МОРФОЛОГИЯ РАСТЕНИЙ - Т. А. Сауткина - 2012
ГЛАВА 7. ЦВЕТОК КАК ОСОБЫЙ РЕПРОДУКТИВНЫЙ ОРГАН ПОКРЫТОСЕМЕННЫХ РАСТЕНИЙ
7.4. Общий план строения цветка
Цветки покрытосеменных растений чрезвычайно разнообразны по строению, форме, величине, однако у них имеются четко выраженные общие признаки (рис. 150). В состав цветка входят цветоложе, околоцветник (иногда он отсутствует), тычинки и пестики. При наличии тычинок и пестиков цветки называют обоеполыми, если в цветке имеется один из этих элементов, — однополыми (рис. 151). Очень редко тычинки и пестики вообще отсутствуют (воронковидные цветки василька — Centaurea cyanus).
Рис. 150. Обобщенная схема строения обоеполого цветка: 1—цветоложе; 2—чашелистики чашечки; 3—лепесток венчика; 4—тычинка; 5—пыльник тычинки; 6—завязь пестика; 7—столбик пестика; 8—рыльце пестика; 9—семяпочки в завязи
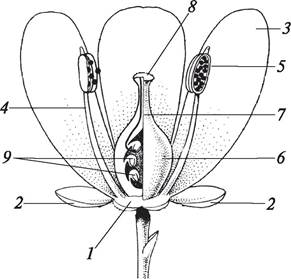
Рис. 151. Однополые цветки: А—дынное дерево (Carica papaya); Б—болотник болотный (Callitriche palustris); В—бегония всегда цветущая (Begonia semperflorens): 1— мужской цветок; 2—женский цветок; 3—тычинки; 4—пестик; 5—чашечка; 6—венчик; 7—верхушечные листья; 8—нижняя завязь цветка
Цветоложе (лат. receptaculum) — осевая часть цветка, на которой располагаются все его элементы. Цветоложе может быть вытянутым — коническим, что подтверждает его осевое происхождение, или укороченным — плоским. Коническое цветоложе встречается у представителей примитивных семейств Магнолиевые (Magnoliaceae), Лютиковые, (Ranunculaceae). У большинства же покрытосеменных растений цветоложе плоское, а у представителей семейства Розоцветные (Rosaceae) оно блюдцевидно расширенное (земляника лесная — Fragaria vescan) или бокальчато вогнутое (слива обыкновенная — Prunus vulgaris) и называется гипантий.
Элементы цветка расположены на цветоложе в определенном порядке.
Все элементы цветка можно подразделить на стерильные (не принимающие непосредственного участия в репродуктивных процессах) и фертильные (в которых происходят репродуктивные процессы). К стерильным элементам относится околоцветник цветка, к фертильным — тычинки и пестики.
Околоцветник цветка, или его покровы (лат. periantium), в зависимости от строения может быть двойным и простым.
Двойной околоцветник состоит из чашечки (лат. calyx) и венчика (лат. corolla).
Чашечка образована чашелистиками, имеющими листовую природу, о чем свидетельствуют анатомические данные. Для чашелистиков характерно наличие трех листовых следов, т. е. такое же число листовых следов, которое типично и для вегетативных листьев. Чашелистики могут быть не сросшимися, тогда образуется свободнолистная чашечка (лютик едкий — Ranunculus acris, звездчатка злаковидная — Stellaria gramínea). Если чашелистики в той или иной степени срастаются, формируется сростнолистная чашечка (куколь обыкновенный — Agrostemma githago, вероника дубравная — Veronica chamaedrys). Чашелистики чаще всего окрашены в зеленый цвет, но могут иметь и иную окраску: желтую (лютик едкий), синюю (живокость крупноцветковая — Delphinium grandiflorum), оранжевую (купальница азиатская — Trollius asiaticus), белую, розовую, фиолетовую (гортензия садовая — Hydrangeahortensis). Чашелистики на стадии бутона выполняют защитную функцию, а после раскрытия бутона зеленые чашелистики принимают участие в процессе фотосинтеза, а ярко окрашенные — в привлечении насекомых-опылителей.
За чашечкой располагается венчик, состоящий из лепестков. Как и чашечка, он может быть свободнолепестным (звездчатка злаковидная — Stellaria graminea, очиток едкий — Sedum acre) и спайнолепестным (колокольчик средний — Campanula media, наперстянка крупноцветковая — Digitalis grandiflora). Морфологическая природа лепестков различными авторами трактуется по- разному. По мнению одних (И. В. Гёте и его последователи), лепестки произошли, как и чашелистики, из верхушечных листьев, другие авторы (О. П. Декандоль и др.) считали их производными тычинок. Э. Арбер, Дж. Паркин, Б. М. Козо-Полянский, А. Имс полагали, что лепестки могут иметь двоякое происхождение. Подтверждением тому являются данные васкулярной анатомии. Так, у некоторых лютиковых, лепестки, как и тычинки, имеют только один листовой след, в то время как у представителей семейств Магнолиевые (Magnoliaceae), Лимонниковые (Schisandraceae) лепестки, как и чашелистики, имеют по три листовых следа. Лепестки выполняют защитную функцию, а также функцию привлечения насекомых-опылителей, чему способствует не только яркая окраска лепестков венчика, но и образование на лепестках (шелковник — Batrachium) или из них же (борец — Aconitum, водосбор — Aquilegia) различных по строению нектарников.
Если околоцветник однородно окрашен, его называют простым. В зависимости от окраски различают чашечковидный, окрашенный в зеленый цвет, (свекла обыкновенная — Betavulgaris, марь белая — Chenopodium album), и венчиковидный, окрашенный в разные цвета (нарцисс поэтический — Narcissus poe'ticus, безвременник осенний — Colchicum autumnale), околоцветник. Однако иногда простой околоцветник, образованный чашелистиками, бывает венчиковидно окрашенным (борец аптечный — Aconitum napellus, сокирка обыкновенная — Consolida regalis).
Окраска элементов околоцветника обусловлена различными пигментами. Главными пигментами цветков являются флавоноиды (антоцианы и флавоны), они растворимы в воде и содержатся в клеточном соке вакуолей. В зависимости от рН клеточного сока флавоноиды окрашиваются по-разному: так, например, антоциан цианидин в кислой среде дает красную окраску; в нейтральной — фиолетовую; в щелочной — синюю. Желтую окраску элементам цветка придают флавоны (лютеин и др.) и каротиноиды. Каротиноиды растворимы в липидах и содержатся в пластидах — хромопластах. Коричневый пигмент — антофеин — образует черные или коричневые пятна на лепестках венчиков некоторых бобовых (бобы обыкновенные) и многих тропических орхидей. Смешение пигментов при разных рН образует всю гамму окрасок цветка. У некоторых растений (медуницы, незабудки) окраска венчика в процессе цветения меняется, что связано с изменением рН клеточного сока. Белые цветки не содержат пигментов, их окраска объясняется наличием крупных заполненных воздухом межклетников в тканях лепестков, а иногда и чашелистиков (клеродендрон Томпсона), благодаря чему хорошо отражается солнечный свет.
Важнейшими элементами цветка являются тычинки и пестики.
7.4.1. Андроцей и его типы. Эволюция и строение тычинки
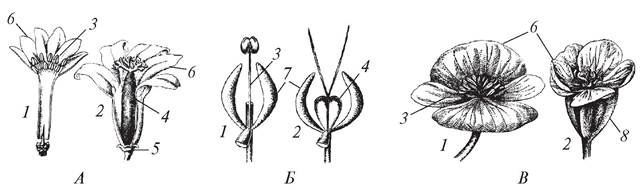
Совокупность тычинок составляет андроцей (androeceum) цветка. В зависимости от строения различают андроцей одно-, дву- и многобратственный. В однобратственном андроцее все тычинки сращены тычиночными нитями (мальва разрезнолистная — Malva excisa, люпин многолетний — Lupinuspoly- phyllus) или пыльниками (сложноцветные) в одну группу. Если тычинки срастаются в две группы, андроцей называется двубратственным (горошек мышиный — Vicia cracca, тыква обыкновенная — Cucurbita pepo). Когда формируется несколько групп сросшихся тычинок (зверобой продырявленный — Hypericum perforatum) или если все тычинки свободные (прострел раскрытый — Pulsatilla patens), андроцей считается многобратственным. Андроцей может быть двусильным, тогда в цветке из четырех тычинок две длинные, а две короткие (сем. Губоцветные — Labiatae), или четырехсильным — в цветке четыре тычинки длинные, а две короткие (сем. Крестоцветные — Cruciferae) (рис. 152).
Рис. 152. Типы андроцея у представителей различных семейств покрытосеменных: 1—многобратственный двусильный (сем. Губоцветные); 2—многобратственный четырехсильный (сем. Крестоцветные); 3—однобратственный (сем. Мальвовые); 4—однобратственный (сем. Сложноцветные); 5—двубратственный (сем. Бобовые)
Тычинка состоит из тычиночной нити, пыльника, образованного двумя половинками (двумя теками), и связника — паренхимной ткани, которая соединяет половинки пыльника. Тычинкапредставляет собой редуцированный микроспорофилл, остатком которого являются тычиночная нить и связник, с микроспорангиями, находящимися в пыльнике.
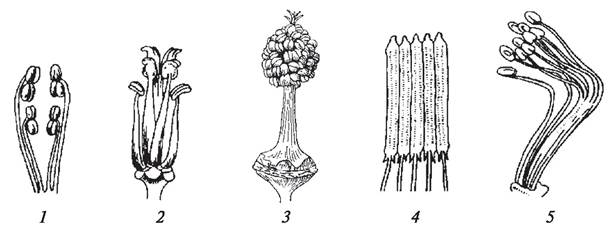
Сравнительное изучение тычинок различных представителей покрытосеменных позволило составить представление об эволюции этого репродуктивного элемента (рис. 153). У современных примитивных покрытосеменных (особенно у рода дегенерия, некоторых магнолий) тычинки устроены очень архаично: они плоские, широкие, не расчлененные на тычиночную нить, пыльник и связник, с линейными микроспорангиями, расположенными ламинально, т. е. на поверхности пластинчатого микроспорофилла. Пыльники располагаются как на адаксиальной (внутренней) стороне и тогда вскрываются интрорзно, так и на абаксиальной (внешней) стороне. В этом случае они вскрываются экстрорзно. В примитивных микроспорофиллах имеются три хорошо развитых проводящих пучка (три жилки). Постепенно в процессе эволюции микроспорофилл редуцировался, уменьшилось число проводящих пучков в мезофилле, исчезла большая его часть и в конечном счете сформировалась специализированная тычинка, дифференцированная на тычиночную нить, пыльник и связник. В тычиночной нити имеется один проводящий пучок, который доходит до связника и поставляет к его живым паренхимным клеткам питательные вещества. По мере дифференцировки тычинки меняется положение пыльника. Постепенно половинки пыльника перемещаются в боковое положение, а затем вследствие дальнейшей редукции связника сливаются в пыльник, состоящий из двух половинок (двух тек), который занимает верхушечное положение. Таким образом, у большинства покрытосеменных пыльник содержит четыре микроспорангия и представляет собой синангий (совокупность спорангиев).
Рис. 153. Стадии эволюции микроспорофилла от примитивного пластинчатого типа Degeneria до высокоспециализированной тычинки: А—Г — уменьшение размеров микроспорофилла, редукция жилок и изменение положения микроспорангиев; Д—строение тычинки: 1—микроспорофилл; 2—микроспорангии; 3—жилки микроспорофилла; 4—формирующийся связник; 5—формирующийся надсвязник; 6—микроспорангии (пыльники); 7—тычиночные нити
У некоторых растений в результате приспособления к опылению часть тычинок утрачивает пыльники и превращается в нектарники — стаминодии (белозор болотный — Parnassia palustris, купальница европейская — Trollius europaeus).
Число тычинок в обоеполых цветках варьирует в широких пределах — от бесконечно большого (сем. Лютиковые — Ranunculaceae, Магнолиевые — Magnoliaceae) до двух (вероника дубравная — Veronica chamaedrys) и даже одной (любка двулистная — Platanthera bifolia). В однополых тычиночных цветках количество тычинок также бывает различным (см. рис. 151, Б, В; 159, В).
Тычинки в цветке располагаются или по спирали (сем. Магнолиевые, большинство представителей сем. Лютиковые), или по кругу (сем. Розоцветные — Rosaceae, Гвоздичные — Caryophyllaceae).
7.4.2. Гинецей и его типы. Эволюция плодолистика, образование и строение пестика
Центральную часть цветка занимает один или несколько пестиков. Совокупность пестиков или сросшихся плодолистиков составляет гинецей(gynoeceum) цветка.
Пестик представляет собой замкнутый видоизмененный и высокоспециализированный макроспорофилл (мегаспорофилл, плодолистик, карпелла), внутри которого заключены семяпочки. Он образуется в результате срастания краев плодолистика и дифференцирован на полую завязь, стилодии (stylodium — суженная часть плодолистика) или столбик (stylos — образование, возникающее в результате срастания нескольких стилодиев) и рыльце.
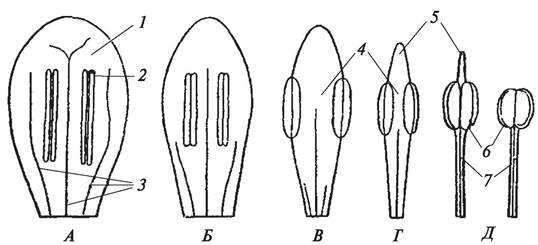
Листовая природа плодолистиков была научно обоснована О. П. Декандолем еще в 1827 г., хотя он ошибочно выводил их не из спороносных, а из вегетативных листьев. И действительно, чем примитивнее цветок, тем более листовидный характер носят отдельные плодолистики, а проводящая система их сходна с проводящей системой листа. Согласно представлениям, А. Л. Тахтаджяна, от открытого развернутого макроспорофилла плодолистик покрытосеменных отличается тем, что он как бы сложен по центральной жилке, т. е. сдвоенный (кондупликатный) (рис. 154).
Рис. 154. Стадии эволюции плодолистика (макроспорофилла): А—примитивный кондупликатный плодолистик типа Degeneria с рыльцевым гребнем; Б, В—стадии срастания краев плодолистика и уменьшение рыльцевого гребня; Г, Д—стадии формирования пестика; Е—сформированный пестик: 1—кондупликатный плодолистик;
2—рыльцевый гребень; 3—завязь пестика; 4—стилодий; 5—головчатое рыльце
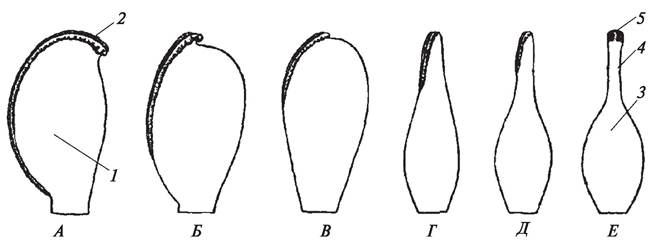
В завязи пестика находятся семяпочки (семязачатки). В связи с кондупликатностью плодолистика семяпочки, нуцеллус которых является макроспорангием, оказываются на адаксиальной (внутренней) стороне, что повышает уровень их защищенности от различных неблагоприятных воздействий (сухости воздуха, высоких и низких температур, резких перепадов температур, повреждения насекомыми и т. д.).
Постепенно сближенные края кондупликатного плодолистика начинали срастаться, что привело в конечном счете к образованию типичного замкнутого пестика. Среди современных покрытосеменных растений самые архаичные пестики обнаружены у дегенерии (Degeneria), у которой каждый пестик состоит из плодолистика, сросшегося только в основании. Верхние же края плодолистика не только не сращены, но фактически даже не соприкасаются друг с другом (А. Л. Тахтаджян, 1964). Неполное срастание краев плодолистиков наблюдается также и у некоторых более продвинутых представителей покрытосеменных растений (пион — Paeonia, платан — Platanus). У резедовых (резеда желтая — Reseda lutea)завязь на верхушке остается незамкнутой.
Так как в кондупликатном плодолистике семяпочки оказались закрытыми и недоступными для пыльцы, возникла необходимость в образовании специальной поверхности, которая могла бы воспринимать пыльцу. Так постепенно у покрытосеменных на краях кондупликатных плодолистиков начала формироваться особая железистая ткань и возникла рыльцевая поверхность, или рыльцевый гребень. Наличие рыльцевого гребня — одна из характернейших особенностей макроспорофиллов примитивных покрытосеменных. Сначала рыльцевый гребень простирался по обоим краям сложенного широкого плодолистика, но по мере срастания краев он укорачивался, менялась его форма, и в конечном счете он превратился в рыльце пестика, которое заняло верхушечное положение. В ходе эволюции дифференцировался и сам плодолистик, постепенно преобразовавшийся в типичный пестик. Верхняя часть его вытянулась и превратилась в стилодий — стерильное образование. У примитивных покрытосеменных стилодий еще кондупликатный, с двумя рыльцевыми гребнями, но постепенно края его срастаются, а на верхушке образуется типичное рыльце.
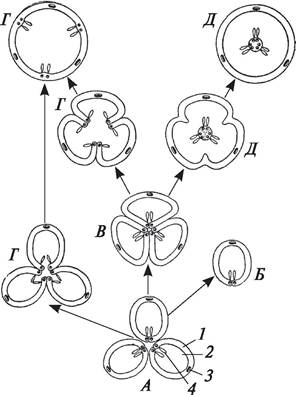
Пестик может быть образован одним или несколькими сросшимися плодолистиками. Гинецей, в котором каждый пестик образован одним сросшимся плодолистиком, называется апокарпным. Апокарпный гинецей может состоять из одного (бобы обыкновенные — Faba vulgaris, вишня обыкновенная — Cerasus vulgaris), нескольких или большого числа пестиков (сем. Магнолиевые, большинство лютиковых). Таким образом, апокарпный гинецей может быть многочленным и одночленным. Многочленный апокарпный гинецей характерен для представителей примитивных семейств, одночленный встречается у эволюционно более продвинутых.
В процессе эволюции происходило усложнение строения гинецея. Благодаря срастанию нескольких плодолистиков сформировался ценокарпный гинецей. Число сросшихся плодолистиков в ценокарпном гинецее варьирует от двух (сем. Крестоцветные — Cruciferae, Злаки — Gramineae) до неопределенно большого (мальва — Malva).
В зависимости от характера срастания плодолистиков ценокарпный гинецей подразделяется на синкарпный, паракарпный и лизикарпный.
В синкарпном гинецее плодолистики полностью срастаются боковыми (латеральными) сторонами, в результате чего образуется многогнездная завязь, число гнезд в которой соответствует числу сросшихся плодолистиков (два у белены черной — Hyoscyamus niger, пять у льна посевного — Linum usitatissimum).
В паракарпном гинецее плодолистики срастаются таким образом, что завязь остается одногнездной (сем. Маковые — Papaveraceae, Крестоцветные — Cruciferae, Тыквенные — Cucur- bitaceae). В лизикарпном гинецее завязь также одногнездная, но он возникает не так, как паракарпный. При образовании лизикарпного гинецея перегородк (стенки сросшихся плодолистиков) разрушаются, но в центре завязи остается колонка — центральная часть сросшихся плодолистиков (сем. Гвоздичные — Caryophyllaceae).
Эволюционное взаимоотношение различных типов гинецея можно представить следующим образом (рис. 155).
Рис. 155. Схема эволюции основных типов гинецея (по Тахтаджяну, 1964, с дополнениями): А—апокарпный многочленный гинецей: 1—стенка завязи; 2—полость завязи; 3—центральный сосудисто-волокнистый пучок плодолистика; 4—семяпочки; Б—апокарпный одночленный гинецей; В—синкарпный гинецей; Г—разновидности паракарпного гинецея; Д—разновидности лизикарпного гинецея
При развитии разных типов ценокарпного гинецея стилодии или остаются совершенно свободными (тмин обыкновенный — Carum carvi, звездчатка средняя — Stellaria media) или частично срастаются (яблоня домашняя — Malus domestica). У многих видов в результате полного срастания стилодиев образуются столбики (редька дикая — Raphanus raphanistrum,вероника лекарственная — Veronica officinalis).
7.4.3. Понятие о семяпочке. Строение и классификация семяпочек
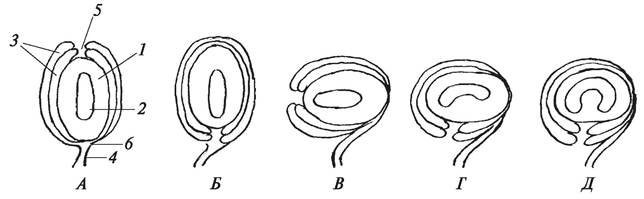
Семяпочка — особая структура голосеменных и покрытосеменных растений, состоящая из макроспорангия (мегаспорангия), защищенного стерильными покровами — интегументами (рис. 156).
Рис. 156. Схема строения и типы семяпочек: А—атропная (или ортотропная) — прямая: 1 — нуцеллус (макроспорангий); 2—зародышевый мешок; 3—интегументы; 4— фуникулус; 5—микропиле; 6—халаза; Б—анатропная — обратная; В—гемитропная — полуповернутая; Г—кампилотропная — односторонне изогнутая; Д—амфитропная — двусторонне изогнутая
Термин «семяпочка» (нем. Samenknospe), введенный И. В. Гёте в 1790 г. в его знаменитой работе «Опыт объяснения метаморфоза у растений», долгое время был традиционным. Однако с середины XIX в. параллельно с этим термином морфологи и эмбриологи растений стали использовать понятие «семязачаток», подчеркивая тем самым, что в конечном счете из семяпочки (семязачатка) образуется семя.
Макроспорангием является центральная часть семяпочки — нуцеллус. В отличие от макроспорангиев высших споровых растений нуцеллус представляет собой массивную выполненную многоклеточную структуру, надежно защищенную интегументами и стенками завязи пестика. Строение нуцеллуса у большинства исследованных видов изучалось преимущественно на светооптическом уровне, что не позволяло выявить ряд существенных моментов.
Электронно-микроскопическое изучение показало, что растущий нуцеллус состоит из меристематических клеток, которые связаны между собой плазмодесмами. По степени развития нуцеллуса Ф. Ван-Тигем в 1898 г. разделил семяпочки на два типа: крассинуцеллятные и тенуинуцеллятные. Для крассинуцеллятных семяпочек характерен массивный многослойный нуцеллус. Как показали электронно-микроскопические исследования, массивный нуцеллус гистологически дифференцирован: в нем выделяется эпидермис, снаружи покрытый слоем кутикулы, и паренхимная ткань. У большинства цветковых растений эпидермис однослойный, так как клетки меристемы, образующей эпидермис, делятся только антиклинально. В то же время у представителей некоторых семейств (сем. Спаржевые — Asparagaceae) меристематические клетки могут делиться периклинально, и тогда в микропилярной части нуцеллуса образуется многослойный нуцеллярный колпачок.
У тенуинуцеллятных семяпочек нуцеллус слабо развит. В клетках эпидермиса данного типа семяпочек плазмодесмы обнаружены только на радиальных стенках.
Основание семяпочки, прилегающее к стенке завязи, называется халазой. У большинства покрытосеменных растений в халазе оканчивается сосудистоволокнистый пучок, идущий из завязи в семяпочку, что позволяет предположить, что халаза выполняет трофическую функцию, передавая продукты метаболизма из завязи в семяпочку, а также защитную функцию.
От халазы отходит семяножка — фуникулус, при помощи которой семяпочка прикрепляется к стенке завязи, и интегументы. В фуникулусе хорошо развита проводящая система, по которой к тканям семяпочки из завязи пестика поступают растворы органических и минеральных веществ.
Интегументы защищают нуцеллус семяпочки и развивающиеся в нем структуры от неблагоприятных условий. Интегументы у различных растений различаются по количеству и степени развития. По числу интегументов семяпочки покрытосеменных растений подразделяются на битегмальные (двупокровные), унитегмальные (однопокровные) и атегмальные (беспокровные). Битегмальные семяпочки характерны для однодольных и многих двудольных, унитегмальные — для спайнолепестных двудольных, атегмальные — обнаружены у некоторых паразитных растений.
Интегументы обрастают нуцеллус, но на противоположном от халазы полюсе не срастаются, в результате чего образуется канал — пыльцевход, или микропиле.
Таким образом, семяпочка является поляризованной структурой. У нее различают два полюса: микропилярный и халазальный.
Строение семяпочек у разных растений различно, что нашло отражение в их морфологической классификации. В зависимости от расположения микропиле по отношению к халазе различают семяпочки атропные, или ортотропные (прямые), анатропные (обращенные), гемитропные (полуобращенные), кам- пилотропные (изогнутые) и амфитропные (обратнообращенные) (см. рис. 156).
В атропных семяпочках микропиле и халаза располагаются на одной вертикальной прямой, в анатропных — микропиле и халаза находятся на одной горизонтальной прямой (на одной стороне), в гемитропных семяпочках микропиле повернуто на 90° по отношению к халазе. В атропных, анатропных и гемитропных семяпочках нуцеллус не изогнут. В кампилотропных и амфитропных — нуцеллус в разной степени изогнут. В настоящее время эмбриологи используют более детальную классификацию семяпочек.
Форма семяпочки, наличие или отсутствие покровов, характер нуцеллуса имеют важное систематическое и филогенетическое значение.
7.4.4. Типы плацентации
Плацентация — это характер расположения плацент — мест прикрепления семяпочек к стенкам завязи пестика. В зависимости от строения гинецея выделяют типы плацентации: ламинальную (поверхностную) и сутуральную (шовную).
При ламинальной плацентации семяпочки прикреплены по внутренней поверхности завязи. Самым примитивным типом ламинальной плацентации является диффузная (ламинально-диффузная), при которой семяпочки рассеяны по всей адаксиальной поверхности завязи (сусак зонтичный — Butomus umbellatus) (рис. 157, Б, 6).
При сутуральной плацентации семяпочки прикреплены или к спинному шву (к средней жилке сросшегося плодолистика), или к брюшному шву (сросшимся краям плодолистика). Сутуральную плацентацию можно подразделить на медианную (ламинально-медианную), латеральную (ламинально-латеральную), центрально-угловую, париетальную (постенную) и центрально-осевую (колончатую) (рис. 157).
Рис. 157. Схемы разных типов плацентации семяпочек: А—в различных типах ценокарпного гинецея; Б—в апокарпном гинецее: 1—центрально-угловая (гинецей синкарпный); 2—париетальная (гинецей паракарпный); 3—центрально-осевая (гинецей лизикарпный); 4—париетально-спинная (медианная); 5—париетально-краевая (брюшная); 6—диффузно-ламинальная; 7—спинной шов; 8—брюшной шов
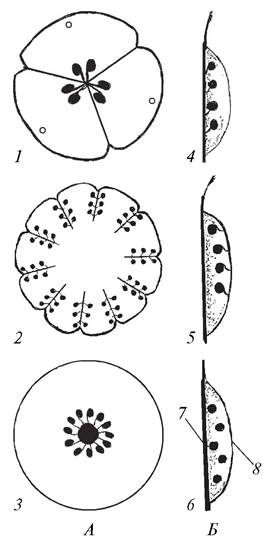
В апокарпном гинецее плацентация может быть медианной и латеральной. При медианной плацентации семяпочки располагаются вдоль средней жилки плодолистика (род лотос — Nelumbo), при латеральной — вдоль брюшного шва (горох посевной — Pisum sativum).
Центрально-угловая плацентация характерна для синкарпного гинецея. При таком типе плацентации семяпочки прикреплены в углах сросшихся плодолистиков (лилия
тигровая — Lilium tigrinum, груша обыкновенная — Pyrus communis). В паракарпном гинецее плацентация париетальная, но семяпочки могут прикрепляться по-разному. Если они располагаются по краям сросшихся плодолистиков, париетальная плацентация называется латеральной (постенно краевой), если вдоль центральной жилки — медианной (постенно - спинной) (фиалка трехцветная — Viola tricolor, огурец посевной — Cucumis sativus).
В лизикарпном гинецее семяпочки прикрепляются к колонке, в этом случае плацентация является центрально-осевой (гвоздика — Dianthus, мыльнянка — Saponaria).
В процессе развития гинецея плацента может сильно разрастаться и образовывать ложные перегородки в завязи (сем. Крестоцветные).
Число семяпочек как в апокарпном, так и в ценокарпном гинецее варьирует от многих до одной. Уменьшение числа семяпочек в завязи пестика, как и уменьшение количества пестиков, а также усложнение строения пестика, — результат эволюционной специализации представителей различных семейств.
7.4.5. Положение завязи в цветке
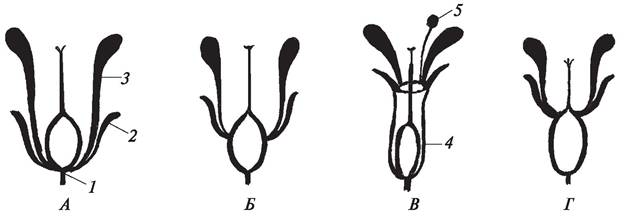
Как результат процесса эволюции можно рассматривать и различное положение завязи в цветке, которое определяют по отношению к околоцвет нику (рис. 158). Если околоцветник прикреплен под завязью, она называется верхней. Такая завязь только основанием прикреплена к цветоложу (роды земляника — Fragaria, лютик — Ranunculus, рожь — Secale). Если околоцветник находится над завязью, то она будет нижней (сем. Колокольчиковые — Campanulaceae, Зонтичные — Umbelliferaе, Касатиковые — Iridaceae). Завязь может быть полунижней, тогда околоцветник прикреплен в центральной ее части (жимолость татарская — Lonicera tatarica). У вишни обыкновенной (Cerasus vulgaris), черешни(Cerasus avium) свободная завязь глубоко «вдвинута» в цветоложе - гипантий. Такую завязь рассматривают как среднюю. Иногда понятия «полунижняя» и «средняя» завязь понимают, как синонимы. Положение завязи в цветке — важный систематический признак. Наиболее примитивным типом считается верхняя завязь.
Рис. 158. Положение завязи в цветке: А—верхняя завязь (цветок подпестичный); Б—полунижняя завязь (цветок полуподпестичный); В—средняя завязь (цветок околопестичный); Г—нижняя завязь (цветок надпестичный): 1—цветоложе; 2—чашелистик чашечки; 3—лепесток венчика; 4—гипантий — цветоложе розоцветных; 5— тычинка
7.4.6. Классификация цветков
Разнообразие строения позволяет классифицировать цветки с использованием следующих признаков: наличие околоцветника и его строение; тип симметрии; особенности строения элементов околоцветника; признак пола цветка; положение завязи в цветке; расположение элементов цветка на цветоложе.
По наличию околоцветника различают цветки ахламидные (рис. 159), апохламидные, гаплохламидные, диплохламидные.
Рис. 159. Цветки без околоцветника (ахламидные): А, Б—обоеполые цветки, соответственно белокрыльника болотного (Calla palustris) и ясеня обыкновенного (Fraxinus excelsior); В, Г—однополые цветки ивы козьей (Salix caprea): 1—пестики; 2—тычинки; 3—кроющие листья; 4—нектарники
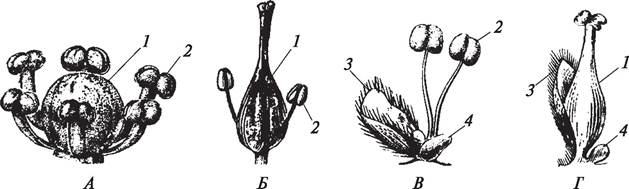
Ахламидные, или голые, цветки не имеют околоцветника и являются первично беспокровными. Растения с такими цветками встречаются как среди на секомоопыляемых (род ива — Salix), так и среди ветроопыляемых (роды осока — Carex, ясень — Fraxinus).
У апохламидных цветков (женские цветки березы) околоцветник также отсутствует, но так как у мужских цветков он имеется, то отсутствие его у женских цветков рассматривается как результат редукции, а, следовательно, как явление вторичное.
У гаплохламидных цветков элементы околоцветника расположены в один круг. Такой околоцветник является простым и может быть, как чашечковидным (зеленым) (крапива двудомная — Urtica dioica), так и венчиковидным, окрашенным в различные цвета (калужница болотная — Caltha palustris, прострел раскрытый — Pulsatilla patens).
В диплохламидном цветке элементы околоцветника расположены в два круга. В зависимости от окраски элементов наружного и внутреннего кругов околоцветник подразделяют на гомохламидный и гетерохламидный.
В гомохламидном околоцветнике листочки однородно окрашены, т. е. он является простым (тюльпан лесной — Tulipa sylvestris, лук репчатый — Allium cepa, ожика волосистая — Luzula pilosa). В зависимости от окраски гомохламидный околоцветник может быть чашечковидным (клен остролистный — Acer platanoides) или венчиковидным (тюльпан лесной — Tulipa sylvestris). В гетерохламидном околоцветнике хорошо различимы чашечка и венчик, такой околоцветник является двойным (незабудка болотная — Myosotis palustris, черемуха обыкновенная — Padus racemosa).
По типу симметрии различают цветки актиноморфные (правильные), зигоморфные (неправильные) и асимметричные.
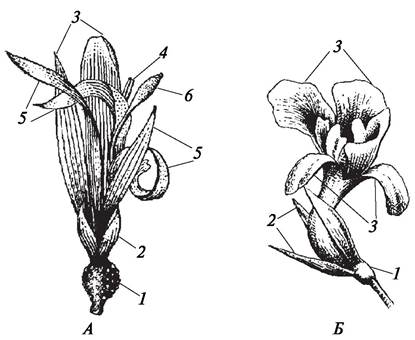
Под симметрией понимают соразмерность, т. е. возможность разделить цветок продольными осями (плоскостями симметрии) на зеркально подобные половинки. Цветок называют актиноморфным (рис. 160), если через него можно провести несколько плоскостей симметрии (прострел раскрытый — Pulsatilla patens, лапчатка серебристая — Potentilla argentea).Через зигоморфный цветок можно провести только одну плоскость симметрии, его можно разде лить лишь на две равные части (львиный зев большой — Antirrhinum majus, борец аптечный — Aconitum napellus) (рис. 161). Асимметричный цветок не имеет ни одной плоскости симметрии (канна индийская — Сanna indica) (рис. 162).
Рис. 160. Актиноморфные цветки: А—схема строения (1—плоскости симметрии); Б—редька дикая (Raphanus raphanistrum); В—лютик едкий (Ranunculus acris); Г—лилия тигровая (Lilium tigrinum)
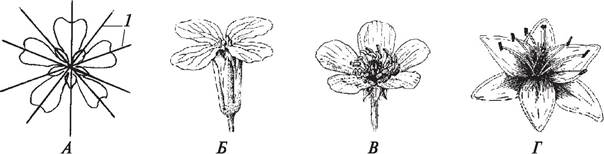
Рис. 161. Зигоморфные цветки: А—схема строения (1—плоскость симметрии); Б—сокирка обыкновенная (Consolida regalis); В—львиный зев большой (Antirrhinum majus);
Г—борец аптечный (Aconitum napellus)
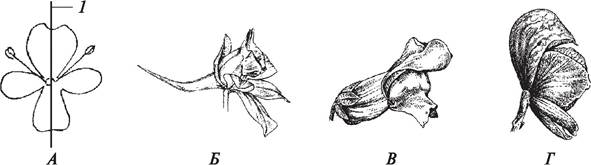
Рис. 162. Асимметричные цветки: А—канна индийская (Canna indica); Б—маранта тростниковидная (Maranta arundinacea): 1—нижняя завязь; 2—чашелистики; 3— лепестки; 4—тычинка; 5—стаминодии; 6—рыльце пестика
Строение околоцветника определяется также тем, как его элементы связаны между собой. Чашечка может быть, как свободнолистной, так и спайнолистной, венчик свободнолепестным и спайнолепестным. Срастание наблюдается как в простом (ландыш майский — nvallaria majalis), так и в двойном околоцветнике. В двойном околоцветнике чашечка может быть сростнолистной, а венчик свободнолепестным (куколь обыкновенный — Agrostemma githago, смолевка обыкновенная — Silene vulgaris) или элементы обоих кругов оказываются сросшимися (львиный зев большой — Antirrhinum majus, картофель — Solanum tuberosum).
Признак пола в цветке определяют репродуктивные элементы — тычинки и пестики. В зависимости от наличия репродуктивных элементов различают цветки обоеполые и однополые (раздельнополые). Обоеполые содержат и тычинки, и пестики (лютик едкий — Ranunculus acris, горох посевной — Pisum sativum). В однополых цветках имеется какой-нибудь один элемент — или тычинки, или пестики. В связи с тем, что тычинка предопределяет мужской пол (так как репродуктивные процессы завершаются образованием мужских гамет — спермиев), цветки, содержащие только тычинки, называются мужскими, или тычиночными. Пестик предопределяет женский пол (так как репродуктивные процессы завершаются образованием женской гаметы — яйцеклетки), поэтому цветки, в которых имеются только пестики, называются женскими, или пестичными (см. рис. 151, 159). Тип цветка по признаку пола у большинства растений является константным. Однако у представителей некоторых семейств наблюдается вариация в строении цветков и расположении их на растении. Так, у зонтичных на одном растении (на одной особи) могут встречаться и мужские, и обоеполые цветки. Такое явление называется андромоноэция. У многих сложноцветных наблюдается гиномоноэция — формирование в одной корзинке женских и обоеполых цветков. У некоторых видов (чемерица Лобеля — Veratrum lobelianum, змеевик большой — Bistorta major) мужские и обоеполые цветки развиваются на разных особях, т. е. проявляется андродиэция. Для представителей семейств Губоцветные, Гвоздичные характерны обоеполые цветки, но часто у них встречается гинодиэция, при которой на разных особях развиваются женские и обоеполые цветки. При гинодиэции половой диморфизм нередко отражается на размерах цветка: обоеполые цветки более крупные, чем однополые.
В связи с наличием у ряда видов однополых цветков, которые могут развиваться на одной и той же или на разных особях, возникло понятие «однодомные» и «двудомные» растения. У однодомных растений мужские и женские цветки располагаются на одной и той же особи (огурец посевной — Cucumis sativus, клещевина обыкновенная — Ricinus communis,кукуруза обыкновенная — Zea mays). Двудомными являются растения, у которых мужские и женские цветки развиваются на разных особях (щавель кислый — Rumex acetosa, ива козья — Salix caprea, конопля посевная — Cannabis sativus). Очень редко встречаются полигамные растения, имеющие мужские, женские и обоеполые цветки (лимонник китайский — Schizandra sinensis). У него на одной особи развиваются как раздельнополые, так и обоеполые цветки.
Важным признаком, который учитывается при классификации цветков, является положение завязи в цветке. В зависимости от положения завязи различают цветки подпестичные — завязь верхняя (сем. Бобовые, Пасленовые), надпестичные — завязь нижняя (сем. Зонтичные, Сложноцветные) и околопестичные — завязь полунижняя или средняя (сем. Жимолостные) (см. рис. 158).
Все элементы цветка по-разному расположены на цветоложе, в связи с чем выделяют три типа цветков: ациклические (спиральные), циклические (круговые) и гемициклические (полукруговые). В ациклических цветках все его элементы расположены по спирали (магнолия крупноцветковая — Magnolia grandiflora, купальница европейская — Trollius europaeus).В циклических цветках все элементы располагаются кругами (гвоздика травянка — Dianthus deltoides, наперстянка крупноцветковая — Digitalisgrandiflora). В гемициклических цветках элементы околоцветника располагаются кругами, а тычинки и пести ки — по спирали (мышехвостник маленький — Myosurus minimus, лютик ползучий — Ranunculus repens).
Совокупность признаков цветка позволяет судить о его примитивности или продвинутости. По каждому признаку можно предположительно указать направление эволюции. Так, простой околоцветник считается более примитивным, чем двойной. Свободнолистность и свободнолепестность возникли раньше, чем спайнолистность и спайнолепестность. Большое количество ты чинок и пестиков — более примитивный признак, чем их ограниченное число, и т. д. Однако очевидно, что формирование признаков цветка у разных семейств покрытосеменных растений происходило параллельно и независимо друг от друга. Поэтому часто у представителей высокоорганизованных семейств в цветке сочетаются особенности высокой организации с примитивными признаками. Это явление получило название гетеробатмии. Так, у розоцветных, прогрессивными признаками строения цветка являются циклическое расположение элементов цветка, наличие нижней завязи и синкарпного гинецея у некоторых представителей. В то же время многие розоцветные имеют верхнюю завязь, апокарпный многочленный гинецей, плод — сборную листовку. Очень часто упрощение строения цветка является не признаком примитивности, а результатом приспособления к условиям существования.
7.4.7. Формулы и диаграммы цветков
Строение цветка — наиболее важный признак покрытосеменных растений. Дать характеристику цветка можно не только словесно, но и при помощи формул и диаграмм.
Формула — это краткая характеристика цветка при помощи символов и числовых индексов. В качестве символов используются начальные буквы латинских названий всех элементов цветка:
P — Perigonium — простой околоцветник;
K — лат. Calyx, нем. Kelch — чашечка;
С — Corolla — венчик;
A — Androeceum — андроцей;
G — Gynoeceum — гинецей.
Тип симметрии цветка обозначают значками:
«*» — актиноморфный (правильный, полисимметричный) цветок;
«↑» — зигоморфный (неправильный, моносимметричный) цветок.
Цифры возле символов показывают число каждого элемента цветка (чашелистиков, лепестков, тычинок, пестиков или сросшихся плодолистиков).
Значком «∞» указывается неопределенно большое число элементов (больше 12). Значок «~» ставится, если число элементов в разных цветках одного и того же вида варьирует (не установившееся число элементов); «+» показывает, что однородные элементы цветка (чашелистики, лепестки, тычинки) располагаются в разных кругах. В скобки ( ) заключаются сросшиеся элементы. Запятыми при необходимости отделяют элементы одного круга, если они имеют какие-то специфические различия.
Черточкой, которая ставится под цифрой, над цифрой или около цифры, характеризующей гинецей, показывают положение завязи в цветке: 2_ — завязь верхняя, (5‾) — нижняя, 1¬— средняя или полунижняя.
Можно условными значками указать пол цветка: «♂» — мужской; «♀» — женский.
Все элементы цветка вводятся в формулу в определенном порядке, от периферии к центру, так, как они расположены на цветоложе.
Примеры формул цветков:
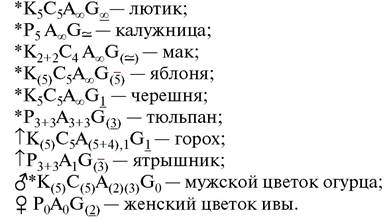
Формулы дают достаточно полную характеристику цветка, однако они не отражают расположение его элементов, тип простого околоцветника, не конкретизируют тип ценокарпного гинецея. Внести эти дополнения позволяет диаграмма цветка.
Диаграмма — это графическое изображение цветка, она строится как проекция поперечного среза бутона на плоскость, поэтому форма диаграммы актиноморфного и зигоморфного цветков будет разной (рис. 163). Для каждого элемента цветка приняты определенные условные обозначения. Чашелистики обозначаются фигурными скобками, лепестки — простыми. Тычинки и пестики, если их немного, можно изобразить в виде формы их поперечного сечения. Если тычинок и пестиков много, их рисуют в виде небольших окружностей. При построении диаграммы необходимо строго отражать взаимное расположение элементов цветка. Лепестки обычно чередуются с чашелистиками.
Рис. 163. Диаграмма цветков: А, Б—актиноморфные цветки; В, Г—зигоморфные цветки: А—птицемлечник зонтичный (Ornithogalum umbellatum); Б—первоцвет обыкновенный (Primulavulgaris); В—сокирка восточная (Consolida orientalis); Г—бобы обыкновенные (Vicia faba)
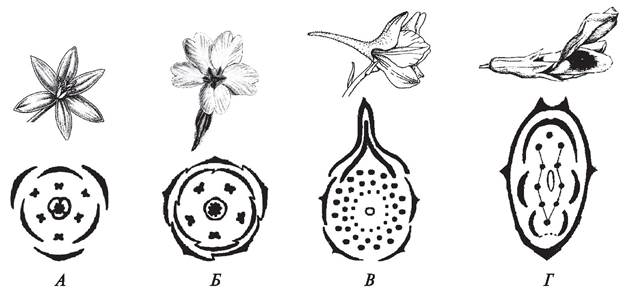
Тычинки в циклических цветках располагаются строго закономерно. У большинства покрытосеменных тычинки наружного круга чередуются с лепестками венчика, такое расположение называется диплостемонией. Реже встречается явление обдиплостемонии, когда тычинки наружного круга располагаются против лепестков венчика (сем. Гвоздичные — Caryophyllaceae, Первоцветные — Primulaceae, свекла обыкновенная — Beta vulgaris).
При наличии формулы и диаграммы можно дать исчерпывающую характеристику цветка.