ФІЗІОЛОГІЯ ТА БІОХІМІЯ РОСЛИН
Опорний конспект лекцій
5. ФОТОСИНТЕЗ
Шлях вуглецю (темнова фаза ф/с)
У результаті фотохімічних реакцій у хлоропластах утворились АТФ і НАДФН. Однак самі по собі ці продукти світлової фази не здатні відновити СО2 і , очевидно, темнова фаза ф/с — складний процес із великою кількістю реакцій. У даний час відомі С3-, С4- шляхи фіксації СО2, фотосинтез за типом сукулентів (САМ-метаболізм) і фотодихання. САМ-метаболізм (Crassulacean AcidMetabolism) ще часом називають метаболізмом органічних карбонових кислот у товстолистих - МОКТ.
С3-шлях ф/с (цикл Кальвіна)
Американському біохіміку Кальвіну і його співробітникам за допомогою міченого 14СО2 вдалося виявити первинний продукт ф/с. Ним виявилася 3-фосфогліцеринова кислота (ФГК). Згодом ними ж було виявлено первинний акцептор СО2 - рибульозо-1,5-дифосфат і всі інші реакції С3-шляху ф/с. Цикл Кальвіна складається з трьох етапів: карбоксилювання, відновлення та регенерації.
1. Карбоксилювання (схема: С5 + СО2 ⇒ С6⇒ 2С3):
1) рибульозо-5-фосфат з участю АТФ фосфорилюється і перетворюється на рибульозо-1,5-дифосфат - фосфорибульозокіназа;
2) поглинається СО2, що приєднується до рибульозо-1,5-дифосфату — рибульозо-дифосфаткарбоксилаза. Отриманий продукт (2-карбоксі-3-кетоарабінітол-1,5-бісфосфат) зразу ж розкладається на 2 молекули 3-фосфогліцеринової кислоти, при цьому засвоюється вода.
2. Відновлення (схема: ФГК ⇒ ФГА):
3) 3-ФГК перетворюється на 1,3-дифосфогліцеринову кислоту — фосфогліцераткіназа + АТФ;
4) 1,3-диФГК за рахунок водню НАДФН2 і тріозофосфатдегідрогенази відновлюється й утворює 3-фосфогліцериновий альдегід;
3. Регенерація (схема:
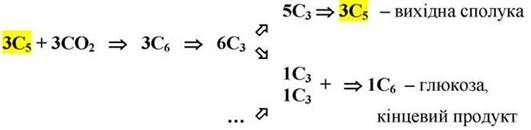
Регенерація первинного акцептора диоксиду вуглецю і синтез кінцевого продукту ф/с. В результаті вищеописаних перетворень при фіксації 3-х молекул СО2 і утворенні 6-ти молекул відновлених фосфотріоз, 5 із них ідуть на регенерацію рибульозо-5-фосфата, а одна - на синтез глюкози:
5) 3-ФГА під дією ферменту тріозофосфатізомерази перетворюється на фосфодіоксіацетон;
6) 3-ФГА і фосфодіоксіацетон конденсуються - альдолаза - з утворенням фруктозо-1,6-дифосфата;
7) від фруктозо-1,6-дифосфату відщеплюється 1 фосфат і утворюється фруктозо-6-фосфат - гексозофосфатаза;
8) транскетолаза каталізує перенесення дво-вуглецевого задишка від кетози (фруктозо-6-фосфат) на альдозу (3-ФГА). В результаті утворюється: еритрозо-4-фосфат і ксилульозо-5-фосфат;
9) трансальдолаза сполучає еритрозо-4-фосфат (альдоза) із фосфодіоксіацетоном, і утворюється седогептульозо-1,7-дифосфат;
10) останній під дією фосфатази та води дефосфорилюється й утворюється седогептульозофосфат;
11) під дією транскетолази з нього і 3ФГА утворюються 2 молекули пентоз: ксилульозо-5-фосфат тарибозо-5-фосфат;
12) рибозо-5-фосфат з участю рибозофосфатізомерази перетворюється на рибульозо-5-фосфат;
13) 2 молекули ксилульозо-5-фосфату з участю рибульозофосфатепімерази перетворюються на 2 молекули рибульозо-5-фосфату. З цих трьох молекул рибульозо-5-фосфату (12 і 13 реакції) починається новий цикл фіксації СО2;
14) із шостої молекули 3ФГА під дією альдолази синтезується (при повторенні циклу) молекула фруктозо-1,6-дифосфату, із якої можуть утворюватися глюкоза, сахароза або крохмаль.
Лише 2 реакції у фотосинтетичному циклі є специфічними для фотосинтезуючих рослин, це - перша і друга. Таким чином, першим продуктом ф/с є ФГК, а кінцевим - фруктозо-6-фосфат. Для синтезу однієї молекули глюкози в циклі Кальвіна необхідно 12НАДФН і 18АТФ, які постачаються фотохімічними реакціями світлової фази.
С4—шлях ф/с (цикл Хетча і Слека).
Цілим рядом досліджень 60-х рр. (Карпілов, Корчак, Незговорова, австралійські вчені Хетч, Слек) було відкрито С4-шлях вуглецю в ф/с.
До групи рослин із С4-шляхом ф/с належать цукрова тростина, кукурудза, сорго, амарант, лобода та інші. Листки цих рослин містять 2 типи хлоропластів: звичайні хлоропласти в клітинах мезофілу і великі хлоропласти, часто без гран, із клітин обкладки (обгортки) провідних пучків.
СО2 через продихи потрапляє до цитоплазми клітин мезофілу і вступає в реакцію з фосфоенолпіруватом (ФЕП) за участю ФЕП-карбоксилази, утворюючи щавлевооцтову кислоту (оксалоацетат).
Далі в хлоропластах щавлевооцтова кислота відновлюється до яблучної кислоти (малату) за рахунок НАДФН, що утворився у світловій фазі ф/с. Щавлевооцтова кислота при наявності NH4+може перетворюватися в аспартат.
Згодом малат (або аспартат) переноситься до хлоропластів клітин обкладки судинного пучка, де він декарбоксилюється малик-ензимом (малатдегідрогеназою декарбоксилюючою) до пірувату і СО2. При цьому утворюється НАДФН.
СО2, який утворився при окиснюючому декарбоксилюванні малату (чи аспартату), приєднується до рибульозо-дифосфату з наступним утворенням 3ФГК — тобто в цих хлоропластах фіксація СО2 здійснюється за типом циклу Кальвіна, а в процесі циклічного фотофосфорилювання (ФСІ) синтезується велика кількість АТФ.
Утворений піруват поступає назад до хлоропластів клітин мезофілу, де може знову перетворюватися в первинний акцептор СО2 - ФЕП.
Така компартментація процесів дозволяє рослинам із С4-шляхом здійснювати ф/с навіть при закритих продихах, оскільки хлоропласти клітин обкладки використовують малат (аспартат), який утворився раніше, як донор СО2. С4-рослини можуть також використовувати СО2, утворений під час фотодихання. Закриття продихів у найспекотніший час скорочує втрати води за рахунок транспірації. Ефективність використання води у С4-рослин у два рази вища, ніж у С3-рослин. С4-рослини стійкі до засолення, тому мають переваги у посушливих та засолених місцях.
Фотосинтез за типом товстолистих (сукулентів)
Для сукулентів характерний добовий цикл метаболізму С4 кислот з утворенням яблучної кислоти вночі. Цей тип ф/с часто називають САМ- метаболізм.
Уночі СО2 поступає до листків, де за участю ФЕП-карбоксилази в цитоплазмі взаємодіє з фосфоенолпіруватом (ФЕП), утворюючи оксалоацетат. Джерелом ФЕП служить крохмаль. Теж саме відбувається із СО2, звільненому при диханні. Оксалоацетат відновлюється до яблучної кислоти (НАДН-залежна малатдегідрогеназа), яка накопичується у вакуолях.
Вдень, коли продихи закриті, малат транспортується з вакуолі до цитоплазми і там декарбоксилюється (малик-ензим) з утворенням СО2 та пірувату. СО2 надходить до хлоропластів, де і захоплюється в цикл Кальвіна.
Таким чином, при САМ-метаболізмі фіксація СО2 з утворенням малату (вночі) та декарбоксилювання малату з вивільненням СО2 і пірувату (вдень) розподілені в часі. У С4-рослин ці ж процеси розподілені у просторі. Є також значні відмінності в анатомії фотосинтетичних органів, і частково у біохімії даного циклу. Для САМ-рослин характерна більш висока, ніж у попередніх груп рослин, ефективність використання води, однак їхня продуктивність значно нижча.
Фотодихання і метаболізм гліколевої кислоти
Фотодихання (Ф/д) — це активований світлом процес вивільнення СО2 і поглинання О2, який відрізняється від дихання мітохондрій. У функціональному плані до дихання цей процес прямого стосунку не має.
Рибульозодифосфаткарбоксилаза в хлоропластах може функціонувати як оксигеназа, каталізуючи окиснююче розщеплення рибульозодифосфату на 3-ФГК і 2-фосфогліколеву кислоту. Молекули О2 і СО2 конкурують між собою в каталітичному центрі рибульозодифосфаткарбоксилазиоксигенази.
Ф/д здійснюється в результаті взаємодії трьох органел - хлоропластів, мітохондрій і пероксисом.
Гліколат із хлоропласта надходить до пероксисоми і там окиснюється гліколатоксидазою до гліоксилату. Перекис водню розкладається каталазою. Гліоксилат амінується і перетворюється в гліцин.
Гліцин транспортується до мітохондрії. Тут із двох молекул гліцину утворюється серин і звільняється СО2. Серин може знову поступати до пероксисоми і там передавати свою аміногрупу на піруват. Із пірувату виникає аланін, а із серину - гідроксипіруват, який відновлюється в гліцерат.
Гліцерат знову може потрапити до хлоропластів, де шляхом фосфорилювання перетворюється на 3-ФГК і вступає в реакції циклу Кальвіна.
Таким чином, існують різні шляхи відновлення СО2: С3- і С4-шляхи, САМ- метаболізм і фотодихання.