ФІЗІОЛОГІЯ РОСЛИН З ОСНОВАМИ БІОХІМІЇ - Притуляк Р. М. - 2016
Опорний конспект лекцій
ТЕМА № 5. ДИХАННЯ РОСЛИН
План
1. Загальні уявлення про дихання рослин і його фізіологічна роль.
2. Теорії дихання А. М. Баха, В. І. Палладіна.
3. Ферментативні системи дихання.
4. Взаємозв'язок процесів бродіння і дихання.
5. Гліколіз як підготовчий етап аеробного дихання, його енергетика.
6. Цикл трикарбонових кислот. Енергетика циклу Кребса.
7. Цикл гліоксалевої кислоти.
8. Пентозофосфатний цикл.
9. Окислювальне фосфорилювання.
10. Дихальний коефіцієнт і його залежність від природи окислювального субстрату.
11. Дихальний газообмін рослин як фактор продукційного процесу. Взаємозв'язок процесів фотосинтезу і дихання
1. Загальні уявлення про дихання рослин і його фізіологічна роль.
В природі існують два основні процеси, в ході яких енергія сонячного світла, що була акумульована в органічній речовині, вивільняється, - це дихання та бродіння.
Дихання - це аеробний окислювальний розпад органічних сполук на прості, неорганічні, що супроводжується виділенням енергії.
Бродіння - анаеробний процес розпаду органічних сполук на більш прості, що також супроводжується виділенням енергії.
Дихання можна визначити як розпад метаболітів через гліколітичний і (або) окислювальний пентозофосфатний шлях з наступним окисленням продуктів у циклі трикарбонових кислот та використанням відновлених піридиннуклеотидів для синтезу АТФ у процесі окислювального фосфорилювання. Дихання - це контрольоване розщеплення або окислення молекул органічної речовини.
У більшості випадків основним джерелом енергії і відновлюючої сили для метаболічної активності рослин є вуглеводи. Проміжні сполуки, які утворюються під час окислення, використовуються як вихідний матеріал для ряду синтетичних реакцій. Відщеплені у процесі окислення органічної речовини електрони використовуються для відновлення НАД, а потім надходять до електротранспортного ланцюга, поступово передаються по системі цитохромів а, в, і с, де на кожному новому етапі переходять на все більш низький енергетичний рівень і нарешті приєднуються до кисню. При цьому утворюється вода.
Субстратами дихання у вищих рослин можуть бути білки, амінокислоти, ліпіди.
Виділена енергія використовується рослиною для утворення складних органічних речовин у процесах метаболізму. Саме дихання є джерелом енергії для росту рослин, різних синтетичних реакцій, поглинання елементів мінерального живлення, транспорту асимілятів. Значення дихання полягає у тому, що цей складний окислювально-відновний процес є джерелом енергії і лабільних сполук, необхідних для процесів життєдіяльності рослинного організму. Тобто, завдяки диханню відбувається перетворення синтезованих у процесі фотосинтезу органічних сполук і використання їх для побудови тіла рослин.
2. Теорії дихання А. М. Баха, В. І. Палладіна.
У 1845 р. швейцарський хімік Х.Ф. Шейнбайн запропонував теорію окислювальних процесів, згідно якої рослинні клітини вміщують сполуки, що здатні легко окислюватися у присутності О, і тим самим активувати молекулярний кисень. Але він помилково вважав, що активація кисню відбувається шляхом утворення озону
Продовжені О.М. Бахом, ці дослідження дали можливість створити у 1897 р. теорію біологічного окислення, яка отримала назву перекисної теорії Баха. її суть полягає в тому, що молекулярний кисень повітря (О = О) після взаємодії з ненасиченими органічними сполуками активується: (— О — О —). Цей активований кисень сполучається з окислювальною речовиною (А), утворюючи перекис (АО2). Такий перекис взаємодіє з другою сполукою (В) і окислює її спочатку одним атомом кисню. Реакція може повторюватися, і тоді другий атом кисню окислить другу молекулу речовини В. Схема цього процесу така:
1. Активування молекулярного кисню:
Вихідна речовина - акцептор кисню (А) - повністю відновлюється, а субстрат (В) окислюється.
У теперішній час відомо, що шлях включення кисню в органічну сполуку, згідно з перекисною теорією біологічного окислення Баха, не має відношення до дихання, але ця теорія відіграла значну роль у сучасному розумінні механізмів активації кисню. Визначенню ролі кисню при окисленні субстрату сприяли дослідження німецького біохіміка О.І. Варбурга та англійського І.Д. Кейлі, які у 20-х роках встановили наявність зв'язку між поглинанням кисню і функціонуванням цитохромної системи.
Ще О.М. Бах поряд із перекисною теорією висловив і другу гіпотезу, згідно якої біологічне окислення може здійснюватися шляхом відщеплення від субстрату попередньо активованого водню. У такому випадку роль кисню полягає в регенерації первинного акцептора водню - хінону. Послідовність цих реакцій така:
Ця гіпотеза постулює наявність у клітині специфічних сполук (хінонів і хіноноподібних речовин), які виконують роль переносників водню дихального субстрату. Вона отримала подальший розвиток у теорії хімізму дихання В.І Палладіна (1912), який припустив наявність специфічних посередників дихання і назвав їх дихальними хромогенами. Віддаючи водень, хромоген окислюється і перетворюється у пігмент, який здатний до зворотного відновлення за рахунок водню субстрату. За В.І. Палладіним, дихальний субстрат окислюється з участю води, яка виконує роль донора водню для відновлення дихального пігменту до хромогену. У процесі відновлення пігменту бере участь і водень субстрату, що окислюється. Кисень води йде на окислення вуглецю субстрату. Отже, вуглекислий газ при диханні утворюється анаеробним шляхом. Кисень, який поглинається під час дихання, згідно теорії В. Палладіна, використовується на окислення хромогенів до пігментів. Роботи О.А. Курсанова і Б.Б. Вартапетяна довели, що поглинутий кисень йде на утворення води.
3. Ферментативні системи дихання.
Окиснювальний апарат рослин має свої специфічні особливості. Насамперед, на відміну від тварин, для нього характерні:
· делокалізація дихального апарата (мітохондрії, пероксисома, цитоплазма та ін.);
· поліфункціональність, тобто наявність каталізаторів, що характеризуються багатьма властивостями;
· принцип, коли в організмі міститься не один, а кілька ферментів, які каталізують ті самі або близькі реакції.
Як відомо, біологічне окиснення будь-якої речовини передбачає втрату нею електронів або електронів разом із протонами. В рослинній клітині відбуваються окиснювальні реакції різних типів.
- Реакції, в яких спостерігається безпосередня втрата електрона в результаті зміни його валентності. Такі реакції каталізують і ферменти оксидази, в активних центрах яких є метали:
Fe2+ → Fe3+ + e
- За рахунок вилучення з окиснюваної речовини водневих іонів за допомогою ферментів гідроксилаз. Акцепторами протонів у таких реакціях часто є сам кисень:
АН2 + В → А + ВН2.
- Пряме окиснення молекулярним киснем за допомогою оксигеназ або трансфераз:
АН2 + О2 → АО2.
- Утворення проміжних гідратованих сполук з наступним вилученням двох електронів і протонів:
СН3СОН + Н2О → СН3СН(ОН)2 → СН3СООН
У біологічних системах усі ці шляхи взаємопов'язані, причому пірата електрона в разі окиснення супроводжується, як правило, і тратою водню. Саме тому процес окиснення певної речовини в живих системах поєднується з відновленням іншої.
Наявність альтернативних механізмів окиснення органічних речовин зумовлює здатність рослин зберігати необхідний ефективний рівень дихання за несприятливих умов.
Щоб знати напрям руху електронів, уведено поняття стандартного відновлювального потенціалу (Ео). За нуль потенціалу умовно прийнято вважати відновлювальний потенціал реакції
Н2 → 2Н+ + 2е-
Універсальним окиснювачем є кисень О2, який має максимальній позитивний потенціал (+0,817 В), а функцію донорів електронів виконують різні органічні речовини. Всі електрони транспортуються до О2 через багатокомпонентну окисно-відновну систему, і окиснення однієї речовини (донора електронів і протонів) поєднується з відновленням іншої сполуки (акцептора електронів і протонів). Ферменти, що каталізують такі реакції, називають оксидоредуктазами. Є три групи оксидоредуктаз: анаеробні дегідрогенази, аеробні дегідрогенази і оксидази.
4. Взаємозв’язок процесів бродіння і дихання.
Бродіння - це дисиміляційний процес з утворенням продуктів неповного окислення. Залежно від кінцевого продукту, розрізняють бродіння спиртове, молочнокисле, оцтовокисле, маслянокисле та ін. Процеси бродіння властиві здебільшого нижчим гетеротрофним організмам (дріжджі, інші гриби, бактерії). У тканинах вищих рослин без наявності кисню також може відбуватися спиртове і молочнокисле бродіння. Продукти бродіння містять значну кількість енергії. Тому енергетичний вихід при бродінні значно нижчий, ніж при аеробному диханні:
С6Н12О6 → 2С2Н5ОН + 2СО2 + 117 кДж.
Таке окислення субстрату за рахунок зв'язаного кисню також називають інтрамолекулярним диханням. Накопичення етилового спирту у тканинах може бути причиною отруєння і загибелі рослин. Крім того, при анаеробному диханні (бродінні) рослина не отримує проміжних продуктів, які звичайно утворюються при аеробному окисленні.
Між диханням і бродінням існує достатньо тісний зв'язок, про що свідчить спільність першого анаеробного етапу (фази) перетворення глюкози.
Перший етап бродіння і дихання - це гліколітичне розщеплення глюкози до піровиноградної кислоти (ПВК). Наступний етап - аеробний. Піровиноградна кислота розщеплюється з утворенням кінцевих продуктів СО, і Н2О при диханні. В анаеробних умовах вона зазнає неповного окислення бродіння.
Про єдність цих процесів свідчить і те, що у рослин виявлені ферменти, які каталізують спиртове бродіння. Крім того, серед проміжних продуктів аеробної фази дихання і бродіння є однакові, наприклад: яблучна, лимонна та інші органічні кислоти.
5. Гліколіз як підготовчий етап аеробного дихання, його енергетика.
На початку XIX ст. М. Соссюр зробив важливе спостереження: при вирощуванні в атмосфері без кисню зелені рослини виділяють СО2. Подальше вивчення цього явища дало змогу Л. Пастеру встановити, що в анаеробних умовах рослини не тільки виділяють вуглекислий газ, але й накопичують спирт та інші продукти спиртового бродіння.
Фундаментальну роль у визначенні хімізму перетворення цукрів мало відкриття Л.О. Івановим явища анаеробного розщеплення не інертної молекули глюкози, а її фосфорного ефіру, який має високу реакційну здатність. Джерелом фосфорної кислоти є АТФ. Перенесення залишку фосфорної кислоти на молекулу глюкози каталізує фермент гексокіназа. Подальше активування гексози відбувається шляхом ізомеризації: глюкозо-6-фосфат перетворюється у фруктозо-6-фосфат. Процес ізомеризації проходить за участю фосфогексоізомерази. Наступний етап супроводжується приєднанням ще одного залишку фосфорної кислоти від молекули АТФ - утворюється фруктозо-1,6-дифосфат за участю ферменту фосфогексокінази. Далі молекула фруктозо-1,6-дифосфату під дією ферменту альдолази розщеплюється на дві фосфотріози: фосфодиоксиацетон і 3-фосфогліцериновий альдегід. Це перша стадія гліколітичного розщеплення глюкози (гліколізу) Завдяки наявності в клітинах специфічного ферменту фосфотріозоізомерази відбувається перетворення фосфодіоксиацетону у фосфогліцериновий альдегід.
На наступній стадії гліколізу фосфогліцериновий альдегід через ряд реакцій перетворюється у фосфогліцеринову кислоту. Спочатку до фосфогліцеринового альдегіду приєднується ще один залишок фосфорної кислоти від АТФ. Утворений 1,3-дифосфогліцериновий альдегід за допомогою специфічної дегідрогенази окислюється до 1,3-дифосфогліцеринової кислоти. Енергія, яка вивільняється при цьому, спочатку зосереджується в одному із фосфатних зв'язків дифосфогліцеринової кислоти, а потім переноситься з участю трансфосфорилази на АДФ і таким чином утворюється молекула АТФ і молекула 3- фосфогліцеринової кислоти. Далі фермент фосфогліцеромутаза ізомеризує 3-фосфогліцеринову кислоту до 2-фосфогліцеринової, від якої потім під дією енолази відщеплюється молекула води. Ця реакція супроводжується перерозподілом енергії в молекулі, внаслідок чого утворюється фосфоенолпіровиноградна кислота, молекула якої має макроергічний зв'язок. Цей фосфат за участю піруваткінази передається на АДФ (утворюється АТФ), а енолпіровиноградна кислота внаслідок своєї нестійкості самовільно перетворюється у піровиноградну кислоту - кінцевий продукт гліколізу. Процес відбувається у цитоплазмі.
Гліколіз - це перший з вивчених метаболічних процесів. Термін "метаболізм" походить з грецької мови і означає "зміна" або ж "акт розкидання". Цим словом визначають сукупність хімічних процесів, які відбуваються в живих організмах.
Перетворення до піровиноградної кислоти під час першого і другого субстратного фосфорилювання супроводжуються утворенням чотирьох молекул АТФ Але для активування глюкози на першій стадії були витрачені дві молекули АТФ Тому чистий вихід гліколітичного субстратного фосфорилювання становить дві молекули АТР.
На другій стадії гліколізу відновлюється по одній молекулі НАД-Н2 на кожну з двох молекул фосфотріоз. Окислення ж однієї молекули НАД-Н2 у електротранспортному ланцюзі мітохондрій у присутності кисню сполучене із синтезом трьох молекул АТФ. Отже, у розрахунку на одну молекулу глюкози синтезуються шість молекул АТФ Усього у процесі гліколізу утворюється вісім молекул АТФ. Вільна енергія гідролізу однієї молекули АТФ становить близько 42 кДж/моль (10 ккал). Тому повний енергетичний баланс гліколізу дорівнює 42 · 8 = 336 кДж/моль, або 80 ккал.
6. Цикл трикарбонових кислот. Енергетика циклу Кребса.
Кінцевий продукт гліколітичного розщеплення глюкози - піровиноградна кислота (СН3СОСООН) - в аеробних умовах зазнає подальших перетворень і окислюється до вуглекислого газу і води в дихальному циклі Кребса (цикл ди- і трикарбонових кислот).
Суть цих перетворень полягає в послідовному ступінчастому декарбоксилюванні і дегідруванні піровиноградної кислоти. Це окислення, як довів англійський біохімік Г. Кребс, супроводжується утворенням ди - і трикарбонових кислот і наступним їх окисленням до СО, за рахунок відщеплення водню.
Згідно теорії окислення В. Палладіна, кисень води, яка бере участь у цих перетвореннях, використовується для окислення вуглецю піровиноградної кислоти, а водень разом із воднем ПВК при участі дегідрогеназ спрямовується до активованого оксидазами кисню повітря. Важливим компонентом циклу окислення є щавлевооцтова кислота, яка сприяє повному окисленню піровиноградної.
Безпосереднього окислення зазнає не сама піровиноградна кислота (ПВК), а її похідна - ацетил-коензим А (ацетил-СоА), який утворюється шляхом окислювального декарбоксилювання ПВК при участі коензиму А (СоА) і НАД+: Подальше окислення ацетил-СоА відбувається у циклічному процесі. Ацетил-СоА конденсується з щавлевооцтовою кислотою і водою. При цьому утворюються лимонна кислота і два електрони, які починають свій шлях до кисню, регенерується молекула СоА. Лимонна кислота через послідовні перетворення (цис-аконітова, ізолимонна, щавлевобурштинова, акетоглутарова, сукциніл-СоА, бурштинова, фумарова і яблучна кислоти) дає щавлевооцтову кислоту (регенерація) і завершує цикл. Цей цикл вміщує 13 основних реакцій, у п'яти з яких вивільнюється енергія і фіксується у вигляді пірофосфатних зв'язків аденозинтрифосфату (АТР). Це такі окислювально-відновні реакції: утворення ацетил-СоА, окислення лимонної кислоти та ізолимонної кислоти через цис-аконітову до щавлевобур штинової кислоти; а-кетоглутарової - до сукциніл-СоА, бурштинової - до фумарової, яблучної - до щавлевооцтової.
Усі реакції циклу можна поділити на три етапи:
1. Декарбоксилювання піровиноградної кислоти й активування з утворенням ацетил-коензиму А.
2. Окислення піровиноградної кислоти.
3. Регенерація щавлевооцтової кислоти.
З кожним обертом циклу зникає одна молекула піровиноградної кислоти, від різних компонентів циклу відщеплюються три молекули СО 2 і п'ять пар атомів водню (електронів), включаються три молекули води. Через систему переносників електрони взаємодіють із системою цитохромів або інших оксидаз (наприклад, поліфенолоксидази) і зрештою - з киснем. Енергія електронів використовується на утворення 12 молекул АТФ. Відтворена щавлевооцтова кислота знову вступає у ланцюг вищезазначених процесів.
Рослина постійно містить майже всі органічні кислоти, які входять до циклу ди- і трикарбонових кислот, а також усі ферментні системи, що беруть участь у перетворенні цих сполук. Місцем локалізації ферментів є матрикс мітохондрій. У ньому ж знаходяться і ферменти окислення жирних кислот та інші.
Відповідно до схеми гліколізу, при анаеробному розщепленні глюкози до піровиноградної кислоти синтезуються 10 молекул АТФ. Але на 1-й стадії гліколізу відбувається дворазове фосфорилювання молекул глюкози. Тому у результаті - накопичення енергії під час гліколізу становить вісім макроергічних зв'язків.
Подальше перетворення піровиноградної кислоти за умов наявності кисню відбувається шляхом послідовного відщеплення атомів водню одночасно з електронами, а також СО2, яке здійснюють відповідно дегідрогенази і декарбоксилази. Приєднуючи водень субстрату, дегідрогенази передають його через проміжні системи кисню. Ці перетворення мають циклічний характер (цикл Кребса). У них велике значення належить органічним карбоновим кислотам, які є вихідними компонентами для різних синтетичних процесів. За повний оборот циклу Кребса відбувається окислення п'яти пар атомів водню. З них дві пари беруться від молекули піровиноградної кислоти, а інші три пари атомів водню - від води, яка приєднується до кислот на протязі циклу.
Перетворення піровиноградної кислоти у циклі Кребса супроводжується вивільненням енергії. Переміщення електронів по ланцюзі супроводжується фосфорилюванням. Утворені при окисленні кожної молекули піровиноградної кислоти п'ять пар електронів викликають синтез у середньому 30 молекул АТР. Трансформована під час аеробної фази енергія становить: 10 ккал · 30 = 300 ккал. В одному молі глюкози вміщено 674 ккал енергії. Отже, енергетична ефективність цього процесу становить близько 52%.
7. Цикл гліоксалевої кислоти.
Гліоксилатний цикл локалізований не в мітохондріях, як цикл Кребса, а у спеціалізованих мікротілах - гліоксисомах. На відміну від циклу Кребса, у гліоксилатному циклі бере участь не одна, а дві молекули ацетил-СоА. При цьому ацетил-СоА використовується не для окислення, а для синтезу бурштинової кислоти.
Із щавлевооцтової кислоти і ацетил-СоА синтезується лимонна кислота, потім утворюються цис-аконітова та ізолимонна, як і в циклі Кребса.
Потім ізолимонна кислота при участі ферменту ізоцитратліази розпадається на гліоксилеву і бурштинову кислоти. Гліоксилева кислота під дією малатсинтетази взаємодіє з другою молекулою ацетил-СоА, що призводить до утворення яблучної кислоти, яка потім перетворюється у фосфоенолпіровиноградну кислоту, а остання у вуглеводи.
8. Пентозофосфатний цикл.
У рослинній клітині поряд з розпадом глюкози шляхом гліколізу і в циклі Кребса, який є головним постачальником енергії, відбувається і другий важливий спосіб її перетворення - пентозофосфатний цикл.
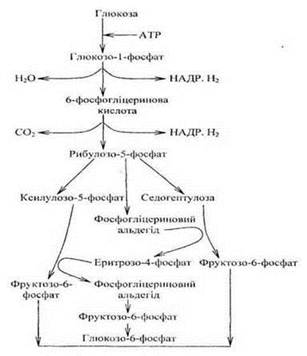
У даному випадку розщепленню глюкози передує утворення и монофосфорного ефіру, а потім нібито відсічення першого вуглецю у ланцюзі.
Відповідно до цього пентозофосфатний цикл часто називають апотомічним окисленням на відміну від гліколітичного циклу, який називають дихотомічним за утворення у ньому двох тріоз.
Окислення глюкози пентозофосфатним шляхом відбувається в цитоплазмі клітини, а також у пластидах. Особливо висока активність пентозофосфатного шляху дихання у клітинах і тканинах з активними синтетичними процесами. Розщеплення глюкози, як і при гліколізі, розпочинається з її фосфорилювання. За допомогою ферменту гексокінази та АТФ утворюються глюкозо-6-фосфат і АДФ. У реакціях пентозофосфатного циклу беруть участь шість молекул глюкози, п'ять із яких регенеруються, а одна окислюється згідно рівняння:
С6Н12О6 + 6О2 → 6СО2 + 6Н2О + 2867 кДж.
У цьому циклі можна виділити два етапи:
а) окислення глюкози;
б) регенерація вихідного субстрату.
Реакції першого етапу каталізує дегідрогеназно-карбоксилювальна система з трьох ферментів. Спочатку відбувається дегідрування глюкозо-6-фосфату за участю ферменту глюкозо-6-фосфатдегідрогенази, в активній групі якого є НАДР - акцептор електронів. Утворена 6-фосфоглюконова кислота зазнає окислювального декарбоксилювання і дегідрування до рибулозо-5-фосфату.
Другий етап циклу пов'язаний із регенерацією глюкозо-6-фосфату, Із рибулозо-5-фосфату під дією ізомераз утворюється рибозо-3-фосфат. Потім із двох фосфопентоз шляхом рекомбінації при послідовній участі ферментів транскаталази і трансальдолази утворюється спочатку семи-вуглецевий цукор седогептулоза і фосфогліцериновий альдегід, потім - еритрозофосфат і фруктозо-6-фосфат. Унаслідок ізомеризації фруктозо-6-фосфат переходить у глюкозо-6-фосфат.
Для кожного оберту підсумкове рівняння пентозофосфатного циклу має вигляд:
6Г-6-Ф+12НАДФ7 Н,0 > 5Г-6-Ф+6СО2+12НАДФ2+Н3РО4.
Як випливає з цього рівняння, при повному окисленні однієї молекули глюкозо- 6-фосфату утворюється 12 молекул НАДФН2. Окислення 12 пар протонів від НАДФ·Н2 у процесі окислювального фосфорилювання забезпечує синтез 36 молекул АТФ що становить 41,87кДж*36 = 1507кДж/моль і практично не поступається енергетичному виходу гліколітичного шляху дихання.
Окислення глюкози пентозофосфатним шляхом виявлено в різних органах рослини. Воно є головним джерелом утворення пентоз, які використовуються клітиною для синтезу нуклеїнових кислот. Цей цикл також постачає рибозу, яка у формі рибулозодифосфату служить акцептором вуглекислого газу у темновій фазі фотосинтезу.
9. Окислювальне фосфорилювання.
Трансформація енергії, яка вивільнюється у процесах дисиміляції, відбувається двома шляхами:
1) фосфорилювання на рівні субстрату при окисленні дегідрогеназами;
2) окислювальне фосфорилювання у субмітохондріальних системах.
Фосфорилювання на рівні субстрату. При цьому типі фосфорилювання вільна
енергія біологічного окислення уловлюється у формі багатих енергією зв'язків між субстратом і фосфатом. Найбільш вивченим прикладом фосфорилювання на рівні субстрату є одна з реакцій гліколізу:
ФГА + НАД+ → ФГК + НАД-Н + Н+.
Процес відбувається у два етапи з утворенням проміжного продукту дифосфогліцеринової кислоти, у молекулі якої є багатий на енергію ацилфосфатний зв'язок. На другому етапі залишок фосфорної кислоти з макроергічними зв'язками переноситься на АДФіз збереженням енергії.
Окислювальне фосфорилювання у субмітохондріальних системах. Енергія, яка вивільнюється при розщепленні органічних речовин, перетворюється в енергію фосфатних зв'язків АТФ Утворення АТФвідбувається за схемою:
АДФ + Р неорг. → АТФ.
Фосфорилювання АДФ з утворенням АТФ відбувається лише у процесі окислення речовин. Цей процес має назву окислювального фосфорилювання. Окислювальне фосфорилювання у дихальному ланцюзі відбувається за рахунок поступового руху електронів між переносниками ЕТЛ. Якщо ж синтез АТФ з якихось причин (дефіцит АДФ) неможливий, то рух електрона по ЕТЛ зупиняється. Це явище має велике значення у регулюванні окисного обміну і називається дихальним спряженням. Згідно гіпотези хімічного спряження, у процесі фосфорилювання, тобто під час руху електрона по ЕТЛ, виникають проміжні, багаті на енергію сполуки, так звані, фактори спряження.
10. Дихальний коефіцієнт і його залежність від природи окислювального субстрату.
Уявлення про хімічну природу субстрату, який піддається окисленню, дає дихальний коефіцієнт. Дихальний коефіцієнт (ДК) характеризує співвідношення об'ємів виділеного під час дихання вуглекислого газу та поглинутого кисню:
![]()
Згідно закону Авогадро, одна грам-молекула будь-якого газу займає однаковий об'єм. Тому при окисленні глюкози відповідно з реакцією С6Н12О6 + 6О2 → 6СО2 + 6Н2О дихальний коефіцієнт дорівнює:
![]()
тобто кількість молекул виділеного вуглекислого газу відповідає кількості атомів вуглецю у молекулі субстрату, а число поглинутих молекул кисню зростає зі збільшенням кількості атомів водню і зменшується зі збільшенням числа атомів кисню у молекулі, що окислюється. Тому при окисленні жирів і білків, молекули яких містять відносно багато атомів водню і мало атомів кисню, дихальний коефіцієнт буде меншим від одиниці (для жирів - близько 0,7, білків - близько 0,8).
Наприклад, при окисленні пальмітинової жирної кислоти (компонента жирів), дихальний коефіцієнт буде дорівнювати:
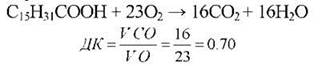
Якщо дихання відбувається за рахунок органічних кислот, то ДК буде вищим за одиницю:
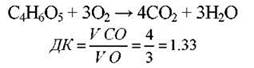
На величину ДК можуть виливати і процеси обміну речовин, які не мають відношення до дихання. Так, при проростанні олійного насіння жирні кислоти перетворюються у вуглеводи, тому дихальний коефіцієнт знижується:
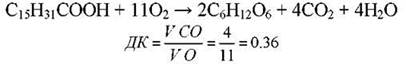
В умовах нестачі кисню процес аеробного дихання може супроводжуватися анаеробним диханням, при якому також виділяється вуглекислий газ. Тому у цьому випадку дихальний коефіцієнт буде зростати. Подібне можна спостерігати при зануренні проростаючого насіння у воду. Утворені продукти неповного окислення при певних концентраціях можуть виявляти токсичну дію.
Таким чином, величина дихального коефіцієнта відображує не лише тип субстрату, який піддається окисленню, але й особливості процесу дихання даної тканини чи органа відповідно до їх стану та впливу конкретних зовнішніх умов.
11. Дихальний газообмін рослин як фактор продукційного процесу. Взаємозв’язок процесів фотосинтезу і дихання
Рівень продукційного процесу значною мірою залежить від інтенсивності дихання, а саме: від його складових - дихання росту і дихання підтримування. Процес фотосинтезу є першим етапом циклу обміну енергії і речовини в клітині. Утворені у світлових реакціях багаті на енергію активні метаболіти - АТФ, НАДФ·Н2 у темнових фазах перетворюються у багаті на енергію але у малоактивній формі запасні сполуки - білки, вуглеводи, жири. Щоб клітина могла використати нагромаджену у вуглецевих сполуках енергію, необхідно цю енергію трансформувати у енергію макроергічних фосфатних зв’язків АТФ. Саме це відбувається під час дихання, яке є другим етапом енергетичного обміну рослин.
Через це обидва процеси - фотосинтез і дихання, пов’язані з перетворенням енергії і утворюють єдиний енергетичний цикл.
Процеси фотосинтезу і дихання, на перший погляд, здаються протилежними. Процес фотосинтезу відбувається при освітленні у хлоропластах, нагромаджується енергія, відбувається поглинання СО2 з його відновленням у органічних речовинах, також відбувається виділення кисню.
Дихання не залежить від світла, відбувається у мітохондріях. При диханні органічні речовини окислюються з виділенням енергії, СО2 і води.
Однак ці процеси мають багато спільного. Обов’язковими учасниками є вода: при фотосинтезі вона є донором Н2, а при диханні окислення речовин відбувається за рахунок кисню води.
Фотосинтез і дихання мають однакові проміжні сполуки - ФГК, ФГА, ФДА, рибу лозу, Ф-1,6-Ф.
Але є також і відмінності. Дихання та фотосинтез постачають АТФ. Але у процесі фотосинтезу джерелом енергії для синтезу АТФ є енергія сонячних променів, а при диханні - енергія окислення органічних речовин. Фотосинтез властивий лише організмам, що мають хлоропласти, тобто це унікальний процес. Дихання, навпаки, є універсальним процесом, притаманним усім живим організмам.