МОРФОЛОГИЯ РАСТЕНИЙ - Т. А. Сауткина - 2012
ГЛАВА 8. ОПЫЛЕНИЕ И ОПЛОДОТВОРЕНИЕ ЦВЕТКОВЫХ РАСТЕНИЙ. ОБРАЗОВАНИЕ ЗАРОДЫША И ЭНДОСПЕРМА
8.1. Опыление покрытосеменных растений
Несмотря на то что образование женского гаметофита начинается значительно позже мужского, созревание гаметофитов происходит почти одновременно. К моменту созревания мужского гаметофита женский гаметофит готов к процессу оплодотворения.
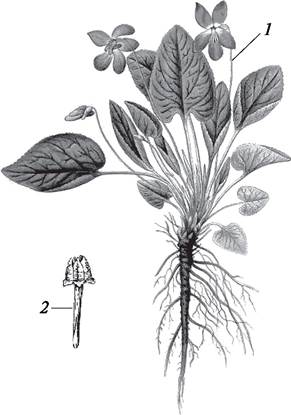
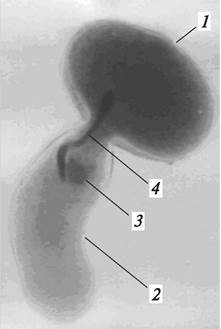
У большинства покрытосеменных растений процессу оплодотворения предшествует опыление — перенос пыльцы из тычинок на рыльце пестика. В ходе эволюции сформировалось два типа опыления: автогамия (самоопыление) и аллогамия, или ксеногамия (перекрестное опыление). При самоопылении пыльца переносится с тычинок на пестик в пределах одного и того же цветка. При перекрестном опылении пыльца одного цветка переносится на рыльце пестика другого цветка. Самоопыление может наблюдаться как в открытых (хазмогамных) цветках (горох — Pisum), так и в закрытых (клейстогамных) (фиалка — Viola, кислица — Oxalis) (рис. 188). У некоторых фиалок и кислицы обыкновенной рано весной развиваются хазмогамные цветки, однако так как весной опылителей мало, эти цветки, как правило, не опыляются и остаются бесплодными. Позже у таких растений наряду с хазмогамными цветками у основания побегов развиваются невзрачные клейстогамные цветки, в которых происходит опыление и оплодотворение, и, как следствие этого, образуются плоды и семена.
Рис. 188. Хазмогамные (1) и клейстогамные (2) цветки у фиалки жестковолосистой (Viola hirta)
Особым типом опыления является гейтоногамия — перенос пыльцы с одного цветка на другой, но находящийся на одном и том же растении. Одни ученые рассматривают этот тип опыления как расширенное самоопыление, а другие — как ограниченное перекрестное опыление.
Перекрестное опыление может быть абиотическим и биотическим. Преобладающим типом абиотического опыления является ветроопыление. У растений выработался ряд приспособлений к ветроопылению (анемофилии): цветение древесных растений до распускания листьев (ольха, орешник); длинные раскачивающиеся под действием ветра соцветия-сережки (ольха, орешник) или длинные тычинки с крупными пыльниками (злаки); невзрачные раздельнополые цветки без околоцветника (осоки) или с сильно редуцированным околоцветником (ситники); большое количество легкой пыльцы с гладкой экзиной (орешник, береза) и т. п.
Древним типом абиотического опыления является гидрофилия — опыление при помощи воды. Типичная гидрофилия характерна для водных (водяная звездочка — Callitriche, наяда — Najas, роголистник — Ceratophyllum) растений, у которых существуют специфические особенности пыльцы. У некоторых гидрофильных растений пыльца сильно вытянута (у Zosteraдлина ее 2500 мкм), что имеет важное приспособительное значение, так как такая пыльца очень подвижна и способна быстро распространяться в воде. Весьма существенно, что у всех гидрофильных растений пыльца при попадании в воду не лопается (что происходит с пыльцой наземных растений) и остается жизнеспособной. Гидрофилия по сравнению с анемофилией довольно редкий тип абиотического перекрестного опыления.
Биотическое опыление может осуществляться как различными насекомыми (энтомофилия) — пчелами, шмелями, дневными и ночными бабочками, осами, жуками, перепончатокрылыми, муравьями, так и позвоночными животными — птицами, летучими мышами, мелкими приматами. В процессе длительной эволюции у растений выработались определенные приспособления для привлечения беспозвоночных и позвоночных животных-опылителей. Такими приспособлениями являются цветки с крупным околоцветником, определенная окраска венчика, наличие специфических запахов, характерная форма цветков, часто имитирующая форму тела самок насекомых-опылителей, большое количество пыльцы, присутствие нектара и т. д. Посещение опылителями цветков различного типа связано с их пищевыми потребностями в различные периоды жизненного цикла.
Среди многих исследователей, к сожалению, распространено представление о самоопылении как о тупике эволюционного развития. Оно сложилось под влиянием идей Ч. Дарвина, который показал приспособления цветков покрытосеменных растений к перекрестному опылению и описал те из них, которые препятствуют самоопылению.
У перекрестноопыляемых растений действительно существуют специальные приспособления, препятствующие самоопылению. К ним относятся дихогамия и гетеростилия. При дихогамии наблюдается неодновременное созревание тычинок и пестиков, проявляется она в форме протерандрии (протандрия) (более раннее созревание тычинок) (рис. 189) и протерогинии (протогиния) (более раннее созревание пестиков) (рис. 190).
Рис. 189. Явление протерандрии у камнеломки жестколистной (Saxifraga aizoides): А—цветок с хорошо развитыми тычинками и недоразвитыми стилодиями; Б—цветок с хорошо развитыми стилодиями и засыхающими пыльниками тычинок
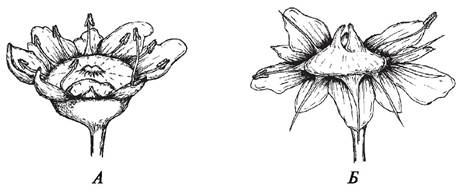
Рис. 190. Явление протерогинии у норичника узловатого (Scrophularia nodosa) (продольный разрез цветка): А—цветок с хорошо развитым столбиком и недоразвитыми тычинками; Б—цветок с хорошо развитыми тычинками и увядающим столбиком
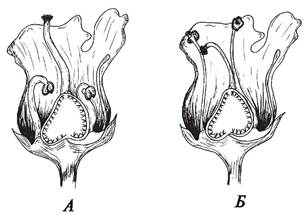
Протерандрия встречается чаще, чем протерогиния, и характерна для представителей семейств Зонтичные, Колокольчиковые, Кипрейные, Гераниевые и ряда других. Протерогиния наблюдается у многих крестоцветных, подорожниковых, у некоторых сложноцветных (дурнишник — Xanthium). При дихогамии разница во времени созревания тычинок и пестиков у разных видов варьирует от нескольких часов до нескольких дней.
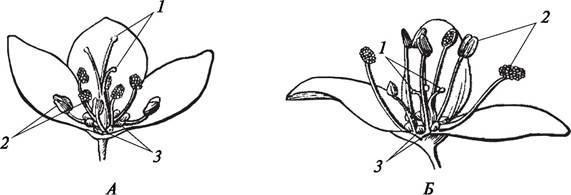
Гетеростилия — особый тип приспособлений, который встречается у представителей семейств Первоцветные, Гречишные, Гвоздичные, Бурачниковые. При гетеростилии в разных цветках одного и того же вида образуются тычинки и столбики пестиков, имеющие различную длину. В цветках с длинными тычинками пестики имеют короткий столбик (или короткие стилодии). В цветках с короткими тычинками столбик пестика длинный (или длинные стилодии). Оплодотворение происходит только в том случае, если пыльца цветков с длинными тычинками попадет на рыльце пестика, имеющего длинный столбик, или если пыльца с цветков с короткими тычинками попадает на рыльце пестика с коротким столбиком. В других случаях пыльца не прорастает, что связано с самонесовместимостью пыльцы и рыльца одного цветка (рис. 191).
Рис. 191. Явление гетеростилии у гречихи посевной (Fagopyrum sagittatum): А—цветок с длинными стилодиями и короткими тычинками; Б—цветок с короткими стилодиями и длинными тычинками: 1—стилодии; 2—тычинки; 3—нектарники
В настоящее время бесспорно установлено, что в процессе эволюции у покрытосеменных растений выработалось два способа опыления — перекрестное и самоопыление. Несмотря на детерминированность способов опыления, они весьма лабильны, подвижны и могут в зависимости от климатических условий в период цветения меняться, т. е. у самоопылителей может наблюдаться перекрестное опыление, а у перекрестноопыляемых растений — самоопыление. Эволюционное значение такой подвижности имеет большое значение. При свободном переопылении внутри больших популяций очень затруднено закрепление новых комбинаций скрещивания. Любая удачная комбинация скрещивания при свободном переопылении будет распадаться и не сможет удержаться в потомстве. Самоопыление у перекрестников способствует изоляции новых форм, закреплению их в последующих поколениях, а значит, способствует видообразованию. Перекрестное опыление, которое время от времени наблюдается у самоопылителей, повышает уровень гетерозиготности потомства, а, следовательно, и жизнеспособность вида.
8.1.1. Прорастание пыльцы на рыльце пестика
Пыльца, попав тем или иным способом на рыльце пестика, прикрепляется к нему и начинает прорастать. Прорастанию пыльцы на рыльце пестика предшествует ряд физиолого-биохимических и цитологических процессов: выделение липидных капель на поверхности пыльцевого зерна, поступление воды из тканей пестика в пыльцу, активизация ферментов, перераспределение в цитоплазме всех компонентов пыльцевого зерна. Первым признаком того, что пыльца начала прорастать, т. е. формировать пыльцевую трубку, является циклоз — круговое вращательное движение цитоплазмы пыльцевого зерна. При прорастании пыльцы интина выпячивается через одну из апертур и образует пыльцевую трубку, в которую постепенно перемещается все содержимое пыльцевого зерна: цитоплазма вегетативной клетки с ядром, всеми органоидами и генеративной клеткой (если пыльца двуклеточная) или двумя спермиями (если пыльца трехклеточная) (рис. 192).
Рис. 192. Прорастающее пыльцевое зерно (пыльца) бобов обыкновенных (Vicia faba): 1—пыльцевое зерно; 2—пыльцевая трубка; 3—ядро вегетативной клетки; 4— генеративная клетка
Пыльцевая трубка внедряется в рыльце, затем проникает в столбик и начинает по нему продвигаться в сторону завязи. Столбик имеет различное строение. Он может быть открытым, закрытым, полуоткрытым. В зависимости от этого пыльцевая трубка продвигается по столбику по-разному. В открытом столбике она скользит по стенке канала. В закрытом и полуоткрытом пыльцевая трубка проходит, разрушая выстилающие столбик железистые клетки, и переходит на стенку завязи. Далее пыльцевая трубка движется по стенке завязи, пока не приблизится к месту расположения семяпочки.
Пыльцевая трубка растет по градиенту концентрации ростовых веществ, а также ионов кальция. Однако, вполне вероятно, что существуют и иные хемотропические факторы, которые способствуют продвижению пыльцевой трубки к семяпочке. Так как при продвижении по столбику и стенкам завязи пыльцевая трубка тесно контактирует с тканями материнского организма, очевидно, что между ними устанавливается какое-то специфическое физиологическое взаимодействие, которое пока еще не изучено.
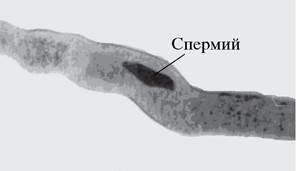
У двудольных растений, для которых характерна двуклеточная пыльца, в пыльцевой трубке генеративная клетка делится митотически и образует два спермия (рис. 193). Если пыльцевая трубка широкая, митоз происходит типично, при митотическом делении ядра генеративной клетки четко вырисовываются все стадии митоза: профаза, метафаза, анафаза, телофаза. В узких пыльцевых трубках не образуется типичной метафазной пластинки. За митотическим делением ядра следует цитокинез. Таким образом, спермии являются гаплоидными клетками, имеющими, кроме плазмалеммы, клеточные оболочки. Цитоплазма спермиев занимает в клетке боковое положение, а также сконцентрирована на полюсах. Ядро не имеет ядрышка, а хроматин ядра находится в конденсированном состоянии. В типичном случае в цитоплазме спермиев содержится характерный для растительных клеток набор органоидов: митохондрии, диктиосоммы, гранулярный эндоплазматический ретикулум, рибосомы, микротрубочки. Имеются также небольшие вакуоли и пластиды. У некоторых злаков, сложноцветных отмечен диморфизм спермиев, который проявляется не только в различии формы, размеров, но и в содержании органелл.
Рис. 193. Спермий в пыльцевой трубке бобов обыкновенных (Vicia faba)
Вопрос о движении генеративной клетки и спермиев в пыльцевой трубке является проблематичным, а представления различных авторов о способах движения — противоречивыми. Одни авторы считают, что генеративная клетка и спермии, независимо от места их образования, движутся активно, другие указывают, что они передвигаются пассивно, увлекаемые током цитоплазмы. Не ясна и функция вегетативного ядра. Некоторые авторы на основании того, что оно всегда располагается в растущем конце пыльцевой трубки, считают, что вегетативное ядро регулирует рост пыльцевой трубки, которая обладает специфической поляризацией. По мере того как пыльцевая трубка удлиняется, пыльцевое зерно (пыльца) и основание пыльцевой трубки опустошаются, и живое содержимое отделяется от пустого участка каллозной пробкой. Таким образом сохраняется целостность мужского гаметофита. Для пыльцевой трубки характерен только апикальный рост, растущая зона отделяется от остальной части пыльцевой трубки все новыми и новыми каллозными пробками.