МОРФОЛОГИЯ РАСТЕНИЙ - Т. А. Сауткина - 2012
ГЛАВА 2. КРАТКАЯ ИСТОРИЯ ИЗУЧЕНИЯ КЛЕТКИ. ОСОБЕННОСТИ СТРОЕНИЯ КЛЕТКИ ВЫСШИХ РАСТЕНИЙ
2.1. Клетка как объект исследования
В настоящее время ни у кого не вызывает сомнения, что все живые организмы состоят из клеток. Клетка — это структурный элемент растений и животных.
Изучение клетки начинается с момента ее открытия. В 1665 г. английский естествоиспытатель Р Гук (1635—1703), исследуя под микроскопом тонкие срезы пробки, увидел картину, напоминающую пчелиные соты. Эти ячейки, отверстия или поры, по выражению Р Гука, он назвал сеllulа — клетка (рис. 1). Наблюдения Р Гука были продолжены М. Мальпиги и Н. Грю, однако они обращали внимание только на форму клеток, не пытаясь выяснить внутреннее строение. Анатомические исследования растений в ХVII—ХVIII вв. были немногочисленными и фактически не вносили ничего нового в представление о клетке по сравнению с представлениями Р. Гука, М. Мальпиги и Н. Грю.
Рис. 1. Изображение срезов пробки (из «Микрографии» Р. Гука, 1665)

Сведения о строении клетки длительное время были отрывочными и примитивными. Практически все ботаники XVII — начала XVIII в. представляли клетки как пустоты в растительной ткани, имеющие общие стенки, или как пузырьки.
Только в 1812 г. немецкий ботаник П. Мольденгауер (1766—1827) применил метод мацерации растительных тканей, получил изолированные клетки и показал, что каждая клетка имеет собственную оболочку. Дальнейшее развитие представление о клетке получило в работе немецкого ботаника Ф. Мейена (1804—1830) «Фитотония», вышедшей в 1830 г. Растительную клетку Ф. Мейен характеризовал как пространство, полностью замкнутое и окруженное растительной мембраной.
С середины XIX в. начинается изучение внутреннего строения клетки. В 1839 г. чешский ботаник Ян Пуркинье (1787—1869) обнаружил содержимое в животных клетках и назвал его протоплазмой, а в 1846 г. немецкий ботаник Гуго фон Моль (1805—1872) перенес этот термин на содержимое живых растительных клеток. Тем самым было подтверждено наблюдение Р. Гука, считавшего, что клетка не пустая, а в ней есть содержимое, которое Р. Гук называл соком. Термин «протоплазма» впоследствии несколько видоизменялся в зависимости от того, какой смысл вкладывали в него исследователи. Так, И. Ганштейн (1822—1880) содержимое одной клетки (без наружной оболочки) называл протопластом. Э. Страсбургер (1844—1912) в 1882 г. предложил для растительных клеток термин «цитоплазма» — обязательная часть клетки, заключенная между плазматической мембраной и ядром.
Постепенно ученые выяснили, что клетка имеет сложное внутреннее строение. Еще в 1676 г. А. Левенгук (1632—1723) фактически обнаружил в клетке пластиды, но назвал их кристаллами. Важное открытие сделал в 1831 г. Р. Броун (1773—1858). При изучении эпидермиса орхидных он дал подробное описание ядра, а в 1842 г. М. Шлейден (1804—1881) впервые в составе ядра обнаружил ядрышки. В 1882 г. В. Флемминг (1843—1905) в клетках животных обнаружил митохондрии, а в 1904 г. Ф. Мёвес (1878—1923) выявил их в клетках растений. В 1846 г. Гуго фон Моль доказал, что протоплазма растительных клеток обладает способностью к самостоятельному движению. Наблюдения Гуго фон Моля подтвердил в 1850 г. Ф. Кон (1828—1898), а в 1854 г. Н. Принсгейм (1823—1894).

Неоднократно делались попытки изучить образование клеток. Так, в 1835 г. Моль описал деление клеток у водоросли кладофора, а в 1838 г. — деление клеток у ряда высших растений, из которых впоследствии формировались замыкающие клетки устьиц. В 1841 г. австрийский ботаник Ф. Унгер (1800—1870) наблюдал деление клеток в точке роста растений. Правильное представление о делении клеток (с позиций сегодняшнего времени) имели такие ботаники, как Н. И. Железнов, К. Негели (1817—1891), немецкий врач и биолог Р. Реман (1815—1865), немецкий гистолог и эмбриолог А. Келликер (1817—1905), русский зоолог, профессор Московского университета Н. А. Варнек (1821—1876) и др. Однако наряду с верными, экспериментально полученными данными об образовании клетки путем деления существовал ряд неверных мнений об этом процессе. Это касалось и представлений таких признанных в области ботаники и зоологии авторитетов, как М. Шлейден и Т. Шванн (1810—1882). Конец всем неправильным суждениям положило меткое высказывание известного немецкого патолога Р. Вирхова (1821—1902): Omnis cellula e cellula, т. е. каждая клетка происходит только из клетки.

Таким образом, к середине XIX в. уже имелись вполне конкретные представления о строении клетки и в различной форме высказывались предположения, что клетка является основой организации растений и животных. Особенно четко это было сформулировано в клеточной теории, которую Ф. Энгельс наряду с законом превращения энергии и эволюционной теорией Ч. Дарвина назвал одним из трех великих открытий XIX в. Основная заслуга оформления клеточной теории принадлежит Т. Шванну (1839), который использовал собственные данные, а также результаты исследований М. Шлейдена и других ученых. Суть клеточной теории состоит в бесспорном признании, что все животные и растительные организмы состоят из клеток. Единство клеточного строения животных и растительных организмов находит подтверждение как в сходстве строения различных клеток, так и в сходстве их химического состава, а также метаболических процессов.
Однако, несмотря на наличие общих закономерностей в строении растительных и животных клеток, клетки высших растений имеют ряд специфических особенностей. К таким особенностям относятся наличие прочной клеточной оболочки (клеточной стенки), а также пластид и вакуолей.
2.2. Строение клеточной оболочки
Клеточная оболочка (стенка) растительных клеток представляет собой сложное образование, расположенное над плазмалеммой — полупроницаемой мембраной, которая ограничивает цитоплазму клетки и отделяет ее от оболочки. Клеточная оболочка — типичный компонент растительной клетки. Благодаря наличию прочной клеточной оболочки сохраняется определенная форма клеток, а протопласт клетки оказывается надежно защищенным.
Оболочка растительной клетки имеет двухкомпонентную структуру. Она состоит из кристаллической основы и матрикса. Скелетную, или кристаллическую, основу клеточной оболочки составляет целлюлоза, или клетчатка. Матрикс содержит гемицеллюлозы, пектиновые вещества, особый структурный белок (экстенсин) и является насыщенным водой пластичным гелем.
Целлюлоза — основной опорный полисахарид клеточных оболочек растений. В химическом отношении она представляет собой в-1,4-Д-глюкан с эмпирической формулой (С6Н10О5)n. Молекулы целлюлозы состоят из остатков D-глюкозы, соединенных β-1,4-гликозидными связями. Количество остатков глюкозы в молекулах целлюлозы варьирует в широких пределах (от 300—500 в искусственных вискозных волокнах до 10 000—14 000 в лубяных волокнах), что определяет степень полимеризации ее молекул. Макромолекулы целлюлозы — линейные неразветвленные цепи, которые могут достигать в длину нескольких микрометров, но имеют в толщину всего около 8 нм, что делает их невидимыми даже в электронный микроскоп. Благодаря высокой степени полимеризации молекул целлюлоза не растворяется в воде, органических растворителях, а также в слабых растворах кислот и щелочей (рис. 2). В оболочке растительной клетки молекулы целлюлозы составляют сложные агрегации. Приблизительно 40—60 макромолекул целлюлозы, располагаясь параллельными тяжами, образуют мицеллы за счет водородных связей. Молекулы целлюлозы в мицеллах связаны таким образом, что формируют кристаллоподобные структуры. Благодаря этому для мицелл (часто говорят, для клеточной оболочки) характерно свойство анизотропии, что обеспечивает двойное лучепреломление. Существование мицелл установлено с помощью рентгеноструктурного анализа.
Рис. 2. Молекула целлюлозы: а — строение; б — схема межмолекулярных водородных связей
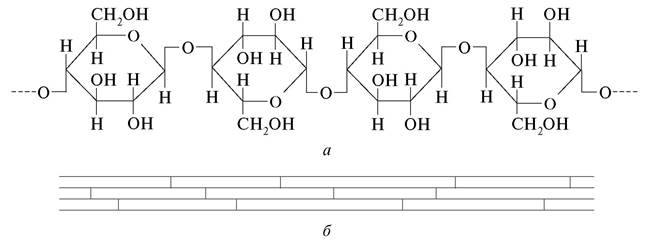
Мицеллы образуют микрофибриллы, или мицеллярные пучки. В состав каждой микрофибриллы входит около 160 мицелл. Толщина микрофибрилл 35— 100 нм, а длина достигает 500—600 нм. Несколько сотен микрофибрилл составляют фибриллы. Фибриллы — довольно массивные образования диаметром от 300 до 1500 нм, что позволяет их видеть с помощью светового микроскопа (рис. 3).
Рис. 3. Схема молекулярного строения первичной клеточной оболочки (стенки)
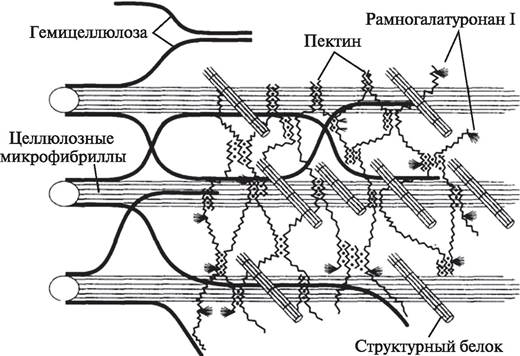
Фибриллы погружены в матрикс. Основу матрикса клеточной оболочки составляют гемицеллюлозы и пектиновые вещества. Гемицеллюлозы — группа полисахаридов, молекулы которых образованы остатками различных сахаров. Чаще всего они представлены остатками молекул гексозных сахаров (глюкоза, галактоза, манноза, фруктоза) и имеют эмпирическую формулу, аналогичную формуле целлюлозы, (C6H10O5)n. Однако в отличие от молекул целлюлозы молекулы таких гемицеллюлоз (гексозанов) менее полимеризированы, они включают не более 200 (от 50 до 200) моносахаридных остатков. Гемицеллюлозы могут быть образованы также остатками пентозных сахаров (рибоза, арабиноза), и тогда их формула — (C5H8O4)n. Молекулы гемицеллюлоз могут быть линейными или разветвленными. Особенности строения гемицеллюлоз отражаются на их химических и физических свойствах. В отличие от целлюлозы, которая не способна впитывать воду, гемицеллюлозы способны набухать в воде, но, как и целлюлоза, не растворяются в ней и органических растворителях. В то же время гемицеллюлозы растворяются в слабых растворах щелочей и легко гидролизуются в кислотах.
Пектиновые вещества — кислые высокомолекулярные полисахариды растений. По химической природе пектиновые вещества представляют собой неразветвленные полимеры галактуроновой кислоты, в которой часть карбоксильных групп этерифицирована метиловым спиртом. В состав пектиновых веществ входят также нейтральные моносахариды — галактоза, рамноза, арабиноза, ксилоза. Пектиновые вещества, входящие в состав матрикса клеточной оболочки, способны сильно набухать в воде и образовывать гель. Некоторые из пектиновых веществ могут растворяться в воде, кроме того, они легко разрушаются под действием кислот и щелочей.
Полисахариды матрикса не просто заполняют промежутки между мицеллами целлюлозы. Они располагаются в оболочке клетки довольно упорядоченно и образуют многочисленные ковалентные или водородные связи как друг с другом, так и с микрофибриллами.