других — увеличение поверхности тела было сопряжено с интенсивным делением ядра без последующего цитокинеза. Так возникли одноклеточные многоядерные организмы, которые по внешнему виду, строению и размерам существенно отличаются от типичных клеток и называются сифональными (ботридиум, каулерпа). Однако эволюция не пошла по пути усложнения и дальнейшего развития сифональных форм. Эта ветвь развития является «слепой». В противовес второму первое направление оказалось весьма прогрессивным и постепенно привело к появлению форм, имеющих при том же объеме большую поверхность тела, чем у шара. Так возникли цилиндрические, палочковидные, серповидные, эллиптические и другие одноклеточные организмы, которые существуют среди водорослей и по сей день.
От одноклеточных, предположительно, возникли и многоклеточные организмы. Роль промежуточного звена между ними сыграли колониальные формы — водные организмы, у которых особи дочерних поколений при бесполом размножении остаются соединенными с материнским организмом и образуют более или менее сложное объединение. Колонии бывают свободно плавающими и прикрепленными. Плавающие колонии имеют шаровидную форму. Прикрепленные колониальные водоросли могут быть нитчатыми, мешковидными, мешковидно-пластинчатыми, древовидными. В примитивных колониях
клетки равномерно расположены в толще объединяющей их слизи и выполняют одинаковые функции. Преимущество колониальных форм по сравнению с одноклеточными организмами состоит в том, что клетки в колонии могут максимально использовать питательные вещества или продукты жизнедеятельности других клеток, главным образом кислород и углекислый газ.
По мере изменения условий существования усложнялась структура колониальных форм: эволюция не только шла по пути изменения внешнего вида колонии, но и была связана с дифференцировкой функций клеток, входящих в ее состав. Наряду со свободно плавающими колониями появляются прикрепленные. В таких колониях способность к делению сохраняется только у некоторых верхушечных клеток, а базальная клетка выполняет функцию прикрепления к субстрату. Если верхушечная клетка делится только в одном направлении, колония приобретает нитчатую форму (улотрикс). Если деление идет в двух направлениях (в длину и ширину), возникают однослойные или двуслойные пластинчатые формы (ульва, энтероморфа). При трехмерном направлении деления формируются многослойные пластинчатые организмы (фукус, ламинария).
Несмотря на сложное внешнее строение, многоклеточные водоросли имеют довольно примитивную внутреннюю организацию. У них нет типичных тканей. Тело растения ветвится дихотомически (от греч. dicha — на две части, tome — разрез, сечение), оно не расчленено на вегетативные органы. Репродуктивные органы (спорангии, гаметангии) у подавляющего большинства водорослей одноклеточные (очень редко гаметангии многокамерные). Нерасчлененное на вегетативные органы тело растений называется слоевищем, или талломом.
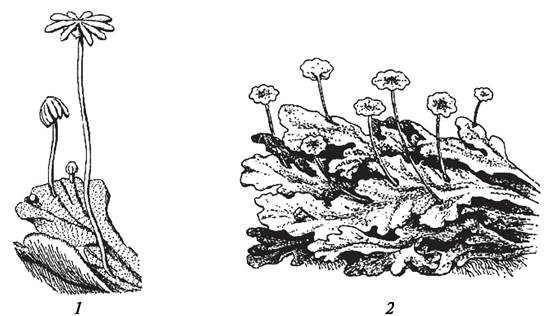
Талломные организмы встречаются и среди примитивных высших растений (некоторые печеночные мхи), однако их немного и эволюционная роль их не так велика (рис. 52).
Рис. 52. Печеночный мох маршанция (Marchantía polymorpha): 1—таллом с женскими подставками; 2—таллом с мужскими подставками
4.2. Реконструированная схема строения псилофитов (риниофитов)
Вершиной эволюционного процесса явилось утверждение на Земле высших растений с присущей большинству из них листостебельной организацией. Возникновение высших растений было результатом выхода на сушу водорослевых предков и их приспособления к условиям существования в воздушной среде.
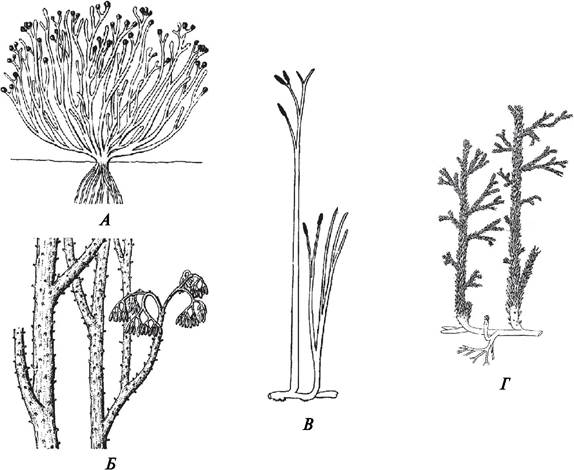
Первыми достоверно известными представителями наземных растений являются риниофиты (псилофиты). Организация риниофитов весьма примитивная. Многие из них (куксония, риния) были очень похожи на водоросли, имели небольшие размеры (от 20 до 100 см), ветвились дихотомически, у них отсутствовала дифференциация на листья и стебли (рис. 53). В то же время у этих растений были явные признаки, отличавшие их от водорослей. У первопоселенцев суши появились некоторые ткани: покровная, с хорошо выраженными устьицами, паренхимная и, что особенно важно, проводящие ткани — ксилема и флоэма.
Рис. 53. Первопоселенцы суши: А—куксония; Б—псилофит; В—риния; Г—астероксилон
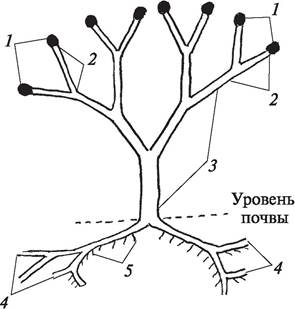
Первые четкие представления о строении первопоселенцев суши дал основоположник теломной теории В. Циммерман в своей книге «Филогения растений» (1930). Поскольку вымершие растения чаще всего обнаруживаются в виде разрозненных остатков, он предложил реконструированную схему строения псилофитов. Согласно представлениям, В. Циммермана, тело псилофитов еще не было расчленено на органы и состояло из осевых дихотомически разветвленных надземных и подземных конструктивных элементов. Растения прикреплялись к субстрату дихотомически разветвленными и погруженными в почвуризомоидами (от греч. rhiza — корень и eidos — вид). На ризомоидах формировались ризоиды, при помощи которых в растения из почвы поступала вода с растворенными в ней питательными веществами. Надземная часть растения состояла из мезомов (от греч. mesos — средний) и теломов (от греч. telos — конец), конечных «веточек», на концах которых располагались спорангии. Мезомы осуществляли функции фотосинтеза и проведения питательных веществ и располагались между теломами и ризомоидами (рис. 54).
Выход растений на сушу сопровождался выработкой сложной системы приспособлений к новым условиям жизни, которые существенно изменили не только их внешний облик, но и внутреннее строение.
Рис. 54. Схема строения псилофита: 1—спорангии; 2—теломы; 3—мезомы; 4—ризомоиды; 5—ризоиды
В процессе эволюции из конструктивных элементов первопоселенцев суши возникли вегетативные органы высших растений. Ризомоиды дали начало настоящим корням, мезомы превратились в стебли и боковые ветви растений, теломы или сохранили свою функцию как спороносные образования, или могли дать начало листьям. Таким образом, в процессе длительной эволюции появились листостебельные растения, хорошо приспособленные к жизни на суше. Движущими факторами, которые привели к изменению формы тела высших растений, несомненно, явились особенности питания (почвенное и воздушное) и влияние условий существования. Основным принципом эволюции тела высших растений было увеличение фотосинтезирующей поверхности.