МОРФОЛОГИЯ РАСТЕНИЙ - Т. А. Сауткина - 2012
ГЛАВА 8. ОПЫЛЕНИЕ И ОПЛОДОТВОРЕНИЕ ЦВЕТКОВЫХ РАСТЕНИЙ. ОБРАЗОВАНИЕ ЗАРОДЫША И ЭНДОСПЕРМА
8.2. Процесс двойного оплодотворения покрытосеменных растений
Промежуток времени между процессами опыления и оплодотворения у различных представителей покрытосеменных растений различен. Самый короткий промежуток времени между опылением и оплодотворением обнаружен у представителей семейства Сложноцветные, он варьирует от 15—30 мин, как, например, у кок-сагыза (Taraxacum kok-saghyz), до 1,5 ч у большинства видов. У некоторых представителей семейства Березовые этот период колеблется от 1 до 4 мес., а у буковых (некоторые виды дуба) — от 12 до 14 мес.
Пыльцевая трубка чаще всего направляется к микропилярному полюсу семяпочки, однако она может войти в семяпочку и через другие участки. Если пыльцевая трубка входит в семяпочку через микропиле, такой способ внедрения называется порогамией, через халазу — халазогамией, через латеральные зоны интегументов и нуцеллуса — мезогамией. Самым древним типом внедрения пыльцевой трубки в семяпочку считается порогамия, она характерна не только для покрытосеменных, но и для голосеменных растений. Хала- зогамия и мезогамия — это специализированные типы вхождения пыльцевой трубки, которые были обнаружены и описаны у ряда древесных растений (казуарины, березы, ореха). Однако каким бы способом не проникла пыльцевая трубка в семяпочку, она «блуждает» по тканям семяпочки, пока не окажется на микропилярном полюсе вблизи микропиле.
Пыльцевая трубка входит в зародышевый мешок через одну из синергид, которая при этом разрушается. Верхушка пыльцевой трубки располагается вблизи яйцевого аппарата и здесь лопается. Содержимое пыльцевой трубки изливается вблизи яйцеклетки. Цитоплазма пыльцевой трубки сливается с цитоплазмой зародышевого мешка, расположенной возле яйцевого аппарата. Ядро вегетативной клетки разрушается, а два спермия некоторое время лежат в цитоплазме вблизи яйцеклетки, а затем начинают расходиться. Один спермий движется в сторону яйцеклетки, а второй — к полярным ядрам. Характер передвижения спермиев в зародышевом мешке окончательно не выяснен. С. Г. Навашин считал, что спермии обладают свойством энантиоморфизма, т. е. они зеркально подобны. Именно благодаря зеркальному подобию спермии актив - но отталкиваются друг от друга и движутся в полости зародышевого мешка.
Иных взглядов придерживалась Е. Н. Герасимова- Навашина. Согласно ее представлениям, спермии попадают в зародышевый мешок в состоянии незаконченного митотического деления, и именно остаточная митотическая активность способствует самостоятельному передвижению спермиев. Имеются и другие предположения.
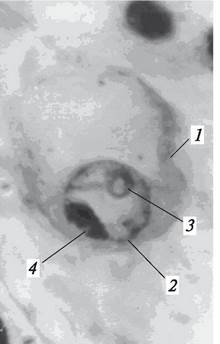
Процесс оплодотворения начинается со слияния спермия с яйцеклеткой. Это слияние происходит следующим образом. Спермий внедряется в цитоплазму яйцеклетки и здесь «сбрасывает» свою цитоплазму, которая смешивается с цитоплазмой яйцеклетки. Далее голое ядро спермия движется в сторону ядра яйцеклетки, внедряется в него (рис. 194) и сливается с ним. Только после того как произойдет слияние ядра спермия с ядром яйцеклетки, процесс оплодотворения яйцеклетки (амфимиксис) и образования зиготы можно считать завершенным. В ядре зиготы восстанавливается диплоидный набор хромосом.
Рис. 194. Этап двойного оплодотворения: спермий в ядре яйцеклетки: 1—яйцеклетка; 2—ядро яйцеклетки; 3—ядрышко ядра яйцеклетки; 4—спермий
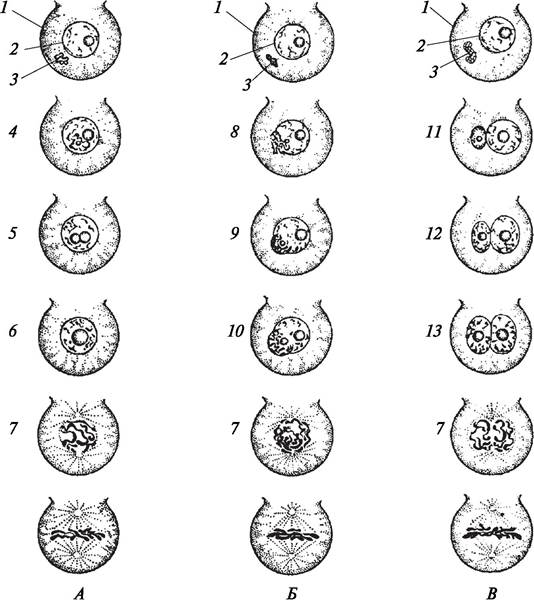
Различают три типа оплодотворения яйцеклетки: премитотический, постмитотический и промежуточный (рис. 195). При премитотическом типе оплодотворения ядра половых клеток сливаются, будучи в покоящемся состоянии. Деление ядра зиготы начинается после выхода из состояния покоя. При постмитотическом типе оплодотворения сливающиеся ядра яйцеклетки и спермия находятся в активном состоянии, т. е. в них начинается процесс митотического деления, который завершается слиянием ядер, и ядро зиготы фактически находится в профазе митоза. При промежуточном типе оплодотворения слияние происходит, когда в одном из ядер, чаще всего в ядре спермия, начинается процесс деления, а ядро яйцеклетки находится в состоянии покоя. Деление ядра зиготы продолжается после слияния ядер.
Рис. 195. Типы слияния яйцеклетки со спермием (типы оплодотворения по Герасимовой-Навашиной, 1961, с дополнениями): А—премитотический; Б—промежуточный; В —постмитотический: 1—яйцеклетка; 2—ядро яйцеклетки с ядрышком; 3—спермий в цитоплазме яйцеклетки; 4—начало митоза спермия, предшествующее слиянию с покоящимся ядром яйцеклетки; 5—слияние спермия с ядром яйцеклетки (амфимиксис); 6—образование зиготы; 7—начало митоза ядра зиготы; 8—вхождение спермия в состоянии митоза в ядро яйцеклетки; 9—начало митоза в ядре яйцеклетки; 10—слияние спермия и ядра яйцеклетки в состоянии митоза, образование зиготы; 11— спермий и ядро яйцеклетки в покоящемся состоянии; 12, 13—начало слияния делящихся ядер спермия и яйцеклетки, образование зиготы
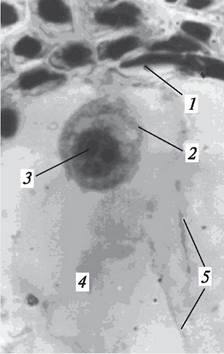
После оплодотворения меняется форма оплодотворенной яйцеклетки. Если яйцеклетка имеет грушевидную форму, то зигота — округлую (рис. 196). В яйцеклетке ядро с одним ядрышком, в зиготе ядро может содержать несколько ядрышек. Цитологи считают, что по числу ядрышек в ядре зиготы можно судить о количестве спермиев, проникших в яйцеклетку.
Рис. 196. Зигота в полости зародышевого мешка: 1—разрушающиеся клетки нуцеллуса; 2—зигота; 3—ядро зиготы; 4—полость зародышевого мешка; 5—тяжи цитоплазмы зародышевого мешка
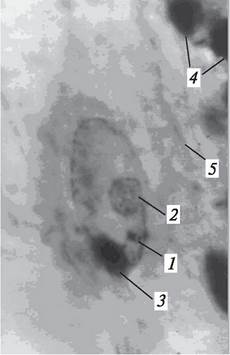
Второй спермий подходит к полярным ядрам или к центральному ядру (ядру центральной клетки) зародышевого мешка и начинает сливаться с ними (рис. 197). Процесс слияния может проходить по - разному. Если полярные ядра обособлены, сначала спермий может оплодотворить микропилярное ядро, а затем слиться с халазальным. Порядок слияния может быть и иным: спермий может войти в халазальное ядро, а потом оплодотворенное ядро сливается с микропилярным. Если до оплодотворения образуется центральное ядро зародышевого мешка, процесс слияния спермия и центрального ядра зародышевого мешка протекает подобно тому, как проходит оплодотворение яйцеклетки. Таким образом, после оплодотворения полярных ядер или центрального ядра зародышевого мешка образуется полиплоидное первичное ядро эндосперма или полиплоидная (в простейшем случае триплоидная) первичная клетка эндосперма с первичным ядром. В некоторых зародышевых мешках отклоняющихся типов первичное ядро эндосперма является высокополиплоидным. Так, в зародышевом мешке Ререromiа-munа содержится от 7 до 13 поляр - ных ядер, после оплодотворения которых образуется очень крупное первичное ядро эндосперма, содержащее от 8n до 14n.
Рис. 197. Этап процесса двойного оплодотворения: спермий в центральном ядре зародышевого мешка:1—центральное ядро зародышевого мешка (ядро центральной клетки зародышевого мешка); 2—ядрышко ядра центральной клетки зародышевого мешка; 3—спермий; 4—разрушающиеся клетки нуцеллуса; 5—тяжи цитоплазмы зародышевого мешка
Явление оплодотворения у покрытосеменных было открыто профессо - ром Киевского университета С. Г. Навашиным в 1898 г. и названо им процессом двойного оплодотворения.Это открытие послужило толчком к изучению данного события у покрытосеменных, особенно у тех, которые имеют большое практическое значение.
После оплодотворения зигота и первичное ядро эндосперма впадают в состояние покоя, а затем начинается их развитие. Из зиготы образуется зародыш
нового организма, а из первичного ядра (или первичной клетки) эндосперма особая питательная ткань — эндосперм. Процесс, при котором зародыш развивается из зиготы, возникшей в результате оплодотворения, называется амфимиксисом. Однако у ряда растений зародыш семени развивается без оплодотворения — апомиктично.