МОРФОЛОГИЯ РАСТЕНИЙ - Т. А. Сауткина - 2012
ГЛАВА 2. КРАТКАЯ ИСТОРИЯ ИЗУЧЕНИЯ КЛЕТКИ. ОСОБЕННОСТИ СТРОЕНИЯ КЛЕТКИ ВЫСШИХ РАСТЕНИЙ
2.3. Образование клеточной оболочки
Образование клеточной оболочки связано с делением клетки (цитокинезом). В конце телофазы митоза в экваториальной зоне делящейся клетки между нитями ахроматинового веретена (веретена деления, митотического веретена) образуется фрагмопласт (рис. 4). Впервые эту структуру обнаружил и описал немецкий ботаник Л. Еррера в 1888 г. и указал, что она имеет бочонковидную форму и состоит из фибрилл. В настоящее время установлено, что фрагмопласт представляет собой систему микротрубочек. Микротрубочки — это лишенные мембраны органеллы, представляющие собой цилиндрические образования диаметром около 24 нм, длина которых может достигать нескольких микрометров. В центре микротрубочка имеет полость, а ее стенка образована 13 рядами димерных комплексов глобулярных белков — тубулинов. Микротрубочки располагаются преимущественно в периферических слоях цитоплазмы. При делении клетки веретено деления и фраг- мопласт оказываются тесно связанными друг с другом благодаря наличию в них общих микротрубочек. Фрагмопласт обнаружен у всех высших растений и у ряда зеленых водорослей (например, Coleochaetae).
Разделение материнской клетки на две дочерние начинается с формирования клеточной пластинки (рис. 4), в образовании которой принимают участие диктиосомы комплекса Гольджи (аппарат Гольджи). Диктиосомы — это одномембранные органеллы, состоящие из плоских округлых цистерн. В состав диктиосом растений входит от двух до семи (реже больше) цистерн, расположенных одна над другой в виде стопки. Между отдельными цистернами стопки имеются небольшие промежутки, заполненные фибриллярными и трубчатыми элементами, функция которых неизвестна. Диктиосомы выполняют секреторную функцию. В них происходит энергичное образование пузырьков (пузырьки Гольджи, тельца Гольджи), в которых содержатся секретируемые вещества — полисахариды или полисахаридно-белковые комплексы, обладающие высокой вязкостью. Вязкость содержимого пузырьков Гольджи объясняется наличием большого количества пектиновых веществ, за что их часто называют пектиновыми пузырьками.
Рис. 4. Фрагмопласт (1) и формирование клеточной пластинки (2) при делении материнской клетки
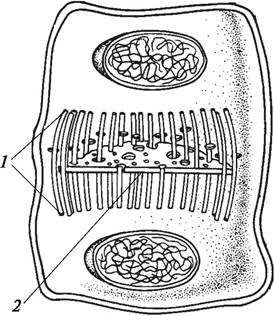
Пектиновые пузырьки мигрируют в область фрагмопласта, выстраиваются в горизонтальном направлении в его центральной части, сливаются и начинают формировать клеточную пластинку. На первых стадиях развития клеточная пластинка закладывается в виде диска в центральной части фрагмопласта. Пектиновые пузырьки неплотно прилегают друг к другу. При слиянии пузырьков в клеточной пластинке остаются небольшие промежутки — плазмодесменные каналы, выстланные плазмалеммой. В них проходят плазмодесмы — мельчайшие цитоплазматические тяжи, соединяющие протопласты дочерних клеток в единую структуру — симпласт. В центре каждой плазмодесмы находится десмотрубочка, представляющая собой не трубчатое образование, а компактный тяж из структурных белков, который с обеих сторон граничащих клеток контактирует с цистернами эндоплазматического ретикулума. Постепенно, по мере того как в центральной части фрагмопласта микротрубочки исчезают, на его периферических концах появляются новые, и фрагмопласт расширяется по направлению к стенкам делящейся материнской клетки. Вместе с ним расширяется и клеточная пластинка, формирование которой идет от центра к периферии. Когда клеточная пластинка достигнет оболочки делящейся клетки, процесс деления фактически завершается. Образовавшиеся дочерние клетки должны достичь определенных размеров (вырасти) и сформировать собственные оболочки.
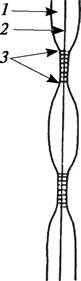
В период роста клетки формируется первичная клеточная оболочка. В образовании клеточной оболочки принимают участие комплекс Гольджи и плазмалемма клетки. На первых этапах развития оболочка дочерних клеток состоит из остатков оболочки материнской клетки и участков вновь образующейся первичной клеточной оболочки. Первичная клеточная оболочка начинает формироваться между клеточной пластинкой и плазмалеммой. Как полагают, гемицеллюлозы и пектиновые вещества доставляются к месту синтеза оболочки пузырьками Гольджи, а образование целлюлозы связано с деятельностью плазмалеммы. Первичная клеточная оболочка довольно пластичная. В ее состав входит около 15 % целлюлозы, остальная часть приходится на долю матричных компонентов. Встраивание молекул целлюлозы в клеточную оболочку может происходить двумя способами: путем интуссусцепции и путем аппозиции. При интуссусцепции новые молекулы целлюлозы внедряются между уже встроившимися в оболочку молекулами. При аппозиции новые молекулы целлюлозы накладываются изнутри на уже отложенные элементы клеточной оболочки. В первичной клеточной оболочке, как и в клеточной пластинке, есть участки, через которые проходят плазмодесмы. Участки первичной клеточной оболочки, пронизанные плазмодесмами и более тонкие, чем остальные, называются первичными норовыми полями (рис. 5).
Рис. 5. Схема строения первичной клеточной оболочки (клеточной стенки): 1—первичная оболочка; 2—срединная пластинка; 3—первичное поровое поле с плазмодесмами
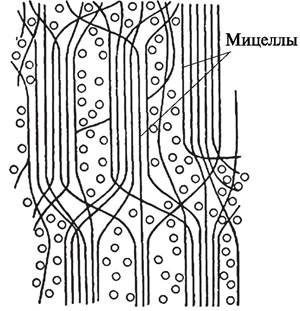
Так как в первичной клеточной оболочке преобладают гемицеллюлозы и пектиновые вещества, содержится много воды, а молекулы целлюлозы расположены неупорядоченно (рис. 6), оболочка способна растягиваться, а клетка может расти как в длину, так и в ширину.
Рис. 6. Расположение мицелл целлюлозы в первичной клеточной оболочке
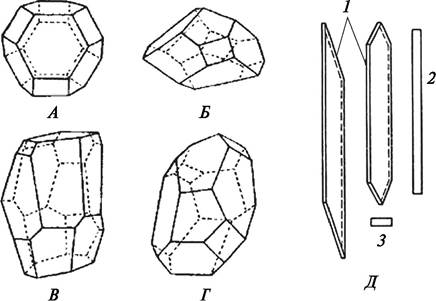
Характер роста определяет будущие морфологические особенности формирующейся клетки. Если клетка более или менее равномерно разрастается в длину и ширину, она становится изодиаметричной (с равными продольным и поперечным диаметром) или таблитчатой (один из диаметров не более чем в два раза превышает другой). Такие клетки называются паренхимными. Если длина значительно превышает ширину, клетки называются прозенхимными (рис. 7).
Рис. 7. Схематическое изображение формы паренхимных (А—Г) и прозенхимных Д клеток: А—геометрически правильный 14-гранник с восемью шестиугольными и шестью четырехугольными гранями; Б—клетка из сердцевины айланта (Аilanthus sp.); В—Г — клетки сердцевины посконника (Eupatorium sp.); Д—клетки камбия: 1—продольный тангентальный разрез; 2—продольный радиальный разрез; 3—поперечный разрез
Первичные клеточные оболочки характерны для клеток меристемы, молодых растущих клеток, а также для клеток некоторых постоянных тканей (паренхимы, эпидермиса, колленхимы, флоэмы).
У большинства клеток, достигших своих предельных размеров и прекративших рост, изнутри клетки над первичной клеточной оболочкой путем аппозиции начинает формироваться вторичная клеточная оболочка (рис. 8). В состав вторичной клеточной оболочки входят те же компоненты, что и в состав первичной, но соотношение полисахаридов в ней иное. Большая часть (до 60 %) приходится на долю целлюлозы, а во вторичных оболочках лубяных волокон льна содержание целлюлозы может достигать 95 %. Кроме того, увеличивается степень полимеризации ее молекул, упорядочивается расположение микрофибрилл в матриксе. Объем матрикса и количество воды значительно уменьшается, но связи между компонентами вторичной клеточной оболочки усиливаются.
Рис. 8. Схема строения вторичной клеточной оболочки: А—общий план строения: 1—первичная клеточная оболочка; 2—срединная пластинка; 3—апертура простой поры; 4—замыкающая пленка поры; 5—вторичная оболочка; 6—входное отверстие поры; Б—направление мицелл целлюлозы в различных слоях вторичной клеточной оболочки
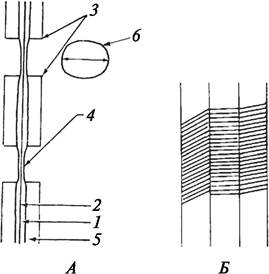
Для вторичной оболочки характерна слоистость. Она возникает в процессе формирования оболочки и связана с расположением в ней микрофибрилл целлюлозы. Наиболее часто вторичная оболочка состоит из трех слоев, которые различаются ориентацией микрофибрилл. В каждом слое микрофибриллы располагаются строго параллельно друг другу, но угол наклона микрофибрилл в каждом слое по отношению к предыдущему меняется.
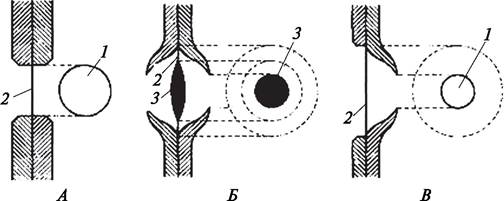
Вторичная клеточная оболочка не сплошная. В тех местах, где в первичной оболочке были сформированы первичные поровые поля, вторичная оболочка прерывается. Перерыв во вторичной клеточной оболочке, расположенный над первичным поровым полем, называется порой (рис. 9). Поры в соседних клетках образуются одна против другой, т. е. возникает пара пор. Эти поры соединяются поровыми каналами и идут от полости клетки до первичной клеточной оболочки, которая становится замыкающей пленкой поры. Поры бывают простыми и окаймленными. Простые поры образуются в том случае, если вторичная клеточная оболочка резко обрывается над первичными поровыми полями. Поровые каналы простых пор по всей длине имеют одинаковый диаметр. При формировании окаймленной поры участки вторичной клеточной оболочки со стороны полости клетки нависают над входным (внутренним) отверстием поры, поэтому входное отверстие гораздо уже, чем наружное, упирающееся в первичную клеточную оболочку (замыкающую пленку пор), и поровый канал имеет форму воронки. Простые поры характерны для клеток паренхимы, лубяных волокон. Окаймленные поры образуются в водопроводящих элементах — трахеидах. У хвойных растений в центре замыкающей пленки окаймленных пор имеется утолщенный участок, образованный срединной пластинкой и ограниченный с двух сторон первичной клеточной оболочкой двух прилегающих друг к другу клеток. Это утолщение называется торусом. Так как первичная клеточная стенка между торусом и стенкой порового канала очень тонкая, торус подвижен. При движении воды по трахеидам торус может закрывать часть пор, что ускоряет передвижение воды по водопроводящим элементам.
Рис. 9. Схема строения пор: А—простая; Б—окаймленная; В—полуокаймленная: 1—входное отверстие поры; 2—выходное отверстие поры; 3—торус
2.4. Вторичные изменения вторичной клеточной оболочки
В процессе жизнедеятельности растения вторичная клеточная оболочка может претерпевать вторичные изменения: одревеснение, опробковение, кутинизацию, минерализацию, ослизнение.
Одревеснение, или лигнификация (от лат. lignum — древесина), — процесс, связанный с отложением в матриксе вторичной клеточной оболочки лигнина — сложного полимерного соединения фенольной природы, химическое строение которого окончательно не установлено. Лигнин — один из важнейших компонентов вторичных клеточных оболочек — образуется протопластом и откладывается только в оболочках клеток высших растений (за исключением мохообразных). Он откладывается в матриксе оболочки некоторых специализированных клеток (склеренхимы, трахеид, члеников сосудов) между микрофибриллами, значительно повышает ее прочность, но снижает эластичность. В то же время лигнин делает оболочку не способной пропускать воду с растворенными в ней минеральными веществами, нарушает газообмен и в конечном счете способствует отмиранию живого содержимого клетки. Одревесневшие клеточные оболочки обладают ультраструктурой, которую можно сравнить со структурой железобетона.
Из жироподобных веществ во вторичных оболочках клеток некоторых тканей (феллемы, эндодермы) откладывается суберин (от лат. suber — пробка). Он вызывает опробковениеклеточной оболочки (явление суберинизации). Суберин не растворяется в органических и неорганических растворителях, в кислотах, но разрушается в концентрированных щелочах. Суберин может откладываться на внутренней поверхности клеточной оболочки или в виде отдельных пластинок, или сплошным слоем. В последнем случае протопласт оказывается в полной изоляции от внешней и внутренней среды и отмирает.
По химической природе к суберину близки гидрофобные полимеры кутин и воск. Кутин откладывается на наружной поверхности клеточной оболочки, образуя тонкую пленку — кутикулу. Кутикула выполняет защитную функцию и предохраняет надземные органы растений от излишнего испарения воды.
Аналогичные функции выполняет и восковой налет, образующийся на поверхности различных органов растения. В отличие от кутина и суберина воск растворяется в органических растворителях и легко плавится. Воск откладывается в виде аморфного или кристаллического слоя и выполняет защитную функцию. Кроме того, воск может входить в состав кутикулы, что усиливает ее барьерные свойства.
Минерализация клеточных оболочек обусловлена отложением в них кремнезема или солей кальция (оксалата, карбоната). Кремнезем в аморфном состоянии откладывается в оболочках эпидермальных клеток некоторых растений (злаков, хвощей, волосков крапивы). Оксалат и карбонат кальция могут откладываться в клеточных оболочках или образуют в клетках цистолиты и различные кристаллы.
Ослизнению оболочек способствует наличие в них большого количества гемицеллюлоз, и особенно пектиновых веществ. Ослизнение сопровождается связыванием большого количества воды, что приводит к образованию на поверхности слизей, представляющих собой смесь полисахаридов и их производных. Ослизнение носит приспособительный характер. В одних случаях оно помогает растению пережить засушливый период и повышает жизнеспособность растений пустыни, в других — способствует размножению, так как семена с ослизнившейся оболочкой надежно прикрепляются к субстрату и быстро прорастают.
Таким образом, оболочка растительной клетки высших растений представляет собой сложную многофункциональную структуру, которая изменяется в зависимости от возраста, функциональных особенностей клетки и видовой специфики растения.
2.5. Пластиды как специализированные органоиды растительной клетки
Для растительной клетки обязательными органоидами являются пластиды (от греч. plastos — вылепленный, оформленный). Пластиды высших растений — двумембранные органеллы, имеющие в зависимости от типа различную форму. У высших растений различают три типа пластид — хлоропласты (от греч. Moms — зеленый и plastos), хромопласты (от греч. chroma — краска, цвет и plastos) и лейкопласты (от греч. leicos — белый и plastos) (рис. 10). Названия пластид по окраске дал немецкий ботаник А. Шимпер (1856—1901): хлоропласты — зеленые, хромопласты — оранжево-красные и лейкопласты — бесцветные. Окраска пластид зависит от пигментов, которые имеются в пластиде. В хлоропластах высших растений содержатся зеленые пигменты — хлорофиллы а и b, а также каротиноиды — пигменты каротин и ксантофил (оранжевый и желтый). В хромопластах накапливаются только каротиноиды. Лейкопласты пигментов не содержат.
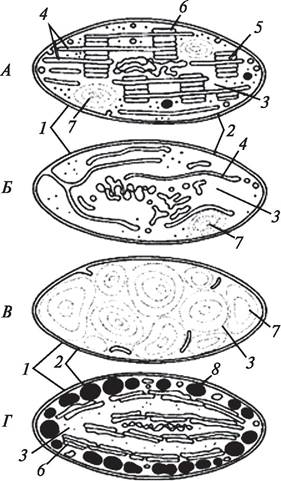
Рис. 10. Субмикроскопическое строение пластид: А—хлоропласта; Б—лейкопласта; В—амилопласта; Г—хромопласта: 1—внешняя мембрана; 2—внутренняя мембрана; 3 —матрикс (строма); 4—ламеллы стромы; 5—грана; 6—тилакоид; 7—крахмальное зерно; 8—липидная капля с пигментами
Субмикроскопическая структура всех пластид сходна. Двумембранная агрануллярная оболочка окружает строму — бесцветную белковую основу пластид, в которую погружена упорядоченная система мембран (тилакоидов) — носителей пигментов. Кроме того, в строме имеются рибосомы, ДНК, ферменты, осуществляющие синтез и гидролиз запасных веществ, и другие компоненты. В пластидах происходит первичный и вторичный синтез органических веществ, а также их накопление.
Число пластид в клетках разных растений (15—50) и их размеры (3—10 мкм) значительно варьируют. Форма пластид также различна, она зависит в первую очередь от субмикроскопической структуры, которая определяется количеством и характером расположения тилакоидов.
Самое сложное субмикроскопическое строение характерно для хлоропластов (рис. 10, А). Хлоропласты высших растений имеют однотипную округло-линзовидную или эллиптическую форму. Эта форма поддерживается благодаря системе строго упорядоченных внутренних мембран — тилакоидов, погруженных в строму. Тилакоиды образуются из внутренней мембраны оболочки хлоропласта и представляют собой уплощенные мешковидные или дисковидные структуры. В строме хлоропласта дисковидные тилакоиды располагаются друг над другом в виде стопки монет и образуют граны. В хлоропластах высших растений формируется от 40 до 60 или более гран. Тилакоиды в гранах связаны друг с другом с помощью фрет, а отдельные граны соединены одиночными тилакоидами — ламеллами. Основная функция хлоропластов — процесс фотосинтеза.
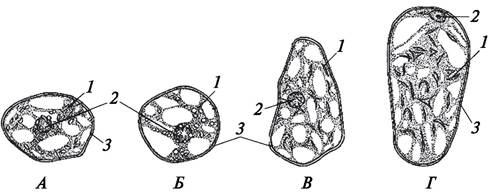
В отличие от хлоропластов в строме хромопластов или вообще нет тилакоидов, или образуются единичные тилакоиды. Вследствие отсутствия сложной субмикроскопической организации хромопласты могут менять свою форму. Форма хромопласта во многом определяется тем, в каком состоянии находятся в них пигменты. Каротиноиды могут растворяться в липидах и накапливаться в пластоглобулах — сферических включениях жирных масел, в которых растворены каротиноиды. В этом случае хромопласты имеют более-менее определенную форму. Если каротиноиды накапливаются в белковых фибриллах или откладываются в виде кристаллов, хромопласты приобретают различную форму, так как их оболочка плотно облегает сформированную структуру (рис. 10, Г; 11). Функция хромопластов — синтез и накопление каротиноидов.
Лейкопласты — самые мелкие из пластид, обычно образуются в подземных органах растений, реже в клетках эпидермиса (рис. 10, Б). Внутренняя мембрана оболочки лейкопласта образует небольшое количество одиночно расположенных тилакоидов. Форма лейкопластов чаще всего сферическая, но может быть и иной. В лейкопластах имеются ДНК, рибосомы, ферменты, осуществляющие синтез и гидролиз запасных веществ, что определяет их функции. В лейкопластах осуществляется накопление или вторичный синтез углеводов, белков и жиров. Лейкопласты, содержащие крахмал, называются амилопластами (рис. 10, В; 12). Амилопласты в большом количестве находятся в корневищах, клубнях, в запасающих тканях семян и т. д. Лейкопласты, в которых накапливаются или вторично синтезируются белки, называются протеопластами. Протеопласты встречаются в эпидермисе листьев представителей семейства Комелиновые (традесканция, зебрина, сеткреазия и др.), в оболочке пыльцевых зерен растений из семейства Ластовневые (ластовень, ваточник). Олеопласты — лейкопласты, накапливающие жиры, характерны для многих представителей класса Однодольные. Они могут находиться в эпидермисе листьев (сем. Орхидные), в листочках околоцветника (птицемлечник из сем. Лилейные), стенках завязи (функия из сем. Лилейные).
Рис. 11. Разнообразие формы хромопластов в клетках мякоти зрелых плодов: А—шиповник собачий (Rosa canina); Б—ландыш майский (Convallaria majalis); В—рябина обыкновенная (Sorbus aucuparia); Г—боярышник кроваво-красный (Crataegus sanguinea): 1—хромопласты; 2—ядро клетки; 3—оболочка клетки
Все пластиды генетически связаны между собой. Они формируются из пропластид, бесцветных образований, ограниченных двумя элементарными мембранами, но не имеющими внутренней мембранной системы. Пропластиды всегда имеются в цитоплазме клетки и при делении ее передаются дочерним клеткам. Обычно в клетке встречается только один тип пластид. Совокупность всех пластид клетки называется пластидомом.
В процессе жизнедеятельности растительного организма пластиды способны менять свое строение и функцию. При разрушении внутренней структуры хлоропласты могут превращаться в хромопласты. Хромопласты могут образовываться также из лейкопластов. Хромопласты практически не могут давать начало другим типам пластид, так как это специализированные органеллы с низким, по сравнению с хлоропластами и лейкопластами, уровнем внутриструктурной организации. Хромопласты часто рассматривают как последнюю стадию организации пластид.
2.6. Вакуоли растительных клеток
Вакуоли (от лат. vacuus — пустой) в растительной клетке представляют собой полости в цитоплазме, окруженные избирательно проницаемой (полупроницаемой) мембраной — тонопластом и заполненные клеточным соком (рис. 12). Основным компонентом клеточного сока является вода, в которой растворены различные вещества, представляющие собой продукты жизнедеятельности клетки. Состав клеточного сока клеток разных растений различен. В плодах, корнеплодах сахарной свеклы, сахарном тростнике содержится много сахаров (сахароза, глюкоза, фруктоза). Вакуоли клеток семян богаты белками, которые концентрируются там в виде коллоидного раствора или в кристаллической форме. В клеточном соке незрелых плодов накапливаются органические кислоты: лимонная, щавелевая, янтарная, уксусная. Клеточный сок некоторых растений содержит дубильные вещества — таннины, безазотистые циклические соединения вяжущего вкуса. Много таннинов в клетках коры дуба, ивы, листьях чая, в незрелых плодах грецких орехов. Для некоторых клеток накопление таннинов — основная функция. Только у высших растений в клеточном соке могут содержаться алкалоиды — разнообразные в химическом отношении азотсодержащие вещества гетероциклического строения, горького вкуса. Известно около 10 тыс. алкалоидов (кофеин, атропин, никотин и др.), чаще всего они находятся в клеточном соке в виде солей. В клеточном соке могут находиться гликозиды — соединения сахаров со спиртами, альдегидами, фенолами и другими веществами. К гликозидам относится и пигмент клеточного сока антоциан.
Рис. 12. Клетки нижнего эпидермиса листа зебрины повислой (Zebrina pendula): 1 —оболочка клетки; 2—клеточное ядро; 3—вакуоли; 4—лейкопласты
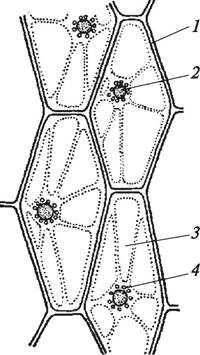
В вакуолях некоторых растений высокая концентрация минеральных солей приводит к образованию кристаллов в виде друз, рафид, стилоидов, кристаллического песка.
Вакуоли растительной клетки выполняют различные функции. Они регулируют водно-солевой обмен, поддерживают тургорное давление в клетке, служат местом отложения запасных веществ и выводят из процесса метаболизма токсичные вещества. Тургорное давление, возникающее в вакуолях, приводит к округлению клеточных стенок, в результате чего возникают межклетники. Система межклетников и клеточных стенок получила название апопласт. Он представляет собой главный путь перемещения по растению водных растворов минеральных веществ.
Процесс образования вакуолей в растительных клетках не вполне выяснен. Одни авторы считают, что они образуются при расширении цистерн эндоплазматического ретикулума, но есть и другие мнения. Вполне вероятно, что вакуоли могут возникать различными способами: путем слияния агранулярных пузырьков, отделившихся от эндоплазматического ретикулума, или из элементов комплекса Гольджи.
Таким образом, характерные особенности строения растительных клеток определяют их свойства и в конечном итоге свойства образуемых ими растительных тканей.