МОРФОЛОГИЯ РАСТЕНИЙ - Т. А. Сауткина - 2012
ГЛАВА 7. ЦВЕТОК КАК ОСОБЫЙ РЕПРОДУКТИВНЫЙ ОРГАН ПОКРЫТОСЕМЕННЫХ РАСТЕНИЙ
7.3. Гипотезы происхождения цветка
В связи с тем, что цветковые растения играют большую роль в жизни человека, вопрос об их происхождении, а также о происхождении их специализированного репродуктивного органа — цветка — по-прежнему занимает первостепенное место в научных исследованиях ботаников разных профилей. Несмотря на то что морфологи, систематики, эмбриологи, палеоботаники накопили большое количество фактического материала, до настоящего времени не потеряли своего значения слова великого натуралиста Ч. Дарвина «об ужасной тайне, окружающей цветковые растения».
Действительно, вопрос о морфологической природе частей цветка, который пытался выяснить еще И. В. Гёте, не разрешил проблемы происхождения цветка как особого репродуктивного органа.
Стремление отыскать возможных предков покрытосеменных растений и на основании изучения их репродуктивных органов попытаться выявить орган, который мог бы быть предшественником цветка, побудили ботаников обратить внимание на ископаемые голосеменные. Так возникли псевдантовая (от греч. pseudo — ложь, anthos — цветок) и эуантовая, или стробилярная (от греч. eu — истина, anthos — цветок), гипотезы происхождения цветка.
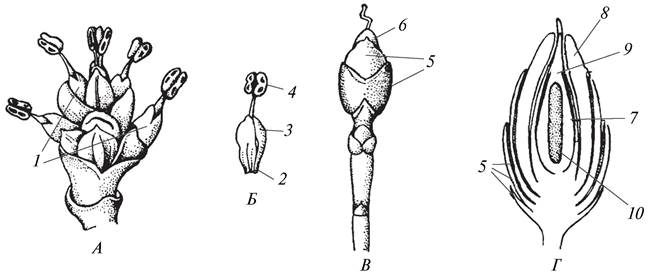
Псевдантовая гипотеза была сформулирована австрийским ботаником Р. Веттштейном в 1901 г. В качестве предковой группы покрытосеменных растений он рассматривал ископаемые голосеменные эфедровые, у которых типичных шишек нет, но существуют своеобразные структуры, которые называют микростробилами и макростробилами (рис. 147). Отдельный микростробил внешне похож на тычинку покрытосеменных, а группа микростробилов, окруженных кроющими листьями, — на их соцветие. Макростробил состоит из одного семязачатка (семяпочки) и окружен покровом (стерильными чешуевидными листьями). Р Веттштейн полагал, что из микростробила в результате редукции кроющих листьев мог возникнуть тычиночный цветок с простым околоцветником, а из макростробила — голый (без околоцветника) женский цветок. Согласно представлению, Р. Веттштейна, обоеполый цветок покрытосеменных гомологичен соцветию (собранию микро- и макростробилов) эфедровых, но убедительно доказать, каким образом из однополых цветков могли образоваться обоеполые, ученому не удалось. Постепенно ботаники от этой гипотезы отказались. Однако в свое время псевдантовая гипотеза сыграла определенную роль и была положена в основу ряда систем покрытосеменных растений. Самой известной из них является система А. Энглера (1844—1930), единственная система, филогенетические связи в которой прослеживаются до рода. До настоящего времени ею пользуются в ряде крупных гербариев, в том числе и в Гербарии Ботанического института имени В. Л. Комарова Российской академии наук (г. Санкт-Петербург).
Рис. 147. Репродуктивные структуры эфедры (Ephedra sp.): А—микростробилы; Б—внешний вид микростробила; В—внешний вид макростробила (семяпочки); Г—продольный разрез семяпочки: 1—собрание микростробилов; 2—отдельный микростробил; 3—кроющий лист микростробила; 4—синангий (пыльник с микроспорангиями); 5—стерильные чешуевидные листья; 6—семяпочка (макроспорангий); 7—нуцеллус семяпочки; 8—интегумент; 9—пыльцевая камера; 10—женский гаметофит
К концу ХК — началу ХХ в. создались предпосылки для осмысления путей происхождения цветка в процессе эволюции. Опираясь на данные палеоботаники и морфологии, американский ботаник и филогенетик Ч. Бэсси (1845—1915) отмечает, что репродуктивный орган покрытосеменных состоит из стебля, на котором развиваются спороносные и стерильные листья, а позднее высказывает предположение, что покрытосеменные произошли от стробилоносных саговниковых.
Независимо от Ч. Бэсси немецкий систематик Г. Галлир в 1902 г. на основании изучения позеленевшего пролиферирующего цветка водосбора (Aquilegia vulgaris) рассматривает цветок как видоизмененный репродуктивный побег. Таким образом, Ч. Бэсси и Г. Галлир фактически явились основоположниками эуантовой, или стробилярной, гипотезы происхождения цветка. Однако окончательно она была сформулирована несколько позже.
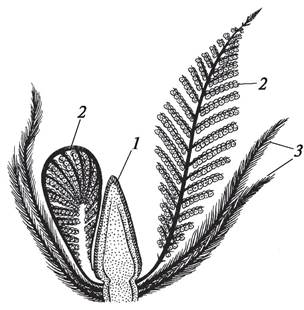
В 1906 г. немецкий физиолог растений Г. Виланд (1877—1957) описал обоеполый стробил ископаемых голосеменных — беннетитовых (рис. 148).
Рис. 148. Реконструкция продольного разреза стробила беннетита Cycadeoidea dacotensis: 1—собрание макроспорофиллов с макроспорангиями; 2—микроспорофиллы с микроспорангиями; 3—кроющие листья стробила
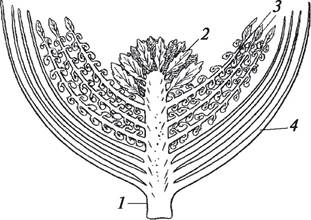
Это открытие позволило Э. Арберу и Дж. Паркину окончательно сформулировать стробилярную гипотезу происхождения цветка. Согласно этой гипотезе, цветок представляет собой обоеполый стробил, приспособленный для размножения. Цветок в представлении Э. Арбера и Дж. Паркина — это новейшая модификация стробила — антостробил (рис. 149). Э. Арбер и Дж. Паркин считали, что беннетито- вые, очевидно, не прямые предки покрытосеменных растений, но, возможно, они являются предками каких-то гипотетических полупокрытосеменных, которые до сих пор не выявлены. В своем труде «О происхождении покрытосеменных» (1907) авторы устанавливают признаки примитивности и специализации цветка и высказывают свою точку зрения на филогенетическую систему покрытосеменных. Она диаметрально противоположна точке зрения Р. Веттштейна. Если Р. Веттштейн считал наиболее примитивными семейства, представители которых имели мелкие раздельнополые ветроопыляемые цветки без околоцветника или с редуцированным околоцветником, то, по Э. Арберу и Дж. Паркину, самые примитивные семейства включают представителей с крупными обоеполыми насекомоопыляемыми цветками, имеющими простой или двойной околоцветник.
Рис. 149. Проантостробил — гипотетический цветок анцестрального вида покрытосеменных: 1—ось цветка; 2—макроспорофиллы; 3—микроспорофиллы; 4—покровные листья
Открытие псилофитовых (риниообразных), признанных наиболее примитивными высшими растениями, дало основание вновь вернуться к проблеме происхождения цветка и выдвинуть теломную теорию. Основоположником ее является В. Циммерман (1930), который считал, что все органы высших растений развились из простых и недифференцированных элементов, названных им теломами. Согласно представлениям, В. Циммермана, стерильные телломы — филлоиды и фертильные теломы, несущие спорангии, могли срастаться и формировать синтеломы. Цветок, по В. Циммерману, представляет собой синтелом, процесс возникновения которого происходил параллельно процессу формирования вегетативного побега. И хотя детально эта теория не разработана, она имела большое значение для решения некоторых вопросов о природе элементов цветка. С 1940—50-х гг. — периода особенно острых дискуссий сторонников теломной теории и приверженцев классической морфологии — цветок перестали определять, как метаморфизированный вегетативный побег.
По мнению члена-корреспондента Российской академии наук, профессора МГУ имени М. В. Ломоносова В. Н. Тихомирова (1932—1998), принципиально невозможно свести все разнообразие цветков к единой модели. Принимая во внимание полиморфизм цветков, он предполагал, что цветки современных покрытосеменных растений сформировались разными путями в процессе эволюции репродуктивных структур от общей анцестральной (предковой) группы.
В связи с появлением новых научных данных возникают и новые гипотезы происхождения цветка, но фактически ни одна из них не является достаточно убедительной. Суммируя все знания в этой области, известный советский ботаник Б. М. Козо-Полянский (1890—1957) вынужден был констатировать: «Не известны ни предок цветковых растений, ни конкретный орган, исторически исходный для цветка».