МОРФОЛОГИЯ РАСТЕНИЙ - Т. А. Сауткина - 2012
ГЛАВА 6. РАЗМНОЖЕНИЕ РАСТЕНИЙ
6.1. Понятие о размножении. Принципы классификации размножения растений
Растительные организмы независимо от уровня их структурной организации представляют собой сложные многофункциональные системы, способные не только поддерживать жизнь каждой отдельной особи, но и жизнь вида в целом.
Одной из важнейших функций любого живого организма является воспроизведение себе подобных (функция самовоспроизведения). Самовоспроизведение может осуществляться на разных уровнях структурной организации. Увеличение количества нуклеиновых кислот в ядрах клеток перед митотическим или мейотическим делениями — это воспроизведение, которое осуществляется на молекулярном уровне. Образование новых клеток при митозе — это самовоспроизведение на клеточном уровне. И, наконец, воспроизведение может осуществляться на уровне организмов. Это и есть размножение.
Размножение, т. е. способность оставлять после себя потомство, — одно из основных свойств всех живых организмов. Оно поддерживает непрерывность и преемственность жизни видов, и вообще жизни на Земле.
Для растений, большинство из которых ведет прикрепленный образ жизни, размножение имеет еще одно важное значение: оно способствует распространению растений, что приводит к расширению ареала — площади, занимаемой видом.
Обеспечивая в целом существование вида, размножение в то же время не поддерживает жизнь самой размножающейся особи, а иногда приводит даже к ее гибели. Погибают, но оставляют после себя потомство одноклеточные водоросли, однолетние и двулетние цветковые (покрытосеменные) растения, а также многолетние монокарпические виды — цветущие и плодоносящие один раз в жизни (алоэ, агавы, бамбуки, ананасы).
Размножение растений — очень сложный биологический процесс, прежде всего генетический, связанный с передачей наследственной информации от родительских организмов дочерним. Размножение — это и физиолого-биохимический процесс, так как он сопровождается рядом сложных биохимических реакций, связанных с накоплением, перераспределением таких важнейших соединений, как нуклеиновые кислоты (ДНК и РНК), белки, каротиноиды, фитогормоны и др. Наконец, размножение — это процесс эмбриологический, ведь у большинства растений зачаток нового организма формируется в тканях организма материнского.
К сожалению, приходится констатировать, что до настоящего времени в различных учебниках, учебных пособиях, справочных изданиях, словарях понятие «размножение растений» трактуется неоднозначно. В то время как большинство авторов рассматривают размножение как воспроизведение себе подобных, некоторые противопоставляют понятия «воспроизведение» и «размножение» и считают, что размножением можно назвать только такой процесс, при котором происходит увеличение численности особей вида.
Вместе с тем при определении понятия «размножение растений» не столь важна количественная сторона процесса, сколь качественная — появление новых особей вида, что обеспечивает преемственность и поддержание жизни вида в целом. Исходя из этого, дать определение понятия «размножение растений» можно следующим образом: Размножение растений — сложный биологический процесс воспроизведения на уровне организмов, который завершается образованием особей вида, подобных родительским, обеспечивает поддержание жизни вида и его расселение.
Это определение позволяет подчеркнуть сущность процесса размножения — воспроизведение себе подобных, т. е. образование тех морфофизиологических структур (особей вида), которые ботаники представляют, как биологический вид. Особь — это наименьшая единица биологического вида, подверженная действию факторов эволюции.
Эффективность любого способа размножения теоретически можно охарактеризовать коэффициентом размножения (K—1) и коэффициентом расселения (K—2).
K - 1 = (а : б) • 100 %,
где а — количество особей, образовавшихся при размножении; б — количество особей, которое теоретически могло бы образоваться.
K - 2 = (с : d) • 100 %,
где с — сумма расстояний, на которые удалились дочерние особи от материнской; d — сумма теоретически возможных расстояний, на которые могли бы удалиться при размножении дочерние особи.
Фактически эти показатели в природных условиях высчитать почти невозможно, а, следовательно, охарактеризовать эффективность того или иного способа размножения можно только гипотетически (приблизительно).
Неоднозначность понимания сущности процесса размножения и употребления терминов «размножение» и «воспроизведение» породила соответствующий подход и к классификации типов размножения растений, и это не удивительно. Исторически сложилось так, что многие открытия (особенно это касается изучения клетки) ботаники делали после зоологов и автоматически переносили термины, введенные зоологами, на ботанические объекты. Так, очевидно, случилось и с представлением о размножении растений. Многие ботаники упрощенно подходят к пониманию процесса размножения растений, сопоставляют его с таковым у животных и исходя из этого классифицируют типы размножения растений приблизительно так же, как и животных. Чаще всего в учебниках выделяют три типа размножения растений: вегетативное, бесполое и половое. При этом не учитывается, что процессы индивидуального развития (онтогенез) и особенности циклов развития (жизненных циклов) у растений специфичны, очень сложны и протекают не так, как у животных. Если у животных процесс онтогенеза и жизненный цикл, как правило, совпадают, то у споровых и многолетних семенных растений в течение онтогенеза может проходить несколько жизненных циклов.
Очевидно, что сложность в выработке единой классификации и выделения типов размножения растительных организмов объясняется чрезвычайным разнообразием структурной организации растений (от одноклеточных водорослей до победителей в борьбе за существование — покрытосеменных) и, как следствие этого, очень сложными и разнообразными циклами развития.
На основании особенностей жизненных циклов у водорослей и высших растений (мохообразные, плаунообразные, хвощеобразные, папоротникообразные, голосеменные и покрытосеменные) мы считаем возможным выделить пять типов размножения растений: бесполое, половое, гаметоспоровое, спорогаметное и семенное. В этой классификации учитывается характер репродуктивных процессов, происходящих в жизненных циклах растений, конечным результатом которых является образование себе подобных, т. е. образование потомства, особей вида.
6.1.1. Бесполое размножение
Бесполое размножение осуществляется или частями тела растения, или специализированными гаплоидными клетками — спорами, которые образуются в спорангиях в результате митотического деления. Такие споры называются митоспорами. С учетом особенностей репродуктивных элементов бесполое размножение можно подразделить на вегетативное размножение и собственно бесполое (агамное, или спорообразование) размножение.
Вегетативное размножение. При вегетативном размножении дочерние организмы образуются из участков слоевища, фрагментов типичных или метаморфизированных вегетативных органов, а также специализированных образований — выводковых почек, которые формируются у некоторых плаунообразных и папоротникообразных.
Вегетативное размножение — это универсальный способ размножения. Он присущ как водорослям, так и высшим растениям. Вегетативное размножение происходит в природе самопроизвольно, а также широко используется в практической деятельности человека, поэтому его можно подразделить на естественное и искусственное.
В основе вегетативного размножения лежит свойство регенерации, т. е. способность восстанавливать недостающие вегетативные органы растения и формировать новые дочерние организмы. Так как при вегетативном размножении от родительского организма отделяются какие-либо части (одноклеточные или многоклеточные), сформировавшиеся дочерние особи являются точной копией родительской особи. Следовательно, при вегетативном размножении консервативно закрепляются как генотипические, так и фенотипические особенности вида.
Естественное вегетативное размножение — это процесс, который происходит в природе без вмешательства человека.
У водорослей естественное вегетативное размножение происходит разными способами. Самыми примитивными способами вегетативного размножения у этих организмов считаются деление клетки надвое, что характерно для амебоидных, монадных и коккоидных форм, а также фрагментация, которая наблюдается у колониальных и многоклеточных, нитчатых и пластинчатых водорослей. Причиной распада многоклеточного слоевища на фрагменты часто являются случайные механические факторы (действие волн, сильные течения, повреждения животными) или естественное отмирание части тела. Иногда для вегетативного размножения служат специальные образования. У некоторых сифоновых, бурых и красных водорослей образуются специальные органы вегетативного размножения — выводковые почки — фактически миниатюрные дочерние особи. Харовые водоросли образуют клубеньки, из которых после зимы вырастают новые особи.
У высших растений способы естественного вегетативного размножения более разнообразны и специализированы. У споровых растений (мхи, плауны, хвощи, папоротники) дочерние организмы могут развиваться из участков типичных стеблей (плауны), корневищ (хвощи, папоротники) или особых почек — выводковых и адвентивных (придаточных). Выводковые почки образуются на поверхности слоевища в особых выводковых корзинках (печеночный мох маршанция) либо на верхушке стебля (некоторые зеленые мхи). Адвентивные почки характерны для многих тропических папоротников, а также для растений тундровой зоны (баранец обыкновенный — Huperzia selago). У папоротников они чаще всего возникают налистьях (асплениум — Asplenium, цистоптерис — Cystopteris), прилистниках (мараттия — Marattia), реже на корнях (ужовник — Ophioglossum).
У голосеменных способность к вегетативному размножению выражена слабо. В естественных условиях могут размножаться вегетативно кедровый стланик (Pinuspumila), сосна горная (Pinus montana), можжевельник казацкий (Juniperus sabina). У этих растений ветки, соприкасающиеся с почвой, могут укореняться и формировать так называемые отводки. У некоторых видов саговниковых (саговник — Cycas, цератозамия — Ceratozamia) изредка наблюдается образование у основания ствола особых почек, из которых могут развиться дочерние растения.
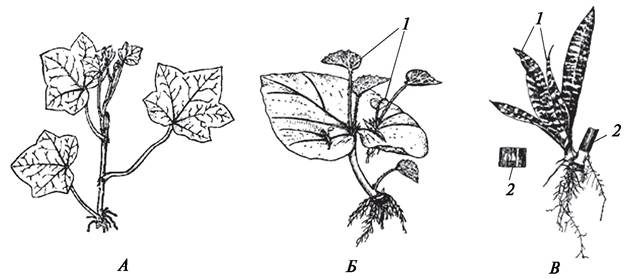
Наиболее разнообразными и специализированными формами вегетативного размножения характеризуются покрытосеменные растения. В естественных условиях вегетативное размножение у покрытосеменных растений осуществляется различными частями. Часто встречается размножение корневыми отпрысками (малина, вишня, ивы, сирень и др.), которое происходит благодаря заложению и развитию придаточных почек на корнях. Сравнительно широко распространено размножение стеблевыми отводками (смородина, калина, липа и др.). Иногда встречается также размножение при помощи черенков (ива ломкая). Черенок — это часть (отрезок) любого вегетативного органа. Черенки могут быть стеблевыми, листовыми и корневыми (рис. 131). На черенках в ответ на механическое повреждение образуются раневые меристемы, благодаря работе которых закладываются почки и придаточные корни и начинают формироваться новые особи вида. Иногда почки могут закладываться на не отделенных от материнского растения вегетативных органах.
Рис. 131. Вегетативное размножение растений: А—укорененный черенок плюща обыкновенного (Hedera helix); Б—укорененный лист бегонии королевской (Begonia rex); В —укорененная часть листа сансевьеры трехполосой (Sansevieria trifasciata): 1—образовавшиеся молодые растения; 2—часть листа сансевьеры
Вегетативное размножение осуществляют также специализированные органы: столоны — подземные (седмичник) и надземные (земляника), корневища (пырей, сныть), клубни (топинамбур, чистец, картофель), луковицы (тюльпаны, гиацинты, нарциссы), корневые шишки (корнеклубни — чистяк, георгина).
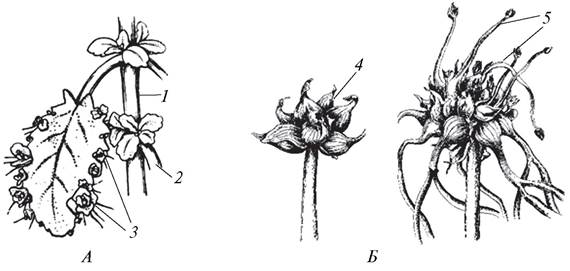
Особым типом естественного вегетативного размножения является вивипария, или живорождение. Различают ложную и истинную вивипарию. При ложной вивипарии на листьях (бриофиллюм — Bryophyllum, сердечник луговой — Cardamine pratensis), побегах (баранец обыкновенный — Huperzia selago) или в соцветиях (мятлик живородящий — Poa vivipara,лук огородный — Allium oleraceum) формируются придаточные почки, из которых уже на растении образуются миниатюрные дочерние особи, имеющие все вегетативные органы (рис. 132). При отделении от материнского растения такие «детки» легко укореняются и быстро развиваются во взрослые особи.
Рис. 132. Вивипария ложная: А—каланхое перистое (Kalanchoe pinnatum); Б—лук репчатый (Allium cepa var. vivipara); 1—главный побег; 2—боковой побег; 3—«детки», развившиеся из адвентивных почек; 4—формирование луковичек-деток в соцветии; 5—прорастание луковичек-деток в соцветии
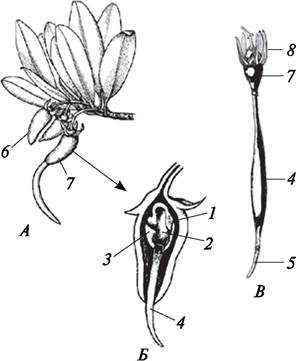
При истинной вивипарии формируются семена, которые прорастают и образуют молодые дочерние особи, находясь еще на материнском растении. Истинная вивипария характерна для представителей мангровых деревьев (виды рода ризофора — Rhizophora mucronata. R. mangle и др.), обитающих на береговых отмелях океанов (рис. 133).
Рис. 133. Истинная вивипария: А—ветвь ризофоры (Rhizophora mangle) со зрелыми плодами и прорастающим семенем; Б—продольный разрез прорастающего семени, находящегося в плоде; В—проросток ризофоры: 1—кожура семени; 2—семядоля; 3—эндосперм; 4—гипокотиль; 5—зародышевый корень; 6—зрелый плод; 7— прорастающее семя, находящееся в плоде; 8—остатки околоцветника
Поскольку при вегетативном размножении дочерние организмы являются точной копией материнского организма, эта особенность нашла широкое применение при искусственном вегетативном размножении.
Искусственное вегетативное размножение — это размножение, которое человек использует в своей практической деятельности. Этот вид вегетативного размножения широко используется в тех случаях, когда растение не образует семян (многие сорта картофеля), образует семена низкого качества (мята перечная — Mentha piperita), если растение имеет гибридное происхождение и при размножении семенами дает расщепляющееся потомство (сорта гвоздики садовой — Dianthus caryophyllus). Искусственное вегетативное размножение — это часто единственный способ размножения, позволяющий сохранить генетическую чистоту сорта (картофель — Solanum tuberosum, земляника садовая — Fragaria magna).
Способы искусственного вегетативного размножения те же, что и естественного. В культуре многие растения размножают корневищами (ирис — Iris, мята перечная — Mentha piperita), луковицами (тюльпаны — Tulipa, нарциссы — Narcissus), клубнями (картофель, топинамбур — Helianthus tuberosus), клубнелуковицами (гладиолусы — Gladiolus, крокусы — Crocus). Широко используется размножение с помощью различных по происхождению черенков (стеблевых — зеленых и одревесневших; листовых; корневых) (см. рис. 131).
Совокупность новых особей, полученных от одного материнского растения вегетативным путем, называется клоном. В связи с промышленным развитием биотехнологии широкое распространение получил метод микроклонирования растений в культуре in vitro («в стекле»), т. е. вне живого организма. При этом используются различные питательные среды, содержащие фитогормоны. Существует два способа микроклонирования: новые организмы можно получить из клеток апикальных меристем и из эксплантатов — клеток постоянных тканей, выделенных из различных органов растений. Микроклонирование из апексов позволяет получить очень большое количество идеально однородного посадочного материала (регенерантов), освобожденного от грибной, бактериальной и, главное, вирусной инфекции. Такой метод применяют для размножения и оздоровления многих декоративных растений (гвоздика — Dianthus, хризантемы — Chrysanthemum), а также сельскохозяйственных растений (земляника садовая, картофель). При получении регенерантов из эксплантатов наблюдается широкий размах изменчивости (соматоклональная изменчивость) признаков, поэтому такой способ микроклонирования используется в селекции как дополнительный источник изменчивости организмов.
Особым типом искусственного вегетативного размножения являются прививки, спонтанное образование которых в природе происходит крайне редко. При прививке человек хирургическим путем отделяет часть одного растения (черенок, почку) — привой и прививает его на другое растение — подвой. Прививку почкой («глазком») чаще всего производят летом, а черенком — весной. Широко распространен метод размножения растений с помощью прививки в промышленном, а также в декоративном садоводстве. Так размножают различные сорта яблони, груши, вишни, розы, сирени и др.
Вегетативное размножение — самый древний способ размножения. Оно возникло, как полагает М. М. Лодкина (1965), на доклеточном уровне жизни как ответная реакция организмов на нарушение соотношения процессов роста и питания.
Коэффициент вегетативного размножения у различных растений отличается, но он относительно невелик, хотя у некоторых растений (мускари — Muscari, птицемлечник — Ornithogalum) за один вегетационный период может сформироваться до 30 луковичек-деток. Коэффициент расселения у вегетативно размножающихся видов низок. Дочерние организмы концентрируются вокруг материнских особей или располагаются вблизи них. Благодаря этому многие растения способны быстро формировать крупные группировки (живучка ползучая — Аjugа reptans, лапчатка гусиная — Potentilla аnseriа, пырей ползучий — Elytrigia гереns).
Собственно, бесполое размножение (агамное). Собственно, бесполое размножение происходит в тех случаях, когда из особых специализированных клеток (спор), как правило, покидающих материнский организм, развиваются особи, воспроизводящие морфофизиологические признаки родителя. Такой тип размножения характерен для примитивных талломных организмов: одноклеточных и нитчатых зеленых водорослей (хламидомонада, хлорелла, улотрикс, эдогониум), а также эвгленовых, рафидофитовых, криптофитовых, золотистых, многих желто-зеленых (трибонема) и некоторых бурых водорослей. Споры, обеспечивающие воспроизведение родительских признаков, образуются в результате митотического деления, их называют митоспорами. В подавляющем большинстве случаев митоспоры формируются в клетках гаплоидных талломов и так же, как они, гаплоидны.
Реже митоспоры образуются на диплоидном талломе, в этом случае они будут диплоидными, и из них тоже разовьется диплоидное растение, подобное материнскому организму (например, кладофора из зеленых, эктокарпус из бурых водорослей).
У большинства водорослей образуются подвижные споры со жгутиками — зооспоры. Зооспоры могут различаться по строению, по числу и длине жгутиков. Подвижные зооспоры обычно не имеют настоящей твердой оболочки и окружены нежным перипластом.
У некоторых водорослей споры неподвижны (апланоспоры). Они одеваются оболочкой еще внутри материнской клетки (хлорококковые из зеленых водорослей, многие желто-зеленые, бурые, красные водоросли), и в ряде случаев уже подобны родительскому организму (например, автоспоры хлореллы). Зооспоры (споры) образуются или в клетках, не отличающихся по форме от вегетативных, или в особых одноклеточных репродуктивных органах — зооспорангиях (спорангиях). В одной клетке или спорангии их может быть от одной (эдогониум из отдела Зеленые водоросли) до нескольких сотен (кладофора из того же отдела). Чаще всего споры микроскопически малы и образуются в больших количествах. Обычно они выходят в воду через отверстие в стенке спорангия целой группой, окруженные общей слизистой оболочкой. Вскоре оболочка разрушается, а зооспоры моментально расплываются в разные стороны. Осев на субстрат, они покрываются твердой оболочкой и прорастают, образуя новый организм водоросли того же вида.
Таким образом, митоспоры осуществляют процесс собственно бесполого размножения и способствуют расселению вида.
Собственно, бесполое размножение по сравнению с вегетативным имеет более высокие коэффициенты размножения и расселения. По мнению М. М. Лодкиной, спорообразование как способ размножения растений возникло на клеточном уровне организации как ответная реакция организма на условия, существенно затрудняющие основные жизненные процессы и прежде всего рост.
Процесс спорообразования характерен для подавляющего большинства водорослей (исключение составляют диатомовые водоросли —Bacillariophyta, харовые — Charophyta, бурые из класса Cydosporophyceae) и всех без исключения высших растений.
Кроме митоспор у растений есть еще и мейоспоры. Они также образуются в спорангиях, но отличаются от митоспор по особенностям образования и строения. Так как мейоспоры развиваются только на спорофитах, образованию мейоспор всегда предшествует мейоз (редукционное деление). Образование мейоспор характерно для многих водорослей и всех без исключения высших растений. Как и типичные митоспоры, мейоспоры — это специализированные гаплоидные клетки, но строение их и функции, особенно у высших растений, имеют свои особенности. Мейоспоры у водорослей могут быть представлены зооспорами (ульва), реже апланоспорами (улотрикс при прорастании зиготы). У высших споровых растений (плаунов (рис. 134), хвощей (рис. 135), папоротников (рис. 136), в отличие от водорослей мейоспоры всегда неподвижны, одеты двумя оболочками — экзоспорием и эндоспорием. Экзоспорий образован целлюлозой и пропитан спорополленином, который повышает прочность оболочки. В экзоспории имеется «щель разверзания», через которую происходит прорастание споры. У высших споровых растений мейоспоры при попадании на субстрат прорастают и образуют гаметофит (заросток) (см. рис. 134, 135, 136), который по морфологическим, цитологическим и функциональным особенностям отличается от спорофита, в спорангиях которого образовались споры.
Таким образом, бытующее и широко распространенное утверждение, что у высших споровых растений (плаунов, хвощей, папоротников) «размножение происходит с помощью спор» в корне не верно.
Рис. 134. Спорофит, спорангий, споры и гаметофит плауна булавовидного (Lycopodium clavatum) (А—Г) и гаметофит дифазиаструма сплюснутого (Diphasiastrum complanatum) (Д): А—внешний вид спорофита со спороносными колосками; Б—спорангий на спорофилле; В—споры с проксимального и дистального полюсов; Г, Д— строение гаметофитов: 1—антеридии; 2—архегонии; 3—гифы гриба; 4—ризоиды
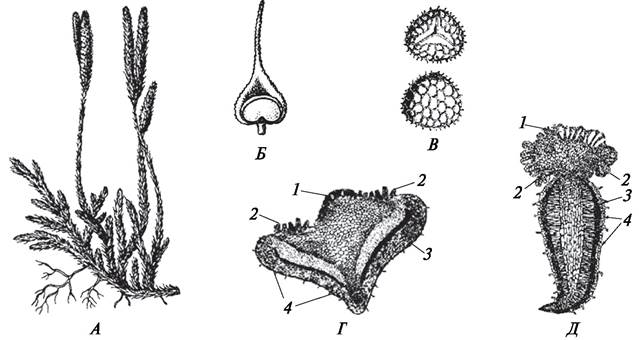
Рис. 135. Спорофит, спорангиофор, спора, гаметофиты хвоща полевого (Equisetum агvеnsе): А—внешний вид весеннего и летнего побегов спорофита; Б—спорангиофор со спорангиями; В—спора с элатерами; Г—обоеполый гаметофит; Д—женский гаметофит
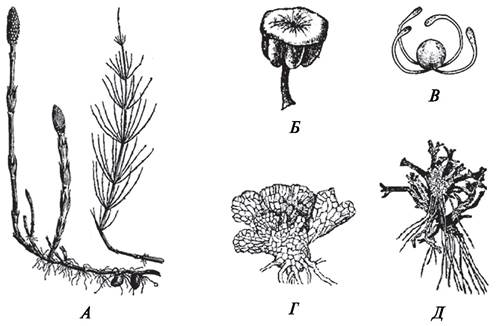
Рис. 136. Спорофит, спорангий, споры, гаметофит щитовника мужского (Dryopteris filix-mas): А—внешний вид спорофита; Б—спорангий со спорами; В—споры (вид с проксимального полюса и с экватора); Г—обоеполый гаметофит
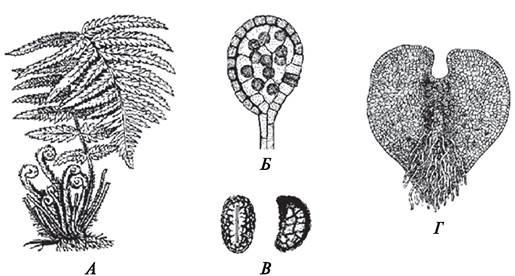
У высших споровых растений спора не осуществляет процесс размножения, понимаемый как воспроизведение себе подобных. Являясь составной частью процесса размножения, она выполняет две функции: способствует расселению вида (на образовавшемся из споры гаметофите впоследствии образуется зародыш нового организма) и благодаря наличию прочной наружной оболочки помогает пережить неблагоприятные для развития вида условия.
Среди высших растений существуют равноспоровые (явление изоспории) и разноспоровые (явление гетероспории) виды. У равноспоровых растений все споры одинаковы по морфологическим и физиолого-биохимическим свойствам. Из таких спор образуются обоеполые заростки. Они существуют совершенно независимо от спорофитов и по типу питания могут быть как автотрофными (фотосинтезирующими — у настоящих папоротников, хвощей), так и микотрофными (у ужовниковых папоротников, плаунов).
Разноспоровые растения появились в процессе эволюции наряду с равноспоровыми. Первоначально возникла физиологическая гетероспория, при этом не различающиеся по морфологическим признакам споры при попадании на субстраты разной трофности формировали раздельнополые заростки. Экспериментально доказано, что у хвощей на бедных питательными веществами почвах образуются мужские заростки, а на богатых — женские. Впоследствии разноспоровость стала морфологической. У разноспоровых растений стали развиваться спорангии двух типов: микроспорангии и макроспорангии (мегаспорангии). В микроспорангиях образуются многочисленные мелкие споры, получившие названия «микроспоры», в макроспорангиях — малочисленные относительно более крупные споры — «макроспоры» (мегаспоры). При дальнейшем развитии из микроспор образуются мужские заростки, а из макроспор — женские.
Разноспоровость — первая ступенька на пути половой дифференцировки особей. У разноспоровых плаунообразных (Salviniа), папоротникообразных (Salviniа) функцию расселения вида осуществляет только макроспора. Именно из макроспоры образуется женский гаметофит, на котором формируются архегонии с развивающейся в них яйцеклеткой, происходит процесс оплодотворения, и на завершающем этапе появляется зародыш нового спорофита. Мужской заросток, который развивается из микроспоры, после образования в антеридиях сперматозоидов погибает.
У голосеменных и покрытосеменных растений, для которых характерна морфологическая гетероспория, ни макроспоры, ни микроспоры никогда не покидают материнского растения. У представителей этих отделов высших растений макроспора утратила функцию расселения, но процесс спорообразования у них происходит всегда и имеет большое биологическое значение. Он обязательно предшествует половому процессу.
У всех без исключения высших растений и у высокоорганизованных водорослей размножение происходит только при сочетании двух процессов — спорообразования и полового процесса.