ФІЗІОЛОГІЯ РОСЛИН З ОСНОВАМИ БІОХІМІЇ - Притуляк Р. М. - 2016
Опорний конспект лекцій
ТЕМА № 6. МІНЕРАЛЬНЕ ЖИВЛЕННЯ РОСЛИН
План
1. Розвиток вчення про мінеральне живлення рослин.
2. Методи дослідження мінерального живлення рослин.
3. Механізми поглинання елементів мінерального живлення рослиною.
4. Поняття про макро- і мікроелементи.
5. Доступні для рослин форми сполук фосфору і сірки та їх участь в обміні речовин.
6. Фізіологічна роль К, Са, М>g та ін. елементів.
7. Фізіологічна роль мікроелементів.
8. Азотне живлення рослин. Перетворення нітратів.
9. Діагностика мінерального живлення.
10. Іонний транспорт мінеральних елементів у рослині.
11. Фізіологічні основи застосування добрив.
1. Розвиток вчення про мінеральне живлення рослин.
Мінеральне живлення - це поглинання рослинами необхідних мінеральних елементів і включення їх до обміну речовин. Вищі рослини переважну кількість цих елементів поглинають через кореневу систему, тому це живлення називають також кореневим живленням. Воно є одним із основних факторів регуляції росту, розвитку і продуктивності рослин.
Мінеральне живлення можливе завдяки автотрофності рослин, тобто їх здатності використовувати речовини неорганічної природи для формування своїх тканин і органів. Вчення про мінеральне живлення має давню і досить суперечливу історію.
Майже впритул до відкриття фотосинтезу на рубежі XIX ст. панувала думка про те, що рослини будують своє тіло із "соків землі". Ця думка базувалася на багаторічному досвіді землеробства про важливу роль ґрунту в отриманні врожаю ряду сільськогосподарських культур. Ще у УІ-V ст. до н. є. при вирощуванні жита, пшениці, ячменю, кукурудзи, льону, багатьох овочевих культур для підвищення родючості ґрунту і продуктивності культур використовували гній, попіл, річковий мул.
Арістотель (384-322 рр. до н. є.) вважав, що ґрунт замінює рослині шлунок та інші органи травлення і забезпечує її їжею у вигляді складних речовин.
Німецький агроном А. Теєр наприкінці XVIII - початку XIX ст. це тлумачення Арістотеля розвинув у "гумусову теорію».
Французький природознавець Паліссі у 1563 р. висловив досить вагомі міркування про роль добрив і мінеральних речовин, вважаючи, що гній не мав би ніякого значення, якби у ньому не містилась сіль, яка залишається після його мінералізації.
Англійський хімік Глаубер майже через 100 років (у 1656 р.) своїми дослідами довів позитивний вплив селітри на збільшення врожаю ряду рослин.
Оригінальні думки щодо живлення рослин належать М.В. Ломоносову, який надавав важливого значення не тільки ґрунтовому, але і повітряному живленню.
Французький агрохімік Ж.Б. Буссенго (1837) у своїх дослідах довів, що рослини можна вирощувати і на чистому піску, якщо внести туди мінеральні солі (попіл і селітру).
Німецький хімік Ю. Лібіх розробив теорію мінерального живлення рослин, згідно якої основою родючості ґрунту є мінеральні речовини. При цьому він вважав, що рослини спроможні поглинати азот з повітря у вигляді аміаку. Крім того, Ю. Лібіх не визнавав гумусову теорію і відкидав роль ґрунту як джерела органічних речовин для росту і розвитку рослин. Він вперше сформулював "закон мінімуму», згідно якого приріст урожаю лімітується фактором, який знаходиться у мінімумі. Ю. Лібіх стверджував, що для запобігання виснаженню ґрунтів і зберігання їх родючості необхідно дотримуватись "закону повернення" поживних речовин, поглинутих рослиною. У свій час К.А. Тімірязєв відзначав, що вчення про необхідність повернення речовин є одним із вагомих надбань науки.
Основні положення теорії:
· Корені отримують необхідні для живлення рослин елементи з ґрунту у мінеральній формі.
· Правило незамінності елементів мінерального живлення, тому що кожен елемент виконує певну фізіологічну роль.
· Споживання елементів мінерального живлення відбувається нормовано.
Важливе значення для розвитку теорії мінерального живлення мали досліди І. Кнопа і Ю. Сакса, якими була доведена можливість вирощування рослин до повного дозрівання у водних культурах при забезпеченні їх азотом, фосфором, калієм, сіркою, кальцієм, магнієм, залізом.
У теорію і практику мінерального живлення великий внесок зробили наші співвітчизники, академіки Д.М. Прянишніков та П.А. Власюк.
2. Методи дослідження мінерального живлення рослин.
· Лабораторний метод досліджень.
· Вегетаційний метод досліджень.
· Польовий метод досліджень.
· Виробничий метод досліджень.
Лабораторний - використовують різні субстрати для вирощування рослин. В лабораторних умовах були розроблені поживні суміші для вивчення впливу того чи іншого елементу живлення на рослини.
Суміш Кнопа:
Ca(NO3)2 - 1 г/л;
KH2PO4 - 0,25 г/л;
MgSO4 - 0,25 г/л;
KCl - 0,125 г/л;
FeCl3 - сліди.
Також є суміші Гельрігеля та Прянішнікова.
При складанні поживних сумішей існують певні вимоги:
· Всі елементи мінерального живлення повинні бути урівноважені.
· Жоден елемент не може бути замінений іншим.
· Елементи мають бути у певних співвідношеннях.
· рН повинно бути 6,5-7.
Існують фізіологічно кислі, лужні та нейтральні солі. З кислих солей в першу чергу поглинається катіон (НСl), з лужних - аніон (NaNO3). З нейтральних солей поглинається і катіон і аніон (NH4NO3).
Вегетаційний дослід проводять у контрольованих умовах. Субстрат розміщають у посудину, додають елементи живлення, висаджують рослини та спостерігають за їх розвитком.
Польовий дослід проводиться у польових умовах на дослідній ділянці.
Виробничий дослід виконується у виробничих умовах.
3. Механізми поглинання елементів мінерального живлення рослиною.
Надходження поживних речовин до рослини залежить від їх припливу до поверхні коренів. Існує три способи, якими забезпечується контакт поживних речовин з кореневою системою.
Перший спосіб має назву кореневого перехоплення і полягає в тому, що коренева система у процесі постійного росту займає все більші і більші об'єми ґрунту, у якому містяться поживні речовини, перехоплює і поглинає їх.
Другий - це спосіб масового потоку поживних речовин, що рухаються у формі ґрунтового розчину разом з водою до поверхні коренів і поглинаються ними.
Третій спосіб - надходження поживних речовин до коренів за градієнтом концентрації.
При поглинанні мінеральних елементів біля поверхні кореня їх концентрація зменшується. Внаслідок цього виникає градієнт концентрації, завдяки якому нові іони рухаються в бік кореневої системи.
Усі три способи забезпечують кореневу систему поживними елементами і сприяють мобілізації мінеральних речовин ґрунту. Спосіб руху елемента у ґрунтовому розчині залежить від іона, у вигляді якого цей елемент поглинається рослиною. Наприклад, кальцій і магній (Са++, Мg++) надходять до кореневої системи головним чином за рахунок масового потоку і кореневого перехоплення; катіони К+ - за рахунок дифузії і масового потоку; аніони РО43 - тільки шляхом дифузії.
На сьогодні науковці вважають, що існує кілька механізмів надходження поживних речовин до клітини:
1) проста дифузія речовини крізь пори мембрани за градієнтом концентрації;
2) проходження розчиненої речовини крізь пори мембрани з потоком розчинника;
3) дифузія речовин, що розчиняються в ліпідах, крізь ліпідну фазу мембрани;
4) полегшена дифузія, при якій речовини проникають крізь мембрану у вигляді комплексу їх з мембранними переносниками;
5) обмінна дифузія, коли молекули й іони зовнішнього середовища і цитоплазми обмінюються (шляхом утворення комплексу з мембранними переносниками) молекулами та іонами такого ж виду, і тому концентрація їх у клітині не змінюється;
6) активний транспорт, який вимагає витрат енергії для перенесення речовин через мембрану;
7) піноцитоз.
Спочатку речовина підходить до мембрани, потім взаємодіє з її компонентами певним способом і проникає крізь неї. Після проникнення до клітини речовина підключається до метаболічних процесів.
Залежно від характеру витрат енергії на поглинання і транспорт елементів мінерального живлення рослинами розрізняють два види надходження речовин - активний і пасивний. Перший супроводжується витратами метаболічної енергії і проходить проти градієнта концентрації. Другий відбувається без витрат метаболічної енергії. Пасивне поглинання іде за градієнтом електрохімічної активності.
Вважають, що в рослині існують дві єдині системи транспорту. Перша (пасивного поглинання і транспорту) складається з безперервної гідролітичної системи вільного простору (апопластний шлях); друга (активного поглинання і транспорту) включає в себе протопласти клітин, з'єднані між собою плазмодесмами (симпластний шлях).
4. Поняття про макро- і мікроелементи.
Для нормального життєвого циклу рослинного організму необхідними є такі елементи: вуглець, водень, кисень, азот, фосфор, калій, кальцій, магній, сірка, залізо, марганець, мідь, цинк, молібден, бор, натрій, кремній, кобальт, хлор та
ін. Багато з них є суто мінеральними і поглинаються рослинами, головним чином, з ґрунтового розчину у вигляді іонів. А такі елементи, як вуглець, водень і кисень надходять до рослини переважно у вигляді СО2, Н2О і О2.
Майже вся органічна речовина обов'язково містить чотири елементи - вуглець, водень, кисень і азот, які називаються органогенами. Суха речовина тіла рослин, що залишається після тривалого висушування при температурі 102-105 °С, майже наполовину складається з вуглецю (45%). Кисень становить 42%, водень - 6,5, азот - 1,5% їх маси. На решту елементів, що належать до групи попелових, припадає у середньому близько 5%.
При спалюванні рослин органічні елементи у формі відповідних сполук: Н2О, СО2, Н2S, NН3 у газоподібному стані виділяються в атмосферу, а решта - у формі оксидів становить мінеральну частину рослин і є попелом. Вміст попелу неоднаковий у різних органах і частинах рослин. Найменша його кількість у деревині - близько 1%, у коренях і стеблах - 4-5%, листках - 10-15%, квітках 15-20%, корі - близько 7 і насінинах - 3%. Різною є кількість попелу і у різних рослин. Наприклад, у листках картоплі - 5-13%; буряку - 11-12; ріпи - 8-15%. Склад мінеральних елементів рослин визначається умовами вирощування, віком, фазою розвитку.
Усі хімічні елементи за кількісним вмістом поділено Вернадським на три групи.
Десять з них, які необхідні у найбільшій кількості (від 10 до 0,01% сухої маси рослини), належать до групи макроелементів. Це такі елементи: вуглець, водень, кисень, азот, калій, фосфор, магній, кальцій, сірка.
Елементи, що містяться в рослині у значно меншій кількості (від 0,001 до 0,00001%), складають групу мікроелементів. Серед них - залізо, бор, марганець, цинк, молібден, кобальт і деякі інші.
Третю групу складають ультрамікроелементи. їх вміст у золі рослин не перевищує 106-10-12%. Це - свинець, срібло, літій, ртуть, миш’як та ін.
Такий поділ елементів на три названі групи досить умовний, тому що їх кількість може значно змінюватися. Більш доцільною є класифікація елементів за їх біологічним значенням і фізіологічними функціями.
Вуглець, водень, кисень, азот, фосфор, сірка є основними компонентами органічної речовини, а такі елементи, як калій, кальцій, магній, марганець і хлор виконують регуляційну роль, беручи участь в осмотичній регуляції, балансі електронів і проникності мембран.
5. Доступні для рослин форми сполук фосфору і сірки та їх участь в обміні речовин.
Фосфор. Він належить до елементів-органогенів. У вигляді залишку фосфорної кислоти фосфор входить до складу таких конституційних речовин, як нуклеїнові кислоти, нуклеопротеїди, фосфатиди, до складу окремих запасних речовин, проміжних продуктів обміну, високоенергетичних сполук - рибулозодифосфату, фосфогліцеринової кислоти, фосфогліцеринового альдегіду, аденозинфосфатів (АМФ, АДФ, АТФ та ін.). Фосфор є складовою частиною коферментів, які беруть участь у процесах фотосинтезу і дихання (НАД, ФАД, НАДР, КоА та ін.).
Конституційні сполуки фосфору відіграють особливу роль при побудові ядер (входять до складу ДНК), біологічних мембран. Сполуки фосфору беруть безпосередню участь в енергообміні при таких фізіологічних процесах, як поглинання і транспорт елементів мінерального живлення, перетворення і переміщення запасних органічних сполук.
В усіх тканинах рослин присутні фосфорні ефіри цукрів - сахарофосфати, які відіграють провідну роль у процесах перетворення цукрів при диханні (наприклад, гліколіз), темновій фазі фотосинтезу та інших процесах метаболізму.
Фосфор поглинається коренями з ґрунту у вигляді вищого окислу РО43. Поряд з цією формою до рослин можуть надходити також іони НРО42 і Н3РО4. Фосфорному живленню належить велике значення в початковий період росту і розвитку, оскільки він прискорює ці процеси, сприяє кращому засвоєнню поживних речовин, поглинанню вологи. Мінеральні фосфати, які накопичуються у клітинному соці, беруть участь в утворенні буферних розчинів, які регулюють величину рН клітини. Специфічною властивістю фосфору є здатність прискорювати репродуктивний розвиток рослин, скорочувати вегетаційний період. Метаболізм фосфатів у рослині залежить від багатьох умов. При нестачі води порушуються процеси фотосинтетичного й окислювального фосфорилювання. Азотне голодування призводить до зменшення надходження фосфору в кореневу систему і пересування його до надземних органів. Низька температура зменшує інтенсивність поглинання і метаболізму фосфору, що викликає затримку росту і зниження продуктивності рослин.
Потреба різних рослин у фосфорі неоднакова. Зокрема, злакові поглинають більше фосфору, ніж бобові.
Сірка. Фізіологічна роль сірки, як елемента-органогену, зумовлена тим, що вона у формі сульфогідрильної (R - SH) і дисульфідної (R - S - S - R) груп входить до складу важливих конституційних сполук: окремих білків, ферментів, вітамінів і деяких фізіологічно активних речовин.
Сірка надходить до рослин здебільшого у мінеральній формі - у вигляді сульфатних іонів, але може надходити і у складі окремих амінокислот.
Особливе значення сірки полягає в тому, що вона входить до складу коензиму-А, зумовлюючи його різнобічну високу активність завдяки тіо-ефірному зв'язку. За місцем цього зв'язку можливе приєднання ацетильної групи з утворенням ацетил-КоА (СН3СО~S-КоА), який бере участь в основних реакціях біосинтезу ліпідів, конденсації мономерних молекул до поліпептидних ланцюгів. Він є центром вуглеводного і жирового обміну клітини.
Сірка входить до складу незамінної амінокислоти метіоніну, яка у свою чергу є складовою частиною активного центру багатьох ферментів. Саме метіонінові залишки у складі молекули білка зумовлюють його гідрофобні властивості. Сірка визначає активність і спрямованість багатьох метаболічних процесів. У складі амінокислоти цистеїну вона міститься у формі сульфогідрильної групи, може бути резервом для утворення макроергічних зв'язків. Сульфогідрильні групи слугують активаторами ряду протеолітичних ферментів. В окислювально-відновних процесах важливу роль відіграє реакція перетворення сульфогідрильних груп до дисульфідних і навпаки.
Крім того, сірка входить до ефірних олій родини хрестоцвітних.
У ґрунті сірка перебуває в органічній і неорганічній формах. Переважає органічна форма у вигляді рослинних і тваринних залишків. Неорганічна міститься головним чином у формі сірчанокислих солей кальцію, магнію і натрію. Саме така форма сірки (SO42) доступна рослинам. Менш окислені (SO2) або більш відновлені (H2S) форми її токсичні для рослин.
Поглинута у формі сульфатів сірка відновлюється в рослині й асимілюється при синтезі білків.
Метаболізм сірки в рослині пов'язаний з азотним обміном. В умовах надмірного живлення нітратним азотом посилюється окислення сульфогідрильних груп, що призводить до порушення білкового обміну.
Спеціальних сірчаних добрив промисловість не випускає. Сірка, як домішка, входить до складу фосфорних, частково калійних і калійно-магнезійних добрив. Інколи застосовують елементарну сірку.
6. Фізіологічна роль К, Na, Са, Mg та ін. елементів.
Калій. Засвоюється з ґрунту у вигляді іонів К+. У організмі рослин знаходиться у незв’язаному стані.
Фізіологічна роль:
· Приймає участь у процесі фотосинтезу, зокрема у транспорті електронів по ЕТЛ.
· За його участі виділяється О2.
· Приймає участь у фотофосфорилюванні.
· Сприяє обводненню цитоплазми.
· Приймає участь у розкритті продихів.
· Є коферментом більше ніж 60 ферментів, впливає на вуглеводний і білковий обмін.
· Здатний до реутилізації
· Сприяє підвищенню стійкості до несприятливих умов.
· Найбільше його у молодих рослинах та молодих органах.
Натрій. При нестачі калію натрій має позитивний вплив на ріст і розвиток окремих видів рослин. Значну кількість цього елемента можуть нагромаджувати рослини-галофіти, які ростуть на засолених ґрунтах. Натрій здатний заміщувати калій у поглинальному комплексі ґрунту і таким чином підвищувати доступність останнього для рослин.
Фізіологічна роль натрію інколи аналогічна калію. Наприклад, обидва елементи взаємозамінні при дії на осмотичні властивості клітини, тобто натрій теж сприяє гідратації цитоплазми. Разом з тим, якщо калій активує ферментні системи, сприяючи поєднанню субодиниць ферментних білків, то натрій, навпаки, стимулює їх дисоціацію. Забезпечує осмотичні властивості, надходження води і стимулює розтягування клітин при рості. «Натрієфіли» - буряки, люцерна, капуста, томати. «Натрієфоби» - кукурудза, рис, гречка, соя.
У переважної більшості рослин натрій не входить до числа необхідних елементів. Проте, наявність його у ґрунті, як наголошувалося, має позитивний вплив на рухомість і швидкість поглинання калію і посилення окремих фізіологічних процесів. Підвищена ж кількість засвоюваних форм натрію у ґрунті є шкідливою, оскільки порушує необхідний баланс катіонів рослини.
Кальцій. Кальцій належить до важливих елементів мінерального живлення. Однією з основних його властивостей є здатність впливати на такі фізико-хімічні показники цитоплазми, як густина і проникність. Кальцій є антагоністом одновалентних катіонів, особливо водню. Крім регуляторної ролі, він має і структурну, будучи основою серединних пластинок, що склеюють оболонки сусідніх клітин.
Цей елемент впливає на стан біоколоїдів, ступінь їх гідратації. Він накопичується у старих тканинах і органах рослин у формі оксалатів, пов'язуючи щавлеву кислоту, яка у вільному стані є досить токсичною для цитоплазми. Тому у даному випадку кальцій сприяє знешкодженню щавлевої кислоти. Одночасно цей факт свідчить про неможливість транспортування кальцію від старих органів до молодих, тому що всі солі, утворені кальцієм з органічними кислотами, нерозчинні у воді. Транспорт вільних іонів Са2+ здійснюється через провідну систему ксилеми. Вміст кальцію вищий у наземних органах рослини, ніж у підземних. Цей елемент входить до складу комплексів клітинних органел - ядер, хлоропластів, мітохондрій, у вигляді неорганічних фосфатів та іонів. При взаємодії з негативно зарядженими іонами фосфоліпідів кальцій стабілізує мембрани і зменшує їх пасивну проникність. Він входить до складу фітину і пектинів.
Майже всі реакції, що активуються калієм, інгібуються кальцієм. Разом з тим, кальцій активує діяльність окремих ферментів, наприклад, ферментів-переносників електронів в електротранспортному ланцюзі при фотосинтезі.
Не здатний до реутилізації.
Магній. Як і кальцій, цей елемент надходить до рослини у формі катіона (Мg2+), але на відміну від кальцію нагромаджується переважно в молодих органах і тканинах рослин. Магній входить до складу молекули хлорофілу, а отже, без його участі не може відбутися процес фотосинтезу. Він є поліфункціональним елементом. Багато його функцій подібні до функцій калію і кальцію. Існує думка, що магній сприяє надходженню і руху по рослині фосфору. Разом з фосфором він входить до складу фітину, а тому бере участь у енергетичному обміні клітини. Входить магній і до складу пектинових речовин. Він відіграє важливу роль у вуглеводному обміні, активує ферменти фосфатази, які беруть участь в синтезі і гідролізі сполук глюкози з фосфорною кислотою, є кофактором багатьох ферментів, зокрема ферментів циклу Кальвіна.
Магній пов'язує велику і малу субодиниці рибосоми і підтримує її функціональну активність.
7. Фізіологічна роль мікроелементів.
Мікроелементи характеризуються високою специфічністю і незамінністю. Завдяки участі у ферментативних реакціях вони впливають на обмін речовин, прискорення розвитку рослин, підвищення їх стійкості до грибкових і бактеріальних хвороб, а також до несприятливих зовнішніх факторів (посухи, низьких або високих температур повітря і грунту).
Мікроелементи входять до складу простетичних груп ферментів і контролюють такі життєво важливі процеси, як фотосинтез, дихання, перетворення речовин та ін. Інакше кажучи, мікроелементи є активаторами активаторів, тобто активаторами ферментів. Незважаючи на близькість функцій мікроелементів, кожний з них характеризується певною специфічністю дії.
Залізо. Має здатність міняти валентність і тому має здатність переносити електрони і є складовою ряду ферментів (оксидази, каталази). У смстеиі цитохромів бере участь у синтезі хлорофілу, активує ферменти хлорорфілсинтетази. При нестачі проявляється хлороз.
Альбінізм - генетичне порушення синтезу хлорофілу.
Етіоляція - при відсутності освітлення не проходить утворення хлорофілу.
Бор. Цей елемент у складі ферментів не знайдено, але участь його в багатьох ферментативних процесах доведена. Він позитивно впливає на процес поділу клітин, вуглеводний і білковий обмін. Найбільша кількість бору міститься в паренхімній тканині листкової пластинки молодих рослин. Він сприяє нуклеїновому обміну. Бор впливає на диференціацію і поділ тканини на елементи ксилеми і флоеми, тому особливо необхідний на початковому етапі онтогенезу рослин. Він необхідний також для утворення бульбочок на коренях рослин родини бобових.
Бор не може пересуватись від старих органів рослини до молодих ростучих тканин. Тому при борному голодуванні хворіють і відмирають у першу чергу верхівкові частини стебел рослин.
Мідь. Мідь має безпосереднє відношення до фотосинтезу і дихання. Значна кількість цього мікроелемента концентрується у хлоропластах, входячи до пластохінону. Пластохінон відіграє важливу роль у перенесенні електронів при фотосинтезі. Крім того, мідь входить до складу ряду оксидаз - поліфенолоксидази, цитохромоксидази. Вона захищає від розпаду хлорофіл, прискорює біосинтез триптофану.
Згідно концепції Б.А. Рубіна, входячи до складу широкого спектра ферментних систем дихання, мідь забезпечує рослинам стійкість до несприятливих факторів середовища, зокрема до температурних і фітопатогенних, підвищує солестійкість.
Цинк. Цинк входить до складу ряду ферментів, бере активну участь в окислювально-відновних процесах, посилює дихання, стабілізує його при високих температурах, бере також участь у вуглеводному і білковому обмінах і позитивно впливає на утворення ауксинів і хлорофілу. При достатньому забезпеченні цинк може пов'язуватися із сірковмісними та іншими амінокислотами - цистеїном, гістидином, триптофаном, лізином, серином. Цинк регулює вміст ДНК і РІЖ у клітині, підвищує активність фотосинтезу, активує надходження до рослин азоту, калію, кремнію, марганцю.
Молібден відіграє важливу фізіологічну роль у синтезі, перетворенні і пересуванні вуглеводів по рослині. Маючи важливий вплив на розвиток азотобактера у ґрунті, він відіграє помітну роль у біологічній фіксації атмосферного азоту і в азотному обміні рослинного організму. Молібден входить до складу нітратредуктази і бере участь у процесі відновлення нітратів у кореневій системі та в листках рослин. При нестачі молібдену зменшується вміст загального і білкового азоту, уповільнюється відновлення нітратів, що надійшли до рослин, внаслідок чого зменшується синтез амінокислот. Молібден є також незамінним металокомпонентом ферментів, які беруть участь у тканинному диханні.
Рухомість ґрунтового молібдену і його доступність рослинам залежить не тільки від наявності валових форм, але також від ступеня окультуреності ґрунту і застосування добрив. Фізіологічно кислі добрива, особливо при внесенні їх на невапнованих малобуферних піскових ґрунтах, посилюють молібденове голодування. Аналогічний вплив має надлишок у ґрунті важких металів - свинцю, міді, цинку та ін.
Марганець. Існує думка, що забезпечення рослин марганцем визначає ріст і морфогенез органів рослин. При дефіциті цього елемента спостерігається затримка фази розтягування клітин, особливо у кореневій системі.
Виявлено вплив марганцю на переміщення індолілоцтової кислоти і на проростання насіння. Він активує надходження цієї сполуки з насіння до паростків, сприяє утворенню компонентів "індолілоцтова кислота -дезоксирибонуклеїд", які впливають на морфогенез рослин.
При проростанні насіння марганець сприяє процесам гідролізу і пересуванню фосфорних сполук з насінини до стебла і коренів, прискорюючи біосинтез органічних фосфорних сполук у рослині. Нестача його викликає значний дефіцит індолілоцтової кислоти, порушення транспорту фосфору до наземних органів рослини.
Ступінь забезпечення та інтенсивність поглинання марганцю тісно пов'язані із значенням рН ґрунту. При реакції ґрунтового розчину, близькій до нейтральної або лужної (рН 6,5-7,5), розчинність цього елемента і його засвоєння зменшуються.
Хлор. У рослині хлор міститься в дуже малій кількості. Він впливає на функції хлоропластів (фотосинтетичне фосфорилювання), бере участь у процесах виділення.
Крім розглянутих нами, рослина потребує такі мікроелементи, як кобальт, ванадій, хром та інші.
8. Азотне живлення рослин. Перетворення нітратів.
Азот. Був відкритий Резерфордом у 1732 році. Фізіологічне значення азоту визначається насамперед тим, що він є складовою частиною амінокислот, білків, нуклеїнових кислот і багатьох життєво важливих органічних сполук, які становлять основу протоплазми.
На Землі основна кількість азоту перебуває у газоподібному стані у вигляді молекулярного азоту 14, і складає близько 80% загального об'єму повітря. Запаси азоту в атмосфері оцінюються величиною приблизно 8 тонн на кожний квадратний метр Землі. Але ця величезна кількість азоту недоступна рослині. Посередниками, що сприяють переходу атмосферного молекулярного азоту в доступну форму, є мікроорганізми - азотофіксатори грунту. За рахунок азотофіксаторів, що вільно живуть у ґрунті, а також симбіотичних азотофіксаторів багаторічних і однорічних бобових рослин на 1 га ґрунту може щорічно нагромаджуватися 130-140 кг азоту.
З інших форм азоту в повітрі може бути незначна кількість аміаку й оксидів азоту, які утворюються при електричних розрядах. Ці форми азоту з атмосферними опадами потрапляють у ґрунт. Загальна кількість їх дуже мала, і тому вони не мають суттєвого значення в азотному живленні рослин.
У ґрунті азот міститься переважно у трьох видах сполук: амонійних солях, нітратних солях, органічних сполуках у вигляді різних рослинних і тваринних решток і продуктів їх розпаду (амінокислоти, пептиди, аміни). Рослини засвоюють переважно нітратну й аміачну форми азоту, запаси яких становлять не більше 1 -2% загальної його кількості у ґрунті. Тому навіть багаті на органічну речовину чорноземи мають не більше 200 кг доступного азоту в орному шарі 1 га площі. Такої кількості азоту достатньо для отримання високого врожаю зерна. У дерново- підзолистих ґрунтах з низьким вмістом органічної речовини легкодоступних форм азоту у 3-4 рази менше.
В основу класифікації покладено співвідношення між запасними вуглеводами і білками (C:N) у насінні рослин. Найбільш успішно аміак засвоюється рослинами з переважаючим вмістом вуглеводів. Прикладом можуть бути злаки, у яких співвідношення C:N у зерні становить приблизно 6:1. Паростки злаків, перебуваючи у темряві, легко засвоюють аміак до майже повного використання запасів вуглеводів насіння. В окремих бобових культур (горох, вика) співвідношення C:N менше, і їх живлення аміаком у темряві можливе лише при усуненні фізіологічної кислотності поживного розчину (додавання до розчину СаСО3). Паростки насіння з іще меншим вмістом вуглеводів (наприклад, люпин) зовсім не можуть асимілювати аміак у темряві.
Із слабокислого середовища (рН близько 5) рослини краще засвоюють нітратну форму азоту, а з нейтрального (рН близько 7) інтенсивніше поглинається аміачна форма азоту. Як уже наголошувалося, для використання аміачного азоту рослина повинна мати достатню кількість вуглеводів, які забезпечували б включення його у процеси метаболізму клітин. У протилежному випадку накопичення аміаку в клітинах рослин є згубним.
Вміст азоту в ґрунті у доступній для рослин формі визначається не тільки інтенсивністю мікробіологічних процесів мінералізації органічного азоту й азотофіксації, але й процесів поглинання його кореневою системою рослин, вимивання водою з орного шару. Крім цього, мають місце процеси його денітрифікації аеробними мікроорганізмами, які відновлюють нітратний азот до молекулярного. Останній вивітрюється з ґрунту в атмосферу. Цей процес особливо активний на перезволожених ґрунтах зі слабкою аерацією (наприклад, на рисових плантаціях).
Поглинутий рослиною азот включається до органічних сполук тільки у формі аміаку. Згідно сучасних даних, процес відновлення поглинутих нітратів каталізують флавопротеїдні ферменти. Він супроводжується зміною валентності атома азоту:
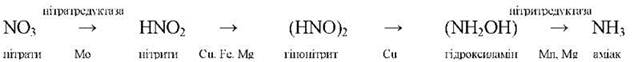
Нітратредуктаза - це індукований клітиною фермент у відповідь на надходження до неї нітрат-іона. Локалізована вона у цитоплазмі. Активність її особливо висока у клітинах молодих органів і меристемних тканинах. Активність нітритредуктази у 5-20 разів вища, ніж нітратредуктази. Тому нітрити не накопичуються, а швидко відновлюються до аміаку. Місцем локалізації нітритредуктази в листках є хлоропласти, а в коренях - пропластиди.
Відновлення нітратів у різних рослин може відбуватись у коренях і наземних органах. У зв'язку з цим виділяють три основні групи рослин.
Перша група - це рослини, у яких нітратна форма азоту практично повністю відновлюється в коренях і надходить до надземних органів у вигляді органічних сполук. До цієї групи належить більшість деревних рослин.
Друга група - це рослини з редукцією нітратів у листках. Це ряд представників родини лободових (буряк, лобода), бавовник, нетреба та ін.
Третя група рослин найбільш чисельна. Рослини цієї групи характеризуються здатністю відновлювати нітрати і в листках, і у коренях. Як правило, активність нітратредуктази в листках вища, ніж у коренях.
Процес відновлення нітратів у листках тісно пов'язаний із процесом фотосинтезу, який є джерелом АТР, енергія якої необхідна для синтезу ферментів (нітратредуктази) і транспорту нітратів, і джерелом органічних кислот, що служать субстратом для зв'язування аміаку.
Аміак, поглинутий кореневою системою з ґрунту й утворений шляхом відновлення нітратів у рослині, вступає у реакції з кетокислотами й утворює амінокислоти та аміди. Провідна роль у процесах зв'язування аміаку в рослині належить реакціям біосинтезу глютамінової кислоти і глютаміну.
Азотне живлення бобових рослин має специфічні особливості, зумовлені їх здатністю вступати у симбіоз із мікроорганізмами. При цьому на коренях рослин утворюються нарости - бульбочки, у яких знаходяться бактерії, що можуть засвоювати атмосферний молекулярний азот. Вони накопичують таку кількість азоту, що можуть забезпечувати не тільки власні потреби в цьому елементі, але і потребу тих рослин, на коренях яких вони розвиваються. У свою чергу рослини забезпечують потребу бактерій у "їжі" у вигляді органічних сполук - продуктів фотосинтетичної діяльності (головним чином дисахаридів і моносахаридів).
Азотофіксуюча активність бактерій залежить від наявності у ґрунті азоту, в першу чергу - нітратів. При високому забезпеченні ґрунту азотом фізіологічна активність бульбочкових бактерій зменшується. Такий же вплив має і підвищена кислотність ґрунту.
Завдяки азотофіксуючій діяльності бульбочкових бактерій на 1га у симбіозі з бобовими рослинами може засвоюватися понад 100 кг атмосферного азоту. Тому необхідно створювати найбільш сприятливі умови для розвитку і фізіологічної активності мікроорганізмів. З цією метою застосовують бактеріальні препарати, наприклад, нітрагін.
9. Діагностика мінерального живлення.
Діагностика потреби мінерального живлення рослин є візуальна і хімічна.
Візуальна - проводиться за зовнішніми симптомами нестачі того чи іншого елементу.
Хімічна - більш досконала. В її основі знаходяться кольорові реакції взаємодії соку рослин з відповідними реактивами.
10. Іонний транспорт мінеральних елементів у рослині.
Пересування речовин у рослині забезпечується двома основними процесами:
1) транспіраційним, або висхідним, що несе воду і розчинені у ній поживні речовини від кореня до пагонів;
2) течією асимілятів, або низхідною течією, що здійснює транспорт утворених при фотосинтезі речовин від листків до частин рослин, які знаходяться нижче (стебло, корінь та ін.) і вище (верхівка стебла, пагонів, плоди та ін.) листка. Висхідна течія здійснюється по ксилемі, що включає специфічні провідні елементи тканини - судини і трахеїди. Низхідна течія здійснюється по флоемі, яка складається із ситовидних трубок і клітин-супутниць.
Транспорт мінеральних елементів по рослині здійснюється такими шляхами:
· Радіальний.
· Дальній.
· Ближній.
Радіальний. Перед тим, як потрапити до провідної системи, поживні речовини проходять через радіальну зону кореня.
Першим бар'єром при надходженні іонів у рослину є клітинна стінка, що складається з двох шарів первинної і вторинної оболонки. Суберин має гідрофобні властивості, тому оболонка стає непроникною для води. Такі суберинізовані оболонки є тільки у клітин ендодерми. Вони мають назву поясків Каспарі.
Целюлозні оболонки всіх клітин і міжклітинний простір рослини утворюють апопластну систему, або апопласт.
Апопластний рух іонів по оболонках екзодерми і мезодерми затримується поясками Каспарі. Лише певна частина іонів надходить до судин ксилеми апопластним шляхом завдяки наявності в ендодермі пропускних клітин. Основна ж кількість поживних елементів у рослинних тканинах переміщується симпластним шляхом.
Симпласт складається з протопластів усіх клітин, що сполучені одна з одною за допомогою плазмодесм.
Іони, що надійшли через плазмалему протопласта, можуть мігрувати по плазмодесмах без витрат енергії. Плазмодесми пронизують клітини кори, пояски Каспарі, ендодерму і паренхіму центрального циліндра у радіальному і поздовжньому напрямах. Число плазмодесм варіює в широких межах, залежно від віку тканин. Вони тісно пов'язані з ендоплазматичним ретикулумом, який сприяє симпластному транспорту.
Дальній. Відбувається по ксилемі, яка представлена судинами, трахеями та трахеїдами. Це висхідна течія, від центрального циліндра до листків.
Ближній. Відбувається у листку, від центральної жилки листка до клітин мезофілу.
11. Фізіологічні основи застосування добрив.
Використання мінеральних добрив — основний засіб підвищення врожайності сільськогосподарських культур. За рахунок азоту, фосфору, калію та інших дефіцитних у землеробстві елементів, які містяться у відповідних типах мінеральних добрив, культурні рослини краще використовують енергію Сонця і ґрунтово-кліматичні фактори, що дає змогу одержувати додаткові врожаї. Нестачу цих елементів неможливо замінити ніякими іншими агротехнічними заходами.
Ефективність добрив залежить від потреб рослин у поживних елементах і здатності ґрунту задовольняти ці потреби. Таку взаємозалежність образно виразив Д. М. Прянишников у вигляді трикутника, де у вершинах кутів розташовуються рослина — ґрунт — добрива, причому рослина розміщена у верхньому куті. Цим підкреслюється, що рослина і її врожайність — основа всіх проблем агрономічних і біологічних наук у практиці сільського господарства. Існують три загальноприйняті правила діагностики живлення.
- Впродовж вегетаційного періоду контролюють ступінь забезпечення посіву майбутнього врожаю основними елементами живлення. Основою діагностики живлення є хімічний склад рослин протягом вегетації. Одночасно з хімічним аналізом рослин слід враховувати етапи їхнього росту і розвитку.
- Визначають кілька поживних елементів, причому не менше трьох основних — азот, фосфор, калій.
- Зіставлення даних хімічної діагностики з урахуванням особливостей росту рослин та кліматичними умовами й агротехнікою.
Також при складанні системи удобрення повинні враховуватися:
· Фізіологічна роль мікроелементів.
· Потреба в них на окремих етапах органогенезу.
· Винос елементів живлення запланованим урожаєм.