МОРФОЛОГИЯ РАСТЕНИЙ - Т. А. Сауткина - 2012
ГЛАВА 6. РАЗМНОЖЕНИЕ РАСТЕНИЙ
6.3. Понятие о жизненных циклах и чередовании поколений у растений
Понятие о чередовании поколений возникло благодаря работе польского ботаника М. Лещик-Суминского (1820—1898), установившего в цикле развития папоротников наличие двух фаз — спороносной и половой. На заростке папоротника, который, по его мнению, является промежуточным, но вполне самостоятельным образованием между спорой и молодым растением папоротника, он нашел половые органы — антеридии со сперматозоидами и архегонии с яйцеклеткой. Это было важным научным открытием. Его книга «К истории развития папоротников» вышла в Берлине в 1848 г.
В. Гофмейстер (1824—1877) — немецкий ботаник, профессор Гейдельбергского, а затем Тюбингенского университетов высоко оценил открытие М. Лещик-Суминского. Вдохновленный широкими научными перспективами, открываемыми его работой, он принялся за сравнительно-эмбриологические исследования архегониальных растений.
В 1849 г. вышла первая работа В. Гофмейстера, в которой он ввел понятие о чередовании поколений у растений. Впоследствии В. Гофмейстер сделал величайшее открытие: он пришел к выводу, что у мохообразных, папоротникообразных и голосеменных происходит правильная смена (чередование) двух поколений — полового и бесполого. Оба эти поколения отличаются друг от друга формой, способом питания, биологическими и морфологическими особенностями. Свои представления об индивидуальном развитии изученных таксонов он изложил в капитальном труде «Сравнительное исследование над прорастанием, развитием и плодообразованием высших споровых растений и образованием семян у хвойных», опубликованном в 1851 г. в Лейпциге.
В ряде работ, вышедших в начале ХХ в. (1911, 1927), профессор Московского университета Л. И. Курсанов описал чередование поколений у зеленых и бурых водорослей.
Итак, жизненный цикл — это свойственная различным таксонам последовательность развития от какого-нибудь этапа до его повторения. Например, у семенных растений жизненный цикл продолжается от семени до семени; у плаунообразных и других споровых растений — от споры до споры. В жизненном цикле высших растений может доминировать или гаметофит (только у мохообразных), или спорофит (у всех высших споровых растений, а также у голосеменных и покрытосеменных).
Схема жизненного цикла, характерная для равноспоровых папоротников, представлена на рис. 138. Из схемы видно, что в жизненном цикле процесс спорообразования сменяется половым процессом, и как следствие этого, бесполое поколение (спорофит) сменяется половым (гаметофитом).
Рис. 138. Схема жизненного цикла у равноспоровых папоротников (геторофазное гетероморфное чередование поколений)
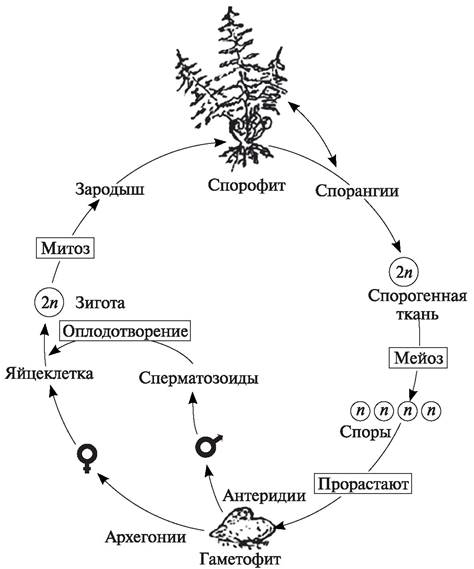
У плаунообразных, хвощеобразных и папоротникообразных спорофит и гаметофит существуют независимо друг от друга, т. е. разделены пространственно и представляют собой два поколения одного и того же вида.
У мохообразных доминирует гаметофит. Спорофит развивается на гаметофите и частично питается за его счет. У голосеменных и покрытосеменных растений, для которых характерна разноспоровость, доминирует спорофит. Гаметофиты формируются на материнских организмах, развиваются за счет спорофита, сильно редуцированы и никогда не покидают материнского растения.
Таким образом, под поколением следует понимать совокупность органических форм, возникающих в результате какого-либо репродуктивного процесса и генетически связанных с предками и потомками. Явление, связанное со сменой репродуктивных процессов и приводящее к образованию в жизненном цикле форм, различающихся морфологически, цитологически и функционально, называется чередованием поколений, или сменой фаз развития.
6.3.1. Общие закономерности чередования поколений (смена фаз развития)
Морфологические особенности спорофитов и гаметофитов у представителей различных таксономических групп, их взаимная связь друг с другом, а также закономерности смены фаз развития (чередование поколений) различны.
Чередование поколений бывает факультативным и облигатным.
У примитивных водорослей чередование поколений носит факультативный характер и чаще всего связано со сменой экологических факторов (понижением температуры воды, подсыханием водоема и т. п.). Так, например, пресноводная нитчатая водоросль улотрикс (Ulothrix) может многократно формировать 4-жгутиковые зооспоры, которые, проплавав некоторое время в воде, оседают на дно, утрачивают жгутики и прорастают в новые гетероталличные нити улотрикса. Осенью в клетках (одноклеточных гаметангиях) улотрикса формируются изогаметы и в результате процесса оплодотворения образуется зигота. Она «одевается» прочной оболочкой, оседает на дно, где и сохраняется до весны.
Схематично факультативное чередование поколений у улотрикса выглядит так: n... n.... n... 2n... n... n и т. д.
Однако у подавляющего большинства водорослей и у всех без исключения высших растений в жизненном цикле наблюдается строго закономерная смена спорообразования и полового процесса и происходит облигатное (обязательное) чередование поколений. Схематично его можно представить так: n... 2n... n... 2n... n... 2n, если в жизненном цикле доминирует гаметофит; или 2n... n... 2n... n... 2n... n, если в жизненном цикле доминирует спорофит.
Переход от спорофита к гаметофиту происходит в результате редукционного деления (мейоза) спороцитов (материнских клеток спор), которое происходит в спорангиях и предшествует образованию спор. Из спор развивается гаметофит, на котором формируются половые клетки — гаметы. Переход от гаметофита к спорофиту — следствие процесса оплодотворения яйцеклетки сперматозоидом (у высших споровых растений) или спермием (у высокоорганизованных голосеменных и покрытосеменных растений), в результате чего образуется зигота, восстанавливается диплоидный набор хромосом и начинает из зиготы формироваться спорофит.
Чередование поколений может быть гомофазным и гетерофазным. Эти термины введены П. Хартманом в 1939 г.
При гомофазном чередовании поколений спорообразование и половой процесс происходят у примитивных гаплоидных водорослей при одной и той же ядерной фазе, иначе говоря, смена репродуктивных процессов не влечет за собой изменений ядерной фазы. Например, зооспоры и изогаметы у улотрикса в различные периоды формируются в результате митоза на одних и тех же гетероталличных гаплоидных нитях (рис. 139).
Рис. 139. Схема гомофазного чередования поколений у улотрикса (Ulotrix sp.)
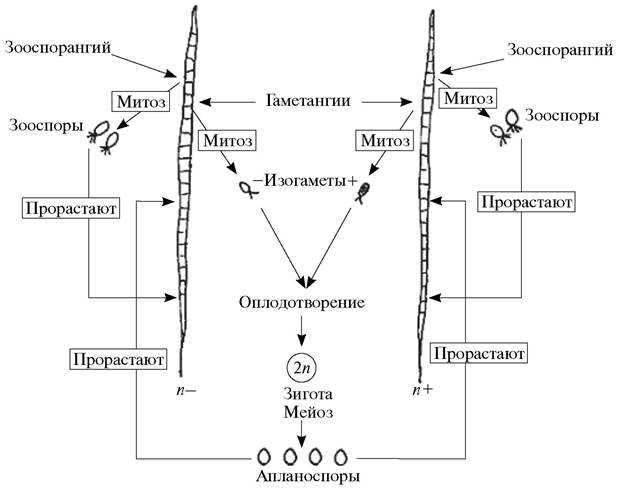
При гетерофазном чередовании поколений смена репродуктивного процесса (или, как неточно говорят, способа размножения — именно так полагал В. Гофмейстер) всегда влечет за собой смену ядерной фазы, т. е. спорообразование происходит на спорофите и сопровождается редукцией числа хромосом,
а образование гамет осуществляется на гаметофите, и в результате процесса оплодотворения число хромосом восстанавливается (см. рис. 138).
Гетерофазное чередование поколений может быть изоморфным и гетероморфным. При изоморфном чередовании поколений гаметофит и спорофит морфологически не различимы, но они отличаются по цитологическим, физиолого-биохимическим и функциональным свойствам (рис. 140). Изоморфное чередование поколений встречается довольно редко и только у водорослей. Оно характерно для зеленых водорослей (ульва — Ulva, энтероморфа — Еnteromorpha, кладофора — Cladophora), а также у представителей ряда порядков бурых и большинства красных водорослей.
Рис. 140. Схема гетерофазного изоморфного чередования поколений у ульвы (Ulva sp.)
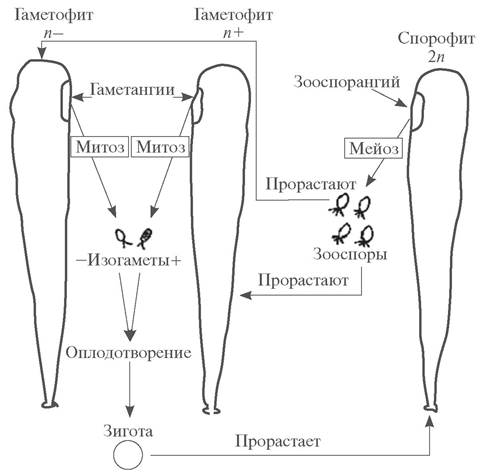
У высокоорганизованных водорослей (бурых, некоторых зеленых и красных) и всех без исключения высших растений чередование поколений гетерофазное гетероморфное, т. е. у большинства растительных организмов гаметофит и спорофит, кроме цитологических, физиолого-биохимических и функциональных различий, имеют четкие морфологические различия (см. рис. 138).
Соотношение полового и бесполого поколений у различных водорослей и высших растений различно. В зависимости от уровня развития гаметофита и спорофита чередование поколений может быть гаплогомофазным, диплогомофазным, гаплодиплофазным и диплогаплофазным.
У видов с гаплогомофазным циклом развития доминирует гаметофит, диплоидна только зигота (примитивные зеленые водоросли). Для таких особей характерен начальный, или зиготный, тип мейоза, т. е. редукционно делится только ядро зиготы, с последующим цитокинезом и образованием апланоспор.
При диплогомофазном чередовании поколений доминирует спорофит, гаплоидны только гаметы (диатомовые — Bacillariophyta, из бурых — представители порядка фукусовые — Fucales, зеленые водоросли, имеющие сифональное строение). В этом случае мейоз предшествует образованию гамет. Такой тип мейоза называется конечным, или гаметным. Он характерен для животных и крайне редок у растений.
При гаплодиплофазном чередовании поколений доминирует гаметофит. У водорослей спорофит существует самостоятельно и может морфологически не отличаться от гаметофита (Ulva), или имеет микроскопическое строение (акросифония из Chlorophyta), или спорофит развивается на гаметофите (филлофора Броди из Rhodophyta). У высших растений только мохообразные характеризуются гаплодиплофазным циклом развития (рис. 141). При этом спорофит в виде коробочки всегда развивается на гаметофите, частично питается за его счет, хотя на ранних этапах развития имеет зеленую окраску и, следовательно, способен к фотосинтезу. Мохообразные являются равноспоровыми растениями, однако гаметофиты их (т. е. сами растения) могут быть как однодомными, так и двудомными. У однодомных растений антеридии и архегони образуются на одном и том же растении (Sphagnum squarrosum, S. compactum, Mnium cuspidatum). У двудомных растений половые органы одного типа (антеридии или архегонии) образуются на разных особях (Marchantía polymorpha, Sphagnum magellaniсum, Polytrichum commune).
Рис. 141. Схема гаплодиплофазного чередования поколений у мохообразных
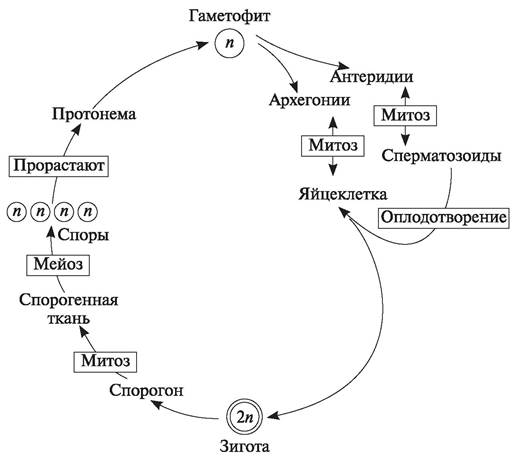
При диплогаплофазном типе развития в жизненном цикле высших растений доминирует спорофит. Гаметофиты могут существовать независимо от спорофитов (плаунообразные, хвощеобразные, папоротникообразные), или они формируются на спорофитах (голосеменные, покрытосеменные).
При гаплодиплофазном и диплогаплофазном жизненных циклах мейоз предшествует образованию спор. Такой тип мейоза называется споровым, или промежуточным. Он характерен для всех без исключения высших растений, а также встречается у многих водорослей.
Растения с гаплодиплофазным и диплогаплофазным циклами развития могут быть как равноспоровыми, так и разноспоровыми.
У разноспоровых (селягинелловые, сальвиниевые) высших растений по сравнению с равноспоровыми, заростки сильно редуцированы (рис. 142). Однако это явление имеет большое биологическое значение.
Рис. 142. Спорофит, спорангии, споры, гаметофиты селягинеллы (Selagmella sp.): А—внешний вид спорофита; Б—микроспорангий с микроспорами; В—микроспора; Г— мужской гаметофит; Д—макроспорангий с макроспорами; Е—макроспора; Ж—женский гаметофит: 1—оболочка микроспоры; 2—оболочка макроспоры
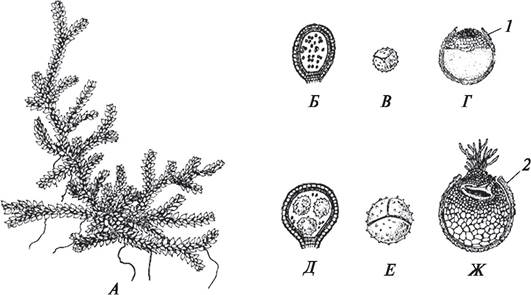
Заростки у селягинелловых, сальвиниевых развиваются почти полностью внутри оболочки споры, следовательно, они надежно защищены от внешней среды (от неблагоприятных условий) и быстро формируются в процессе онтогенеза, что очень важно с эволюционной точки зрения.
У семенных растений (для них характерна гетероспория) при крайней степени редукции мужского гаметофита (пыльцевых зерен, пыльцы) расширяются его функции. Наряду с типичной функцией образования гамет (половой функцией) мужской гаметофит выполняет транспортную функцию. Мужские гаметы доставляются к яйцеклетке (в архегоний у голосеменных растений или в зародышевый мешок у покрытосеменных) пыльцевой трубкой, которая формируется у мужского гаметофита.
В биологии развития разноспоровых растений особого внимания заслуживают три особенности:
1) дифференциация спор — физиологическая и морфологическая, причем функцию расселения вида у высших разноспоровых осуществляет мегаспора;
2) надежная защита гаметофитов от неблагоприятных условий;
3) редукция гаметофитов, и как следствие этого, их быстрое развитие в онтогенезе.
В процессе эволюции у семенных растений мегаспора утрачивает функцию расселения, оно осуществляется семенами. У них более выражены черты приспособления к условиям существования на суше: происходит крайняя редукция гаметофитов, особенно мужских; наряду со сперматозоидами (саговник — Cycas, гинкго — Ginкgo) появляются спермии (хвойные, все покрытосеменные); при прорастании мужского гаметофита образуется пыльцевая трубка и возникает сифоногамия, которая делает процесс оплодотворения не зависящим от наличия воды.
Конечный результат появления разноспоровости — сокращение продолжительности жизненного цикла растительного организма.
6.3.2. Представление об эволюции жизненных циклов растений
У различных таксонов наблюдаются некоторые специфические особенности жизненного цикла, но в целом для растений основные направления эволюции жизненных циклов вырисовываются довольно четко.
1. На ранних этапах эволюции все особи выполняли и вегетативные, и репродуктивные функции. Половой и бесполый процессы равнозначны. Одна и та же особь потенциально могла формировать и споры, и гаметы (см. рис. 139).
2. Постепенно происходит дифференцировка особей по способам репродуктивных процессов на спорофиты и гаметофиты, но морфологически они не различаются (см. рис. 140).
3. Появляются морфологические различия между гаметофитом и спорофитом, но доминирует гаметофит. Спорофит развивается на гаметофите (см. рис. 141).
4. Спорофит отделяется от гаметофита, т. е. происходит преодоление зависимости спорофита от гаметофита (см. рис. 134, 135, 136).
5. В жизненном цикле закрепляется господствующее положение спорофита. Первоначально для него характерна изоспория (равноспоровость). Спора выполняет функцию расселения вида.
6. Появляется гетероспория (разноспоровость), что приводит к половой дифференцировке гаметофитов. Функцию расселения вида выполняет только макроспора (мегаспора) (см. рис. 142).
7. В связи с гетероспорией происходит редукция гаметофитов, что ускоряет процесс их развития. В то же время расширяются функции мужского гаметофита: кроме образования гамет и выполнения половой функции он приобретает транспортную функцию (доставляет мужские гаметы к яйцеклетке).
8. Постепенно устанавливается строгая последовательность смены спорообразования и полового процесса.
9. У высокоорганизованных высших растений (голосеменные и покрытосеменные) гаметофиты достигают крайней степени редукции, развиваются на спорофитах. Макроспора утрачивает функцию расселения вида, она никогда не покидает материнского растения.
10. На основании сочетания спорообразования и полового процесса возникает семя — сложное образование, в котором формируется зачаток нового организма (новый спорофит) и сохраняются остатки материнского спорофита. У покрытосеменных это семенная кожура и у некоторых растений питательная ткань — перисперм (гвоздичные, маревые). У голосеменных в семени есть еще и хорошо развитый и превращенный в трофическую ткань женский гаметофит (первичный эндосперм).
Чередование поколений имеет эволюционное и биологическое значение.
Большое эволюционное значение чередования поколений состоит в том, что смена репродуктивных процессов привела к доминированию в жизненном цикле спорофита, который чрезвычайно сложно устроен и наилучшим образом приспособлен к жизни в разнообразных условиях.
Биологическое значение чередования поколений заключается в том, что постепенно, с усложнением морфофизиологического строения спорофита и редукцией гаметофита совершенствовался половой процесс, и его конечный этап — оплодотворение, которое стало осуществляться без наличия капельножидкой среды, что очень важно в условиях наземного существования.