МОРФОЛОГИЯ РАСТЕНИЙ - Т. А. Сауткина - 2012
ГЛАВА 8. ОПЫЛЕНИЕ И ОПЛОДОТВОРЕНИЕ ЦВЕТКОВЫХ РАСТЕНИЙ. ОБРАЗОВАНИЕ ЗАРОДЫША И ЭНДОСПЕРМА
8.3. Понятие об апомиксисе
Термин «апомиксис» был введен в 1906 г. Г. Винклером для обозначения, как первоначально считалось, размножения, не связанного со слиянием мужской и женской гамет (процессом оплодотворения), но происходящего при участии семян.
Как показали дальнейшие исследования эмбриологов, апомиксис нельзя рассматривать как процесс, связанный с аномалией оплодотворения (М. П. Солнцева, 1991). При этом способе семенного размножения обязательно происходит слияние спермия с полярными ядрами (или с ядром центральной клетки), а в некоторых случаях при апомиктичном образовании семян происходит проникновение спермия в яйцеклетку, хотя не наблюдается слияния с ее ядром. Следовательно, апомиксис нельзя называть бесполо-семенным размножением (термин С. С. Хохлова, 1946), как это делается в ряде работ. Согласно современной терминологии, апомиксис — это особый тип семенного размножения, при котором наблюдаются различные отклонения от типично протекающего макроспорогенеза и полового процесса или обоих этих процессов.
Явление апомиксиса с давних пор привлекало к себе внимание различных исследователей, которые пытались объяснить причины происхождения этого явления, а также дать его классификацию. Несмотря на то что до настоящего времени предложен ряд классификаций типов апомиксиса, практически ни одна из них не является общепризнанной.
По имеющимся представлениям, апомиксис может быть постоянным (наследственным) и случайным (ненаследственным); в зависимости от особенностей макроспорогенеза — нередуцированным (диплоидным) и редуцированным (гаплоидным). Термины «нередуцированный» и «редуцированный» апомиксис обозначают категории апомиксиса и используются традиционно.
В современную классификацию апомиксиса М. П. Солнцева (1991) вводит также понятия «тип апомиксиса» и «форма апомиксиса». В основу выделения типов апомиксиса положен характер нарушений полового процесса. М. П. Солнцева выделяет шесть типов апомиксиса: гемигамия, андрогенез, псевдогамия, партеногенез, апогаметия, адвентивная эмбриония.
Гемигамия — это тип апомиксиса, при котором не происходит слияния ядер спермия и яйцеклетки. Ядро спермия, находящееся в цитоплазме яйцеклетки, не способно слиться с ее ядром, но остается жизнеспособным, стимулирует деление ядра яйцеклетки и само также делится. В результате формируется
зародыш, у которого часть клеток материнского происхождения, а часть — отцовского, что приводит к развитию химерного зародыша. Таким образом, при гемигамии нарушение полового процесса происходит на самом последнем этапе — этапе оплодотворения. Впервые гемигамия была описана у рудбекии, рассеченной (Rudbeckia laciniata) Е. Батталия в 1946 г.
Андрогенез — такой тип апомиксиса, при котором зародыш развивается из яйцеклетки, ядро которой элиминируется и заменяется ядром спермия. Поэтому клетки зародыша имеют ядра отцовского происхождения, а развившиеся из таких зигот растения имеют признаки отцовского вида (андрогинное потомство). Андрогенез не обнаружен эмбриологами растений, но хорошо доказан генетиками. Он описан у скерды кровельной (Crepis tectorum), табака обыкновенного (Nicotiana tabacum).
Псевдогамия — тип апомиксиса, при котором спермий проникает в цитоплазму яйцеклетки, но не способен продвигаться дальше и погибает. В то же время он каким-то образом стимулирует развитие яйцеклетки, в результате чего формируются особи с материнским типом наследственности (матроклинное потомство).
При партеногенезе зародыш развивается или до момента опыления (часто говорят «без оплодотворения»), поэтому развивающиеся дочерние организмы всегда обладают материнским типом наследственности (гиногенез) или после опыления, тогда спермий проникает в яйцеклетку, но не сливается с ней, а занимает ее место, так как яйцеклетка погибает (андрогенез). Впервые редуцированный партеногенез, при котором формировались гаплоидные растения материнского типа, был получен у дурмана обыкновенного (Daturastramonium) при действии на растение низкой температурой. Нередуцированный партеногенез характерен для лапчаток, мятликов и ряда других растений. Андрогенез отмечен у некоторых видов табака, перца, скерды.
Апогаметия — тип апомиксиса, при котором зародыш развивается не из зиготы, а из синергид (синергидная апогаметия) или антипод (антиподная апогаметия). Синергидная апогаметия характерна для многих орхидных и встречается значительно чаще, чем антиподная, обнаруженная у некоторых видов ястребинок. Зародыши, возникшие из синергид или антипод, меньших размеров, чем образовавшиеся из зиготы.
Адвентивная эмбриония — тип апомиксиса, при котором развитие зародыша начинается в тканях нуцеллуса или интегументов, а затем он внедряется в типичный зародышевый мешок, где и проходит его дальнейшее развитие. Адвентивная эмбриония описана более чем у 200 видов, относящихся к 47 семействам, стоящим на разных ступенях филогенетического развития (М. П. Солнцева, 1991).
Таким образом, тип апомиксиса определяется эмбриологическими особенностями.
Форма апомиксиса обусловлена разным ходом мейоза, что фактически определяет характер макроспорогенеза (мегаспорогенеза). Различают следующие формы апомиксиса: эуспорию, гемиэуспорию, анеуспорию и апоспорию.
При эуспорической форме апомиксиса макроспорогенез протекает типично, без нарушений мейоза, апомиксис осуществляется на гаплоидном уровне. Аномалии, приводящие к появлению диплоидных элементов зародышевого мешка, могут возникать в процессе формирования женского гаметофита.
При гемиэуспорической форме апомиксиса первый этап мейоза (мейоз-1) проходит правильно, а второй этап (мейоз-2) выпадает. Редупликация хромосом происходит во время длительной интерфазы мейоза-1.
Анеуспорическая форма апомиксиса наблюдается в случае различных нарушений мейоза, вследствие чего происходит восстановление диплоидного числа хромосом.
Апомиксис может развиваться на фоне апоспории, при которой зародышевый мешок развивается не из макроспоры, а из клеток интегументов или нуцеллуса.
Согласно предложению, М. П. Солнцевой, при классификации апомиксиса следует учитывать оба фактора — и форму, и тип апомиксиса (например, эуспорический партеногенез; гемиэуспорическая апогаметия и т. д.). При эуспорических формах апомиксиса только партеногенез происходит без опыления, всем остальным типам апомиксиса предшествует опыление. При геми- эуспории, анеуспории и апоспории такие типы апомиксиса, как гемигамия, андрогенез, псевдогамия, всегда возникают на фоне опыления. Партеногенез и апогаметия при этих формах апомиксиса могут формироваться как при наличии опыления, так и без опыления.
Апомиксис широко распространен в природе и используется в селекции растений и семеноводстве (Д. Ф. Петров, 1988). Апомикты, как правило, преобладают в районах с суровыми условиями обитания. Растения с апомиктичным способом развития зародыша обладают рядом весьма ценных качеств. Они устойчивы к неблагоприятным условиям, характеризуются интенсивным семенным и вегетативным размножением, мощным развитием. Апомиксис как особый способ образования зародыша семени имеет приспособительный характер и не предопределен эволюцией жизненного цикла развития.
8.4. Начальные этапы формирования эндосперма и зародыша
Как было указано ранее, после процесса оплодотворения зигота и первичное ядро эндосперма впадают в состояние покоя, а затем начинается их развитие.
Первым начинает делиться ядро первичной клетки (или первичное ядро) эндосперма (рис. 198). В результате первого митотического деления образуется два ядра, одно из которых перемещается к микропилярному полюсу и располагается вблизи зиготы, а второе мигрирует в область халазы. В дальнейшем деление ядер эндосперма в микропилярной и халазальной зонах зародышевого мешка происходят не синхронно. Более интенсивно делятся ядра в микропилярной зоне, в халазальной темп делений замедлен. Это отражается на величине ядер, они в халазальной зоне крупнее, чем в микропилярной. Кроме того, у некоторых растений, в частности у бобовых, деление ядер в микропилярной и халазальной зонах может происходить по-разному. Вблизи зиготы ядра делятся только митотически. В халазальной зоне наряду с митозом часто наблюдается и амитотическое деление ядер. Амитоз высокоспециализированный тип деления, который не выводит ядро из функционально активного состояния. Это имеет большое биологическое значение, так как именно через халазальную часть семяпочки в зародышевый мешок поступают питательные вещества, которые активно накапливаются в эндосперме.
Рис. 198. Первый этап развития эндосперма у бобов: митотическое деление первичного ядра эндосперма: 1—клетки нуцеллуса; 2—ядрышко центрального ядра зародышевого мешка; 3—первичное ядро эндосперма; 4—полость зародышевого мешка; 5—тяжи цитоплазмы в полости зародышевого мешка
В зависимости от того, происходит ли на ранних стадиях формирования эндосперма цитокинез или нет, различают три типа эндосперма: нуклеарный (ядерный), целлюлярный (клеточный) и гелобиальный (промежуточный) (рис. 199).
Рис. 199. Типы эндосперма: А—нуклеарный (горичник горный — Peucedanum oreoselinum); Б—гелобиальный (рдест плавающий — Potamogeton natans); В—целлюлярный (вахта трехлистная — Menyanthes trifoliata): 1—ядра эндосперма в цитоплазме зародышевого мешка; 2—микропилярная камера зародышевого мешка с нуклеарным эндоспермом; 3—халазальная камера с одной клеткой эндосперма; 4—клетки эндосперма в полости зародышевого мешка; 5—развивающийся предзародыш; 6—клетки нуцеллуса
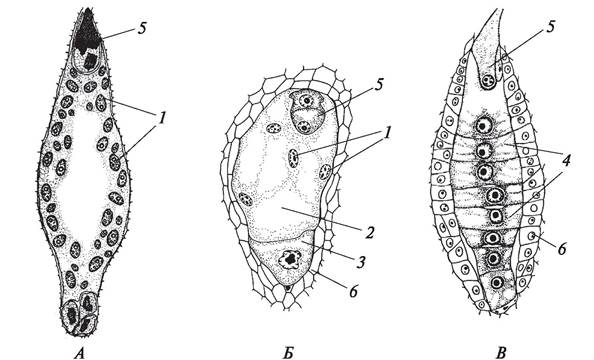
При нуклеарном типе развития эндосперма деление ядер длительное время не сопровождается образованием клеточных перегородок и ядра свободно располагаются в тяже цитоплазмы. Такой тип эндосперма характерен для многих представителей класса Двудольные. Впоследствии нуклеарный эндосперм или дегенерирует и в зрелых семенах отсутствует, как у бобовых, или становится клеточным и принимает типичное для запасающей ткани строение.
При формировании целлюлярного эндосперма каждое митотическое деление ядра сопровождается цитокинезом. Так формируется эндосперм злаков.
Гелобиальный тип развития эндосперма выявлен у однодольных из семейств Частуховые, Сусаковые, Водокрасовые, которые раньше объединялись в порядок Водолюбы — Helobiales. (В настоящее время этот порядок упразднен.) После первого деления ядра эндосперма при гелобиальном типе развития образуется клеточная перегородка, которая делит зародышевый мешок на две неравные камеры: маленькую халазальную и крупную микропилярную. В халазальной камере деление ядер больше не происходит или наблюдается всего несколько делений, а затем ядра дегенерируют. В микропилярной камере ядра делятся путем митоза, каждое деление сопровождается цитокинезом, и развитие эндосперма в дальнейшем идет по целлюлярному типу.
Постепенно по мере развития в клетках эндосперма накапливаются запасные питательные вещества (крахмал, белки, жиры), но уменьшается количество воды, и он становится физиологически мертвой тканью.
Первое деление ядра зиготы начинается только после того, как в зародышевом мешке образуется не менее четырех ядер эндосперма. Ядро зиготы делится митотически. Каждое деление ядра сопровождается заложением клеточных перегородок. В результате делений число клеток — производных зиготы — возрастает и образуется двуклеточный (рис. 200), а затем четырехклеточный предзародыш (рис. 201).
Рис. 200. Митотическое деление ядра зиготы и образование двуклеточного предзародыша: 1—базальная клетка; 2—терминальная клетка
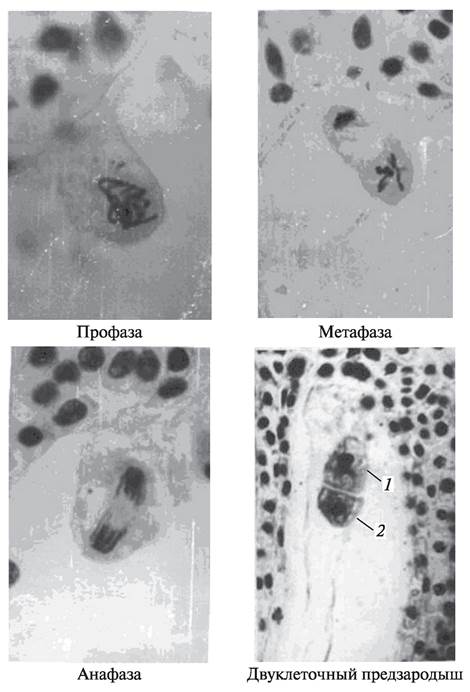
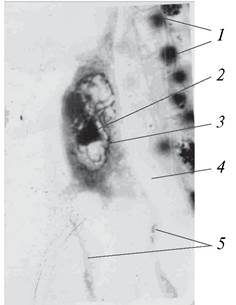
Рис. 201. Четырехклеточный предзародыш бобов обыкновенных (Vicia faba) в полости зародышевого мешка: 1—клетки нуцеллуса; 2—полость зародышевого мешка
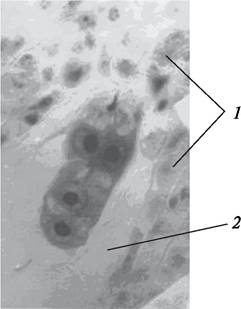
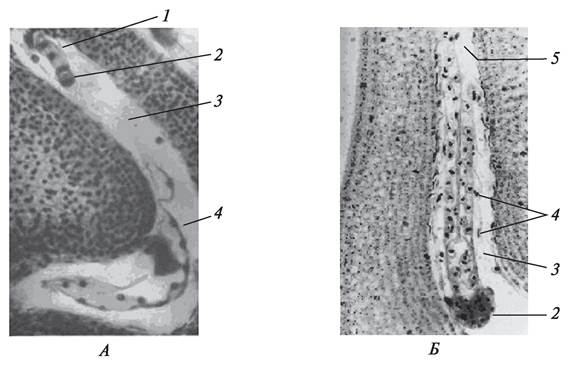
Однако дифференцировка на вегетативные органы не наблюдается до тех пор, пока не сформируется подвесок (суспензор) и не образуется шаровидная масса клеток, которая затем и даст начало самому зародышу. Подвесок имеет различное строение, что особенно хорошо изучено у бобовых (рис. 202).
Рис. 202. Развивающийся зародыш и нуклеарный эндосперм бобов обыкновенных (Vicia faba): А—формирующийся предзародыш и подвесок через 48 часов после опыления; Б—многоклеточный предзародыш и сформированный подвесок через 96 часов после опыления: 1—формирующийся подвесок; 2—шаровидный предзародыш;
3—полость зародышевого мешка; 4—нуклеарный эндосперм; 5—сформированный подвесок из четырех многоядерных клеток
После процесса двойного оплодотворения разрастается семяпочка и постепенно превращается в семя. Пестик также претерпевает существенные изменения и образует плод. Семена и плоды — это конечные продукты тех сложных репродуктивных процессов, которые происходят в цветке.
С образованием плода цветок как репродуктивный орган исчерпывает свои функции и перестает существовать. Таким образом, цветок — это кратковременно существующий репродуктивный орган покрытосеменных растений.