БОТАНИКА ТОМ 1 - КЛЕТОЧНАЯ БИОЛОГИЯ. АНАТОМИЯ. МОРФОЛОГИЯ - 2007
3. ТКАНИ ПОБЕГОВЫХ РАСТЕНИЙ
3.2. Постоянные ткани
В постоянных тканях в норме больше не происходит клеточных делений, их прошедшие дифференцировку клетки больше не способны расти и нередко даже отмирают, заполняясь водой или воздухом Открытая организация растений приводит к тому, что крупные многолетние растения содержат множество мертвых клеток Например, в стволе старого дерева содержится минимальное число живых клеток, а древесина и кора состоят преимущественно из мертвых клеток (Напротив, у животных постаревшие или отмершие клетки в норме быстро элиминируются ). Меристематические клетки примыкают друг к другу плотно, без межклетников, часто они имеют форму неправильных 14-гранников. При переходе в постоянную ткань клетки обычно активно увеличиваются путем постэмбрионального растяжения: клеточная стенка под действием временно нарастающего тургорного давления растягивается необратимо (пластически). Результатом этого является тенденция округления клетки. Соседние клеточные стенки, особенно в углах и вдоль линий смыкания клеток, отрываются друг от друга по менее прочным срединным пластинкам, и возникают заполненные воздухом межклетники (рис. 3.7; 3.8). Эти сначала узкие щели расширяются, соединяются друг с другом и образуют, наконец, связную систему межклетников. Она сообщается с наружным воздухом через устьица или чечевички (см. 3.2.2.2) и обеспечивает газообмен. Межклетные пространства возникают или схизогенно (см. рис. 3.29) путем расщепления клеточных стенок по срединным пластинкам (от греч. schizein — расщеплять), либо путем растворения клеток или клеточных комплексов (лизигенно — см. рис. 3.30, D, Е), или, наконец, путем разрывов ткани (рексигенно) вследствие неравномерного роста (например, стебли многих растений с сердцевинными полостями; см. рис. 4.41). Учитывая удельный объем межклетников, говорят о плотных или рыхлых тканях. Примерами плотных тканей служат покровные и механические ткани, рыхлых — в наибольшей степени хлоренхима.
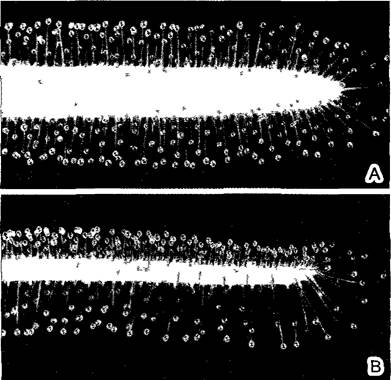
Рис. 3.7. Межклетники (В, С—СЭМ-микрографии)
А—паренхима воздушного корня орхидеи Vanda с узкими межклетниками между округлыми клетками (90х), В—губчатая паренхима (см 4 3 1 3) в листе Parthenocissustricuspidata,между звездчатыми клетками большие межклетные пространства (160х), С—«звездчатая паренхима», белая сердцевинная ткань ситника Juncus, некоторые границы клеток указаны стрелками, межклетники намного превышают по объему очень рыхлую клеточную ткань (230х)
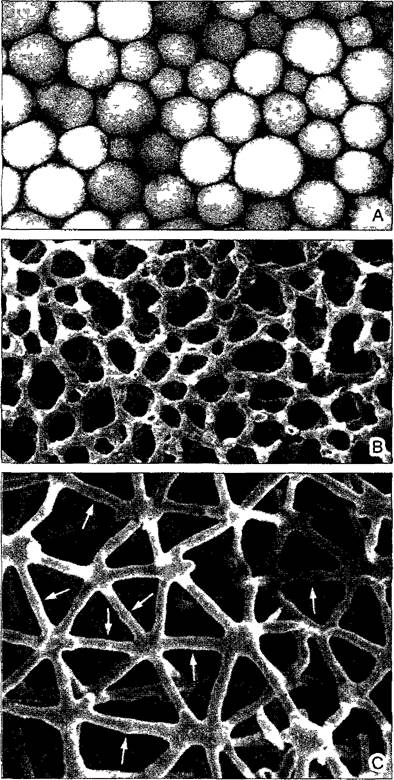
Рис. 3.8. Ткань проветривания (аэренхима, СЭМ-микрографии).
А—воздухоносные ходы в стебле водяной сосенки Hippuris vulgaris, растение укореняется под водой и возвышается над уровнем воды (53х), В—ткань проветривания в черешке листа кувшинки Nymphaea alba (55х)
3.2.1. Паренхима
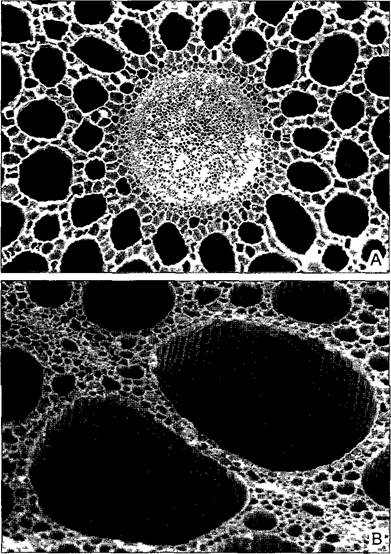
Основная ткань, или паренхима (от греч. para enchyma — масса, заполняющая внутренние промежутки), — это наименее специализированная ткань тела растения. Если из корня, побега или листа мысленно удалить все специализированные ткани, такие как проводящие, покровные и механические, то останется паренхима как основная масса (заполняющая ткань) этих органов. У травянистых растений она составляет главную массу вегетативного тела, потеря тургора в паренхиме из-за недостатка воды ведет к увяданию таких растений. Паренхима состоит в общем случае из крупных, изодиа- метричных («паренхиматических») и тонкостенных клеток. Значительная часть объема основной ткани приходится на межклетники (см. рис. 3.7).
Наряду с мнением, что паренхима мало специализирована, существует представление о функциональном многообразии основной ткани. Особо можно отметить следующие функции.
• Запасающая паренхима служит накоплению органических резервных веществ (полисахариды: крахмальные зерна; полипептиды: кристаллы протеинов; липиды: жирные масла в олеосомах). Такая паренхима доминирует в «мясистых» запасающих органах, таких как корнеплоды, клубни и луковицы, а также в запасающей ткани семян. Запасные вещества часто также откладываются в сердцевинной паренхиме и паренхиме первичной коры. В стволах древесных растений запасающую функцию берет на себя древесинная паренхима, которая обычно пронизывает мертвую древесину как сеть.
✵ Гидренхима: растения очень сухих местообитаний, которые остаются активными и при длительно продолжающемся дефиците воды, запасают воду в вакуолях чрезвычайно крупных паренхимных клеток (диаметром до 0,5 мм). Соответствующие органы внешне заметно вздуты, их объем увеличен, а поверхность уменьшена. В крайних случаях они приближаются к форме шара. Это явление обозначают как суккулентность (от лат. succus — сок). Общеизвестные примеры — это листья очитка (Sedum — см. рис. 4.70) и стебли кактусов (см. бокс 4.1, рис. А; рис. 4.34; 4.35).
• В аэренхиме (ткань проветривания; от греч. aerios — воздушный) сильно развита система межклетников; до 70 % объема ткани приходится на межклеточные воздушные пространства. У болотных и водных растений она обеспечивает газообмен в погруженных органах, так как система межклетников достигает устьиц в плавающих или торчащих из воды листьях или побегах (см. рис. 3.8).
• Хлоренхима (ассимиляционная паренхима) содержит множество хлоропластов. Эта ткань листа (мезофилл) специализирована на фотосинтезе. В палисадном слое мезофилла клетки вытянуты перпендикулярно поверхности листа (см. рис. 4.64). Губчатая паренхима является одновременно и хлоренхимой, и аэренхимой. Клетки этой очень рыхлой ткани неправильно- звездчатые (см. рис. 3.7). Изобилие крупных межклетников создает условия для сильной потери воды губчатой паренхимой, поэтому эта ткань — главный орган1транспирации.
1 По определению, ткань не может быть органом. — Примеч. ред.
3.2.2. Пограничные ткани
Органы травянистых растений и неодревесневших частей древесных растений снаружи покрыты обычно однослойной первичной покровной тканью — эпидермой1
(от греч. epi derma — наружная кожа). Когда она разрывается при вторичном утолщении побегов и корней2 или при повреждениях, то замещается многослойной вторичной покровной тканью, т. е. пробкой (феллема, греч. phellos — пробка). Пробку образует особый камбий — пробковый камбий, или феллоген. Клетки пробки отмирают после отложения в стенках слоев суберина, таким образом, пробка — мертвая ткань (см. 3.2.2.2). В стволах деревьев и толстых многолетних ветвях, и корнях повторяющиеся разрывы пробковых слоев приводят к многократному заложению пробкового камбия и слоев пробки и тем самым в итоге — к образованию толстых омертвевших клеточных масс, которые обозначают как корка (см. 4.2.8.9).
1 Эпидермой покрыты побеги или талломы. На корнях развивается особая первичная покровная ткань ризодерма, или эпиблема, а эпидермы нет. — Примеч. ред.
2 См. предыдущее примечание. — Примеч. ред.
Общую особенность покровных тканей представляет плотное расположение клеток; межклетники отсутствуют. Боковые контакты клеток эпидермы и пробки очень прочные, так что часто эпидерму листа или слой пробки можно, как шкурку, снять с нижележащих тканей (рис. 3.9). Жизненноважный газообмен с наружной средой осуществляется в эпидерме через устьица, способные регулировать его интенсивность, а в пробке — через находящиеся в ней чечевички.
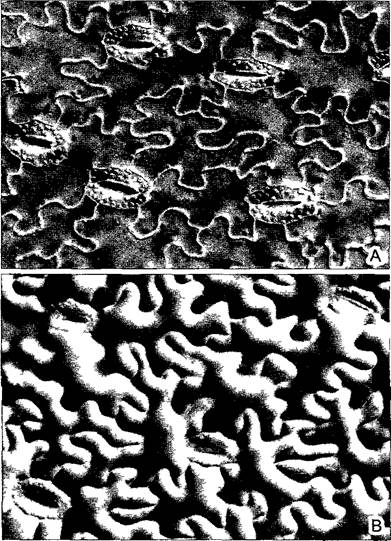
Рис. 3.9. Однослойная эпидерма калужницы болотной Caltha palustris, снятая с нижней стороны листа (интерференционный контраст, 230х)
А—клетки прочно соединены друг с другом, так как плотно сцеплены заходящими друг за друга выступами Содержащие многочисленные хлоропласты замыкающие клетки устьиц — типичные идиобласты в этой однородной ткани (ср также рис 3 13, Е, F), В—отчетливо видно обусловленное тургором подушкообразное вздутие отдельных клеток
Пластиды эпидермы в большинстве случаев представлены лейкопластами или недоразвитыми безгранальными хлоропластами. Однако в листочках околоцветников и плодах цитоплазма эпидермальных клеток часто заполнена хромопластами, что служит привлечению животных и тем самым косвенно способствует опылению или же распространению плодов и семян. В других случаях подобный эффект достигается при помощи окрашивающих веществ вакуолярного сока (химохромы: антоциан, бетациан, флавоноиды). Часто обе формы пигментации встречаются комбинированно.
Внутри тела растения также можно встретить пограничную ткань, а именно однослойную эндодерму. Она как тканевой барьер разграничивает и физиологически изолирует проводящие ткани от паренхимы.
3.2.2.1. Эпидерма и кутикула
Молекулярное строение кутикулы (от лат. cutis — кожа) совершенно в отношении снижения ее проницаемости для воды (см. 2.2 7.6). У растений сухих местообита
ний, где содержание воды имеет критическое значение, испарение воды через кутикулу может быть сокращено до менее чем 0,01 % от равной по площади свободной водной поверхности. Напротив, в таких местах, где проницаемость необходима, например, у железистых клеток, — кутикулы пористые. Поверхности всасывающих органов вообще не покрыты кутикулой. В общем случае, это характерно для ризодермы, самого внешнего слоя молодых корней.
Кутикула способна неограниченно расти поверхностно. В противоположность насекомым, кутикула которых имеет общую с кутикулой растений функцию защиты от испарения, у растущих частей растений не происходит линьки. Вместе с растущей эпидермой постоянно растет и кутикула, которая у растений в норме никогда не играет дополнительной роли панциря. Внеклеточные кутиназы делают молекулярно-ячеистый кутиновый матрикс пластически растяжимым и допускают отложение нового кутина и воска.
Нередко поверхностный рост кутикулы опережает таковой эпидермы, что создает кутикулярные складки, которые простираются поверх границ клеток (рис. 3.10). Кутикулярная складчатость уменьшает смачиваемость: водяные капли вследствие высокого поверхностного натяжения могут касаться только внешних кромок кутикулярных гребней и скатываются с эпидермы. Этот эффект часто усиливается при формировании выпячиваний эпидермальных клеток (папиллозные образования). При дожде постоянно скатывающиеся капли воды эффективно очищают поверхность. При этом смываются споры грибов, которые обычно непрочно зацеплены за складки кутикулы. (Многие грибы относятся к опаснейшим врагам растений, часто они вреднее, чем насекомые.)
Рис. 3.10. Кутикулярная исчерченность (СЭМ-микрографии А—W Barthlott и N. Ehler; В, D—W. Barthlott): А—верхняя эпидерма лепестков пупавки Anthémis tinctona (140х); В—то же уViola tricolor, здесь клетки эпидермы еще более выпуклые, чем у Anthemis, «папиллозные» (95х); С—нижняя сторона листа Parthenocissus tncuspidata с устьицем (700х); D—поверхность семени кактуса Neoporteriа brevicylindrica (120х)
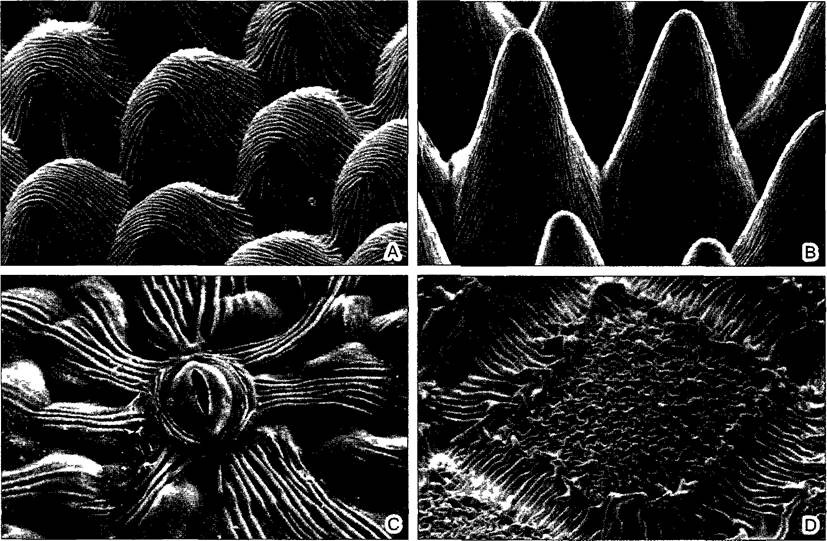
Ту же функцию очистки поверхности может альтернативно выполнять эпикутикулярный воск. Подобные поверхностные чехлы из восковых кристаллов заметны уже невооруженным глазом как голубовато-серый «восковой налет» (например, «сизые» сорта капусты, слив, винограда; особенно мощный у восковой пальмы Сореrnicia, у которой палочковидные кристаллы воска достигают длины до 20 мкм и находят применение как карнауба-воск). Под электронным микроскопом эпикутикулярные воска очень многообразны (рис. 3.11). При этом их форма очень хорошо коррелирует с химическим составом, что объясняется самоорганизацией в соответствии с молекулярными параметрами. Восковой налет не встречается одновременно с кутикулярной складчатостью. Он делает соответствующие поверхности (такие, как верхняя поверхность листа настурции или цветок лотоса) совершенно несмачиваемыми. После стирания налет регенерирует путем новой секреции воска сквозь кутикулу. Молекулы воска при этом перемещаются до внешней поверхности эпидермы, вероятно, с диффундирующей через кутикулу водой.
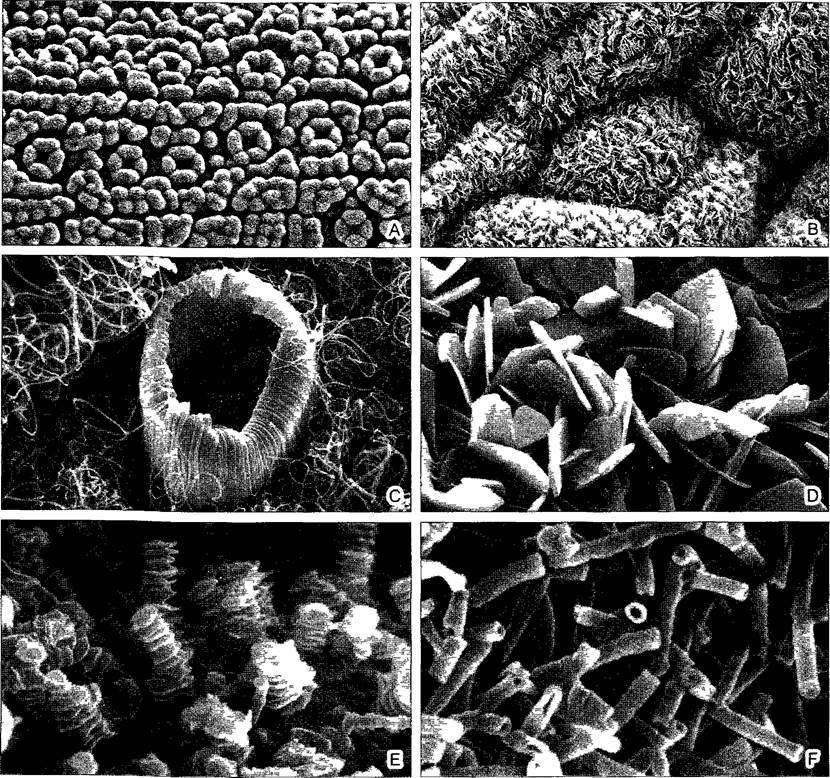
Рис. 3.11. Эпикутикулярный воск (СЭМ-микрографии W Barthlott)
А—нижняя сторона хвоинки тисса Taxus baccata, внешний вид, выпуклые эпидермальные клетки покрыты плотным восковым слоем, который состоит из восковых трубочек (cp F), устьица погружены между 2—4 побочными клетками (230х), В—верхняя сторона листа зверобоя Нуреnсиm buckieyi, частично агрегированные восковые пластиночки (1360х), С—для многих однодольных (здесь дана Hehconia colhnstana) типичны длинные «восковые волоски», вокруг устьица восковой валик (1280х), D— восковые пластиночки у Lecythis chartacea(5400х), Е—поперечно-вальковатые восковые столбики у Wilhamodendron quadnlocelathum, подобные восковые башенки типичны, например, для магнолиевых, лавровых и кирказоновых (6200х), F—восковые трубочки (здесь у жимолости Lonicera tatanca) образуются, если в качестве главных компонентов присутствуют ß-дикетон или 10-нонакосанол (23000х)
Секрецию мономеров кутина и воска неизвестным еще образом всегда стимулирует контакт клеток с воздухом, не насыщенным водным паром. Даже межклетники мезофилла выстланы очень тонким слоем кутина (внутренняя кутикула), выявляемым только при специальных опытах и с применением электронного микроскопа. Чрезвычайно толстые кутикулы с дополнительными кутикулярными слоями встречаются на многолетних листьях и стеблях растений очень сухих местообитаний, например, у кактусов и агав. Подобные кутикулы очень прочны не только в химическом, но и в механическом отношении и могут устоять против жевательных аппаратов мелких животных. В других случаях наружные стенки клеток эпидермы становятся прочными вследствие обызвествления или — чаще — окремнения. Особенно мощное окремнение встречается, например, у злаков и осок. Из-за сильного окремнения хвощи раньше использовали для полировки оловянной посуды («оловянная трава»)
Особенно многообразны по структуре и составу веществ стенки клеток эпидермы плодов и семян. Упомянутые эпидермы в сухом состоянии часто имеют консистенцию рога, но сильно разбухают в воде и становятся при этом мягкими и слизистыми.
Эпидермы некоторых листьев берут на себя функцию гидренхимы, тогда их клетки особенно крупны и могут располагаться многими (до 15) слоями вследствие периклинальных делений клеток протодермы. В других случаях многослойные эпидермы выполняют механическую функцию, как в листьях жестколистных растений (см. рис. 4.71).
Устьичные аппараты (устьица; от греч. — рот) характерны для кутинизированных эпидерм. Они находятся большей частью на нижней стороне листьев, однако не бывает случаев их полного отсутствия на стеблях и околоцветнике. Напротив, на корнях устьиц нет.
Каждое устьице состоит из двух удлиненных замыкающих клеток, которые прочно связаны друг с другом только на своих концах, в то время как их средние части отделены друг от друга схизогенно образованной межклетной щелью — апертурой. Апертура представляет собой проходящий через эпидерму и кутикулу канал между наружным воздухом и особенно крупной межклетной полостью тканей мезофилла или первичной коры, которую, немного вводя в заблуждение, обознача
ют как дыхательную полость1. Ширина апертуры может оперативно регулироваться при помощи изменения формы замыкающих клеток. Щель, ограниченная вентральными стенками замыкающих клеток, открыта тем больше, чем выше их тургор (рис. 3.12; 7.58, А; см. 8.3.2.5). Устьица — основные регуляторы газообмена, особенно транспирации (см. 6.3.4.1; 6 5 7).
1 Этот межклетник принято называть подустьичной полостью — Примеч. ред.
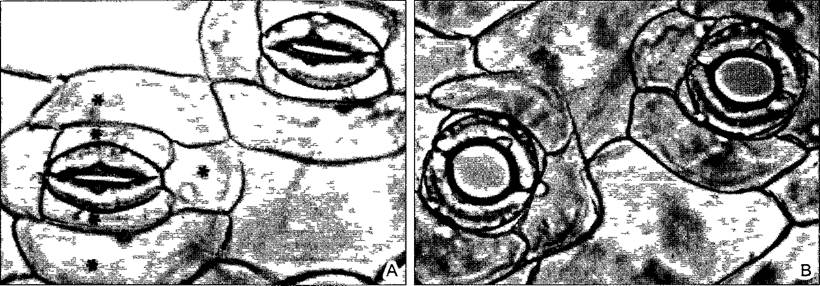
Рис. 3.12. Устьица Commehna communis, утратившие тургор (по К Raschke)
А—в 200 мМ растворе сахарозы; В—в воде в тургесцентном состоянии с широко открытыми устьичными щелями, в этом состоянии замыкающие клетки удлинились и околоустьичные клетки (*) изменили форму (400х)
На нижней стороне листьев, где обнаруживают, как правило, от 100 до 500 устьиц на 1 мм2, занятая апертурами площадь составляет только 0,5 — 2% поверхности листа даже при полностью открытых устьицах. Однако устьичная транспирация может достигать более двух третей от величины эвапорации (испарения со свободной водной поверхности), а с другой стороны, — быть снижена почти до 0.
Замыкающие клетки — это типичные идиобласты эпидермы, от остальных ее клеток они отличаются формой и величиной, а также наличием хлоропластов с ассимиляционным крахмалом. Иногда это (хотя в меньшей мере) свойственно и непосредственно прилегающим к устьицу клеткам, которые обозначают как побочные (= околоустьичные) клетки.Устьице вместе с побочными клетками образует устьичный аппарат. Морфогенетически он соответствует конечному состоянию ме- ристемоида1 (рис. 3.12; 3.13 В, С, Е).
1 Только мезогенные устьичные аппараты, у которых устьице и все окружающие его клетки эпидермы происходят от одного меристемо- ида, можно считать конечными состояниями меристемоидов — Примеч. ред.
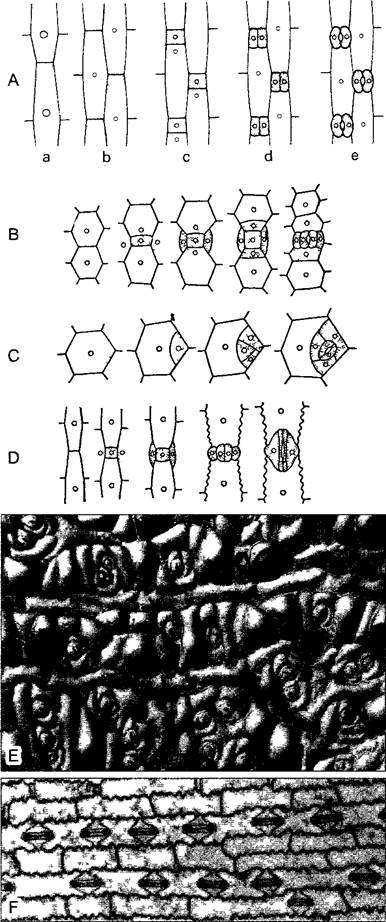
Рис. 3.13. Развитие устьиц у касатика Ins (А), традесканции Tradescantia (В), очитка Sedum (С) и кукурузы Zea mays (D) (A—D по E.Stras- burger и A. de Вагу)
Меристемоиды и устьичные аппараты выделены крапом Е—в качестве примера для С нижняя эпидерма листа Sedum maximum с группами устьиц и околоустьичных клеток между частично папиллозными эпидермальными клетками (60х). F—нижняя эпидерма листа кукурузы (75х)
У злаков замыкающие клетки имеют вид гантелей. Их средние части, узкопросветные из- за сильного утолщения, клеточных стенок и стабильные по форме, при увеличении тургора отодвигаются друг от друга благодаря тому, что тонкостенные, пузыревидные концы клеток раздуваются Максимальное раскрывание апертуры устьиц этого злакового типа мало (пшеница. 7 мкм).
Наряду с этим типом имеются и другие, среди которых тип хвойных выделяется особой сложностью. Устьица на их хвоинках глубоко погружены (см рис. 4 65), в их, обусловленных тургором, движениях активно участвуют побочные клетки с обычно очень неравномерно утолщенными и частично одревесневшими стенками.
Устьицам, выполняющим функции воздуховодов, гомологичны имеющиеся у некоторых растений водяные устьица, или гидатоды, которые служат для выделения капельножидкой воды (гуттация; лат. gutta — капля) (см. 6.3.4.2). Об их наличии свидетельствуют, например, кажущиеся капли росы на листьях настурции. Если выделяемая вода содержит много гидрокарбоната кальция, как у видов кальцефильных камнеломок (Saxifraga), то на гидатодах образуются белые чешуйки карбоната кальция. Многие нектарники выделяют сахаросодержащий секрет через сходные нектарные устьица.
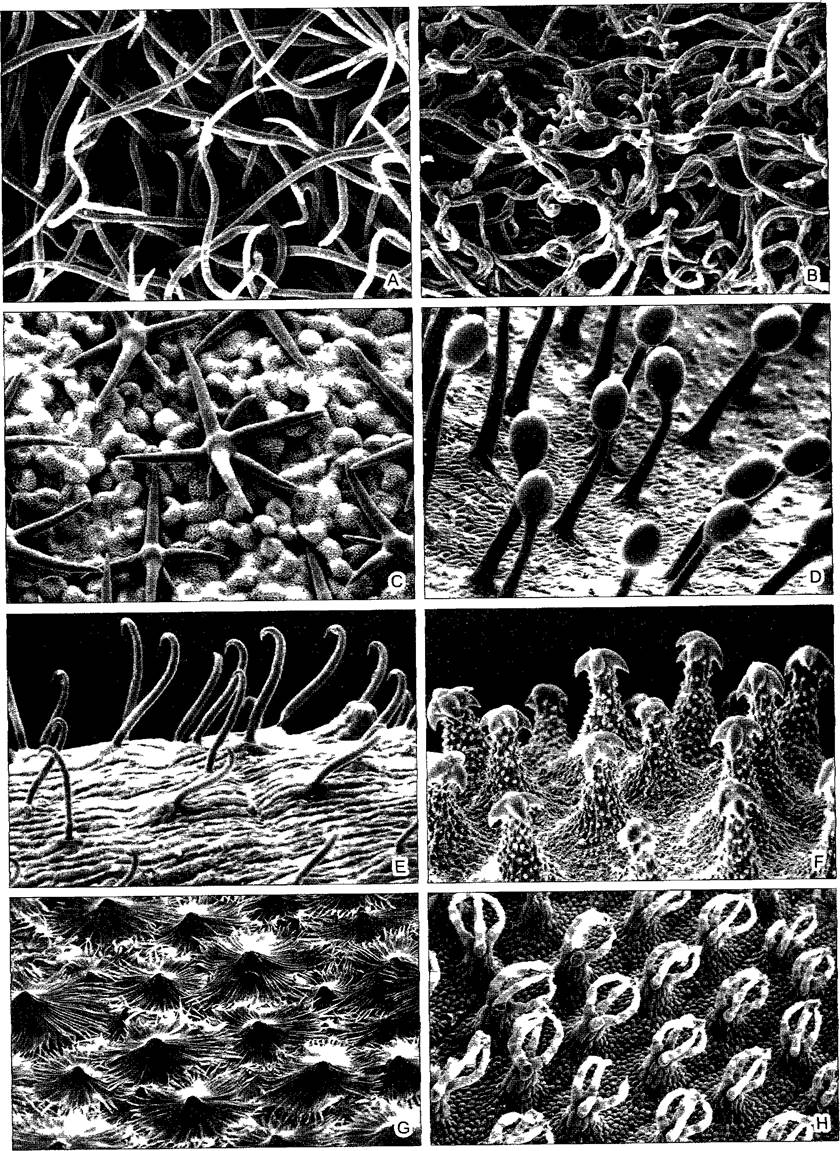
Во многих случаях эпидерма покрыта волосками. Часто отдельные эпидермальные клетки разрастаются в волоски непосредственно или они становятся инициальными клетками меристемоида, так что образуются многоклеточные волоски. Многообразие волосков растений (трихом) исключительно велико, некоторые примеры представлены на рис 3.14. Соответственно многообразны также и их функции. Благодаря волоскам, представляющим по определению идиобласты, эпидерма может далеко выходить за пределы своей первичной функции покровной ткани и принимать участие, например, в процессах поглощения веществ или секреции.
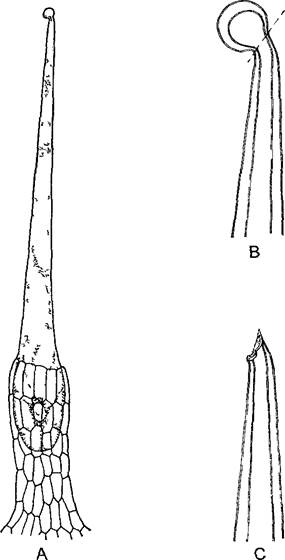
Папиллозные вздутия эпидермальных клеток как линзы фокусируя свет обусловливают блеск поверхности, у околоцветника это может служить для привлечения насекомых. Корневые волоски (см рис 4 75) служат для поглощения веществ.1 Волоски плодов и семян могут способствовать их распространению ветром. Волоски семян представляют собой хозяйственно очень важный хлопок мировое про изводство которого в 1999 г достигло 18 млн т. Волоски хлопчатника, несмотря на свою одноклеточность, достигают 5 см длины и перед отмиранием образуют довольно толстые, состоящие из почти чистой целлюлозы вторичные оболочки с характерной спиральной исчерченностью. Густое шерстистое опушение влияет на транспирацию. Многие растения, обитающие в районах с туманами, при помощи своего опушения поглощают воду из обтекающей их туманной дымки, как и корневые волоски, такие трихомы можно отнести к органам поглощения. Мертвые, заполненные воздухом волоски отражают свет, они кажутся снежно-белыми и могут функционировать как экран от избыточного освещения. В других слу чаях крючковидные цепляющиеся волоски образуются для закрепления на опоре вьющихся или цепляющихся побегов, известные примеры — хмель и подмаренник Galium араrinе. Подобные структуры могут способствовать также распространению плодов и семян. Для защиты нежных листьев от поедания животными служат грубые, часто дополнительно разветвленные щетинистые волоски с пропитанными кремнием твердыми клеточными стенками, даже улитки часто игнорируют растение, если оно представляет собой настоящую зуботерку. Особо изощренный случай — жгучие волоски. Жгучий волосок крапивы (виды Urtica, рис 3.15) — это одна крупная клетка с полиплоидным ядром, которая возвышается на многоклеточном основании (эмергенец, см ниже) над эпидермой листа и стебля. Ее головчато утолщенный кончик при основании отламывается по окремневшему тонкому участку стенки. В этом состоянии жгучий волосок действует как игла шприца, клеточный сок выдавливается и может вызвать болевые ощущения из-за содержания в нем муравьиной кислоты, аце тилхолина и гистамина. Как и во всех крупных клетках, в корневых волосках, растущих волосках хлопчатника и даже в жгучих волосках крапивы (Urtica) можно наблюдать интенсивное движение цитоплазмы, оно было открыто в жгучих волосках Urtica более 300 лет назад (R Hooke, Micrographia1665). Наконец, волоски могут реагировать на раздражения — это чувствительные волоски, например, у венериной мухоловки (Dionaea). Особенно часто встречаются железистые волоски, которые почти всегда несут увеличенную терминальную клетку или многоклеточную головку (см рис 3 14, D, 3 28, D, Е, 9 18)
1 Корневые волоски не гомологичны трихомам побега — Примеч. ред.
Рис. 3.14. Трихомы (СЭМ-микрографии С —F — W Barihlotî, G — С Grunfelder)
А—одноклеточные волоски на нижней стороне листа ежевики (400х), В — гигроскопичные волоски на нижней стороне листа куропаточьей травы Dryas octopetala (350х), С —звездчатые волоски у Vitola surinamensis (Mynsticaceae), дерева дождевых лесов, папиллозные эпидермальные клетки покрыты восковым налетом, волоски без эпикутикулярного воска (285х), D—железистые эмергенцы на листе росянки Drosera capense (65х), Е—крючковидные волоски листьев бобов, «бобовую солому» раньше использовали как средство против вшей и клопов (220х), F—свойство цепляться имеют также якоревидные образования на семенной кожуре чернокорня Cynoglossum officinale (эти образования нельзя относить к трихомам — Примеч. ред. ), из бурачниковых (60х) Это, конечно, не волоски, а эмергенцы, так как в построении этих многоклеточных образований также принимает участие субэпидермальная ткань, у других растений сходные образования представлены волосками {частью даже одноклеточными), G—многоклеточные, концентрические щитовидные волоски Hippophae rhamnoides, которые составляют покров над эпидермой, снижающий транспирацию (160х), Н—пучки волосков на плавающих листьях водного папоротника Salvima natans (50х) Волоски покрыты эпикутикулярным воском, поэтому они делают листовую поверхность несмачиваемой и при принудительном погружении листа увлекают вместе с ним и воздушные пузыри, которые снова вытягивают лист на поверхность воды
Термином эмергенцы обозначает многоклеточные выросты, в возникновении которых участвует также субэпидермальная ткань. Эмергенцы во многом, в том числе по структурному и функциональному разнообразию, соответствуют трихомам, но бывают существенно крупнее. Например, волосовидные железки во многих случаях представлены макроскопическими эмергенцами (см рис 3 31) Мякоть плодов цитрусов (Citrus) образована «внутренними» эмергенцами, которые в виде трубочек с соком врастают в полости завязи. Кроме того, каждый на собственном опыте знает шипы роз и ежевики. Они также являются эмергенцами и поэтому принципиально отличны от колючек, которые являются метаморфизированными листьями (как у барбариса, кактусов) или укороченными побегами (например, у терна). Примечательно, что здесь не совпадают общеупотребительные и специальные названия для ботаников розы не имеют колючек, но имеют шипы, а кактусы не имеют шипов, но имеют колючки.
Рис. 3.15. Жгучий волосок крапивы Urtica dioica (А, 60х) (по D von Denffer)
В—окремневший кончик с предопределенным местом отламывания (400х), С—после отламывания кончика выделяется ядовитый клеточный сок(400х)
3.2.2.2. Пробка
Периклинальный, т е параллельный к внешней или раневой поверхности пробковый камбий (феллоген) откладывает внутрь тонкий, часто содержащий хлоропласты слой паренхимных клеток, — феллодерму. Он становится виден как зеленый слой ткани при обдирании, например, ветвей бузины или стволиков бука. Наружу образуется пробковая ткань — феллема.Полный комплекс тканей — феллоген, феллодерму и феллему — обозначают термином перидерма (см рис 3.17, С)
Часто пробка состоит только из немногих слоев клеток (картофельная кожура, береста молодых стволиков березы), однако пробковый камбий может оставаться активным долгое время и образовать пробку толщиной более сантиметра Самый известный и к тому же имеющий практическое значение пример — пробковый дуб (Quercus súber) Уже 15-летние стволы этого средиземноморского дерева обчищают, те с них снимают уже образовавшуюся перидерму В нескольких слоях от поверхности обчистки возникает новый, особенно активный феллоген, который остается деятельным долгие годы, и производит «бутылочную» пробку Образующуюся пробку снимают примерно раз в 10 лет Среди европейских древесных растений бересклет (Euonymus), а также определенные расы полевого вяза и полевого клена образуют на молодых ветвях крупные пробковые гребни (см. рис. 4.54, А). У бука пробковый камбий остается постоянно активным, так что образуется единый толстый слой пробки вокруг стволов и ветвей.
Опробковение клетки состоит в наложении водонепроницаемого суберинового слоя на оболочку (см. 2.2.7.6). Закончив формирование клеточной оболочки, клетка пробки отмирает и заполняется газом — это случай апоптоза. Вследствие этого пробковая ткань очень легкая, эластичная (ячеистая воздушная подушка!) и является прекрасным изолятором тепла и излучения. Ее можно использовать для звукоизоляции.
Коричневая окраска большинства пробок обусловлена отложением в ней дубильных веществ, которые защищают от проникновения паразитов (насекомых, грибов).
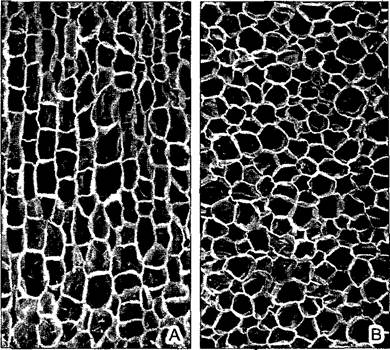
Уже тонкая пробка снижает транспирацию сильнее, чем кутинизированная эпидерма. Как и каждая покровная ткань, пробка не имеет межклетников. Это кажется неожиданным, если учесть, что феллоген образуется как вторичная меристема в паренхиме (например, паренхиме первичной коры стебля), пронизанной системой межклетников. Однако при реэмбрионизации межклетники в плоскости будущего пробкового камбия закрываются из-за локального роста отдельных клеток. Клетки в однослойном феллогене делятся так, что новые клеточные стенки ориентированы исключительно периклинально. Различимое на поперечном срезе расположение клеток четкими рядами и слоями обусловлено синхронными делениями клеток феллогена. Напротив, на тангенциальном срезе еще узнаваемы контуры исходных паренхимных клеток (рис. 3.16).
Рис. 3.16. Пробка: бутылочная пробка, полученная из пробки дуба Quercus súber (СЭМ- микрографии С. Grünfelder).
Слева — поперечный срез, отложенные феллогеном ряды клеток без межклетников; справа — тангенциальный срез, еще узнаваемые контуры исходных клеток паренхимы первичной коры, превратившихся в инициали феллогена (210х)
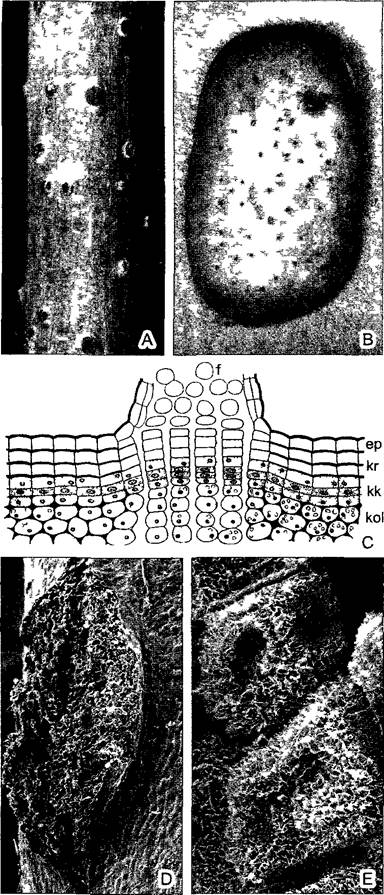
Тотальное опробковение поверхности стеблей сделало бы невозможным жизнь клеток внутри стволов и ветвей из-за удушья. Поэтому пробка местами прервана чечевичками(lenticula — маленькая линза) (рис. 3.17; ср. также рис. 4.55, А). Здесь феллоген откладывает не плотно сомкнутые клетки, а округлые, рыхло расположенные пробковые клетки, между которыми могут диффундировать пары воды, кислород и углекислый газ. Клетки чечевичек, которые в совокупности образуют мучнистую массу, с поверхности плотно покрыты восковыми кристалликами и благодаря этому несмачиваемы. Поэтому при продолжительных дождях чечевички не покрываются водой полностью, но остаются открытыми для газообмена. В бутылочных пробках чечевички должны быть ориентированы поперечно, чтобы герметично закупорить сосуд.
Рис. 3.17. Чечевички (С—по К Маgdefrau D, Е—СЭМ-микрографии С Neinhuis и W. Barthlott)
А—двухлетняя ветвь бузины с чечевичками (1,7х), В—пробка на клубне картофеля с многочисленными чечевичками, С—гистология пробки и чечевички (120х) D— чечевичка Акеbiа quinata (50х) Е—у того же объекта клетки выполняющей ткани чечевички, покрытые воском (1640х), ер—эпидерма, f—клетки выполняющей ткани чечевички, кr—пробка, кк—пробковый камбий, коl—колленхима
Покровную ткань из слабо суберинизированных живых клеток обозначают как кутис1. Иногда при этом речь идет об эпидерме, но чаще о лишенном межклетников клеточном слое непосредственно под эпидермой, который называют гиподермой, или — при наличии поясков Каспари — экзодермой. Кутис часто образуется также при зарубцовывании — образовании листовых рубцов после листопада, опадения плодов и т. п. Запрограммированному отделению подобных органов предшествует образование тонкостенной, подобной камбию отделительной ткани.
1 В настоящее время это понятие вышло из употребления. — Примеч. ред.
3.2.2.3. Эндодерма
Где бы ни образовалась эндодерма (постоянно — в корнях, и нередко — в стеблях и листьях), она заметно отличается от прилежащих снаружи и изнутри тканей. Это связано с ее особой функцией, которую рассмотрим на примере эндодермы корня. Она отграничивает расположенную в центре проводящую ткань (центральный цилиндр) от окружающей паренхимы первичной коры (см рис 4 77 — 4 79).
В эндодерме на стадии первичного строения радиальные клеточные стенки1 (антиклинальные относительно поверхности корня и параллельной ей однослойной эндодермы) химически изменены так, что каждую клетку опоясывает лентовидная область, которую называют пояском Каспари по имени открывшего ее ботаника (ПК, рис 3 18). Он свободен от плазмодесм. Плазматическая мембрана прочно соединена с ним и даже при плазмолизе не отделяется от ПК Сама клеточная стенка в ПК инкрустирована лигнином и липофильными веществами и непроницаема. В этом состоит физиологическая роль ПК и всей эндодермы в зоне всасывания корня поглощенная вода с растворенными минеральными веществами может диффундировать по рыхлым клеточным стенкам паренхимы первичной коры вплоть до эндодермы, т е по всему апопласту снаружи от эндодермы. Общая поверхность клеток паренхимы первичной коры — это единая всасывающая поверхность 2. Внеклеточный, апопластный путь диффузии закрывается лишь ПК эндодермы, вода и ионы попадают в центральный цилиндр только сим- пластически по плазмодесмам (см. рис. 6.22). При этом мембранные транслокаторы избирательно пропускают поступающие ионы. Клетки эндодермы особенно активно перемещают ионы в центральный цилиндр, где они потом снова поступают в апопласт — в мертвые сосуды центрального цилиндра. Эндодерма предотвращает вытекание воды и минеральных веществ из центрального цилиндра.
1 Химически изменены не только радиальные, но и поперечные стенки — Примеч. ред.
2 Всасывающую поверхность представляют только наружные стенки клеток ризодермы и корневых волосков. Под ризодермой развивается слой экзодермы с такими же поясками Каспари, и продвижение водных растворов в глубь корня происходит по симпласту. Эндодерма предотвращает потери воды от неконтролируемой разгрузки кситемы — Примеч. ред.
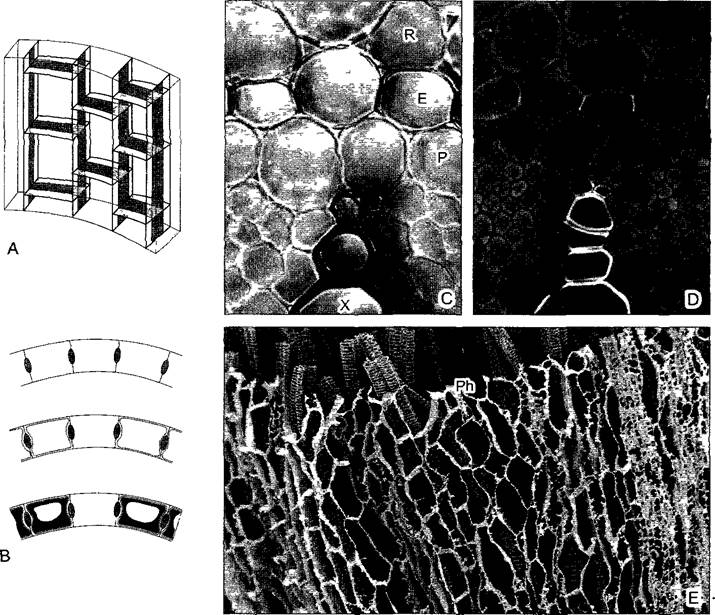
Рис. 3.18. Эндодерма с поясками Каспари (ПК) (С, D—светооптическая фотография I. Dörr; Е—СЭМ-микрография L.Schreiber и R.Guggenheim)
А, В—объемная схема (ПК черные) и на поперечном срезе первичное, вторичное и третичное строение {одна клетка — пропускная); С, D—первичное строение эндодермы корня (Е) Clivia nobihs (350х), Поперечный срез корня, на рисунке ниже эндодермы (в корне — внутри от нее) видна проводящая ткань: С—обработанные флороглюцином с соляной кислотой одревесневшие клеточные стенки или их части (ПК в радиальных стенках эндодермы) окрашены темно; D—флюоресценция в падающем свете после окрашивания акридиновым оранжевым; Е—после энзиматического удаления всех неодревесневших клеточных стенок или их частей в корне кливии (Clivia) (продольно, вид снаружи) сохранились проводящие элементы ксилемы и ПК эндодермы; сетчатая структура, состоящая из ПК, полностью окружает ксилемные элементы центрального цилиндра (11 0х); Е—эндодерма; Р—перицикл; Ph—флоэма; R—первичная кора; X—элементы ксилемы
Ситуация соответствует таковой в эпителиях и эндотелиях высших животных. При этом пояскам Каспари аналогичны плотные контакты («tight junction»), где плотно прилежащие плазматические мембраны соседних клеток склеены друг с другом особыми протеинами. Пояски Каспари и плотные контакты — это образования, которые в разных царствах организмов выработались в процессе эволюции совершенно независимо для выполнения сходных функций и вследствие этого структурно различаются.
За зоной всасывания, в более старых частях корня, клетки эндодермы часто слабо суберинизированы подобно клеткам кутиса («вторичное» строение эндодермы). Наконец, может происходить дополнительное сильное, часто асимметричное утолщение клеточных стенок: «третичная» эндодерма (см. рис. 4.78, В).
Вторичная и третичная эндодермы имеют против ксилемы центрального цилиндра пропускные клетки, которые сохраняют первичное состояние.
3.2.3. Механические ткани
Наземные растения вынуждены иметь клетки с прочными на разрыв и жесткими стенками (см. 2.2.7.4). Ограниченную прочность мелких травянистых растений и нежных органов более крупных растений (листьев, цветков, мясистых плодов) обеспечивает совместное действие тургора и напряжения стенок (тургесценция), что становится ясным при увядании. Напряжения тканей, обусловленные несколько более сильным ростом внутренних частей органов по сравнению с их поверхностью, также могут способствовать упругому состоянию плодов типа ягод. Но такие травянистые или «мясистые» органы в действительности мало прочны, их можно изогнуть, расплющить, растереть. Фактически такой прочности недостаточно растениям более сухих местообитаний и тем более — крупным, особенно многолетним растениям.
Например, силы растяжения и сжатия, которые испытывают корни и стволы высоких деревьев при порывах ветра, далеко превышают те пределы, что способны выдерживать паренхима и покровная ткань. У таких растений функционируют особые механические ткани (стереом; от греч. sterigein— поддерживать). Это плотные, частично мертвые ткани, стенки клеток которых местами или полностью утолщены благодаря отложению особенно богатых целлюлозой слоев оболочки. Вследствие инкрустации некоторыми веществами (большей частью при одревеснении) клеточные стенки могут приобретать дополнительную жесткость и устойчивость к сжатию. Эта возможность реализуется также и в кожуре плодов и семян (орехи, костянки).
Впрочем, в мире животных также выявлены аналогичные принципы обеспечения прочности Гидравлические структуры, прочные на разрыв слои оболочки, напряженные за счет внутреннего давления, преобладают у водных животных и моллюсков, но встречаются, например, и в межпозвоночных хрящевых дисках позвоночных животных У более крупных животных возможно образование сухожилий и скелета путем объединения межклеточных волокон (коллаген, хитин) с инкрустацией или без нее твердыми веществами (карбонаты Са, апатит, продубленные хиноном протеины) Нагрузки от растяжения и сжатия принимают на себя в основном соединительная и опорная ткани, может формироваться также панцирь (хитиновый наружный скелет артропод, черепная коробка позвоночных животных)
Колленхима (от греч. коllа — клей) — это механическая ткань растущих и травянистых частей растения. Ее прозенхимные клетки живые, способные к росту и даже делению. Утолщения оболочки ограничиваются определенными зонами: по ребрам клеток — в уголковой колленхиме (рис. 3.19), отдельными (большей частью периклинальными) продольными стенками — в пластинчатой колленхиме. Это первичная оболочка, ее утолщенные участки сложены перемежающимися ламеллами, состоящими из целлюлозы и пектиновых веществ. Прочность оболочек умеренная, лигнификации не происходит.
Рис. 3.19. Уголковая колленхима в стебле яснотки белой Lamium album, поперечный срез, утолщения оболочек светлые (420х) (световая микроскопия I.Dorr)

Склеренхима (от греч. skleros — твердый, ломкий) — мертвая ткань из очень толстостенных, узкопросветных клеток, которая встречается только в закончивших рост частях растения. Имеются два типа клеток склеренхимы, а именно прозенхимные склеренхимные волокна и изодиа- метричные, или палисадные, каменистые клетки — склереиды.
Тяжи каменистых клеток (см. рис. 2.75) выполняют функции защиты и опоры. Их толстые, заметно слоистые и пронизанные разветвленными поровыми каналами вторичные стенки одревесневают. Склереиды встречаются в твердых оболочках многих плодов и в коре древесных растений,
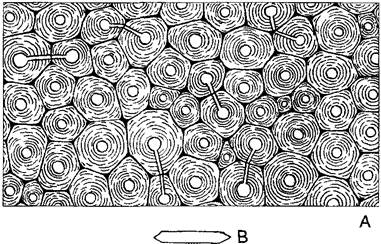
Функции склеренхимных волокон более многообразны (рис. 3.20). В местах напряжения растяжения волокна в основном остаются неодревесневшими (мягкие волокна). в то время как в местах дополнительного сжатия образуются лигнифицированные жесткие волокна. Склеренхимные волокна встречаются прежде всего в стеблях, а часто и в крупных листьях однодольных. Обычно они имеют длину 1 — 2 мм, но некоторые растения содержат существенно более длинные волокна, которые используют в промышленном производстве.
Рис. 3.20. Склеренхимные волокна (А—по Н. Fitting; В, С—по Eames и McDaniels): А—поперечный срез тяжа волокон в листе новозеландского льна Phormtum tenax (360х); В, С—образование древесинного волокна робинии из камбиальной инициали (В) путем двухстороннего верхушечного роста, при этом концы клетки внедряются между соседними клетками (интерпозиция) (150х); D—волокнистые трахеиды в древесине сосны со спиральной текстурой вторичных оболочек (380х)

Исстари использовали в первую очередь лубяные волокна волокнистых растений для изготовления тканей, веревок и канатов. (Обращает на себя внимание языковое родство слов: нем. binden — вязать, Band — лента, повязка, Bast — лыко, луб; соответственно лат. liber — луб, и libellus — переплетенная книга.) Важнейшие волокна из стеблей дают лен (Linum, длина волокон до 7 см), конопля (Cannabis), рами (Boehmeria из сем. крапивных с волокнистыми клетками длиной свыше 50 см), а также джут (Corchorus); сизаль (из агавы) и манильская пенька (из Musa textilis) — волокна из листьев.
Длина склеренхимных волокон всегда превышает размер соседних клеток ткани. Молодые волокнистые клетки растут кончиками, их заостренные концы вдвигаются между другими клетками (интрузивный рост). При этом установившиеся однажды контакты между волокнами и их новым окружением сохраняются и в дальнейшем (интерпозиционный рост). Здесь могут развиваться вторичные плазмодесмы и, наконец, поры. Вследствие параллельного расположения фибрилл во вторичной оболочке волокна его поры щелевидные. Их ориентация позволяет установить направление фибрилл. Оболочки большинства склеренхимных волокон имеют спиральную текстуру (чем приобретают дополнительную эластичность), поэтому щелевидные поры ориентированы косо к оси волокна (рис 3 20, Б)
Не только склеренхимые волокна, но и древесинные части проводящих пучков участвуют в укреплении стеблей, листьев и корней Прочность стволов деревьев, старых ветвей и корней полностью обеспечивает древесина. Между трахеидами — настоящими проводящими элементами древесины — и волокнами есть множество переходов (волокнистые трахеиды — рис 3.20, Б, 4 47, Е)
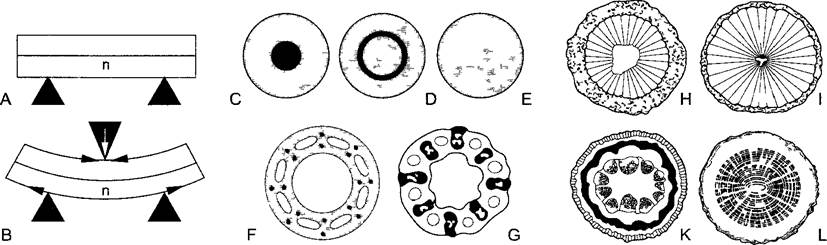
Расположение механических тканей имеет решающее значение для биомеханики отдельных органов и целого растения (рис 3.21). Это особенно отчетливо проявляется в прямостоячих стеблях. В них уже при небольших порывах ветра напряжения изгиба достигают критической величины. Прочность стебля на изгиб тем больше, чем ближе к периферии расположены ткани с высоким сопротивлением изгибающей силе (так называемый модуль эластичности изгибания) и чем лучше эти ткани связаны друг с другом (конструкция связи). В то же время эволюция направлена к оптимизации функционирования при максимальной экономии. Вследствие обоих факторов для осевых органов оптимальна конструкция полого цилиндра. Она реализована главным образом в соломинах злаков, которые по соотношению длины к диаметру (до 500 1) относятся к наиболее замечательным природным конструкциям. Впрочем, полые цилиндрические оси свойственны только относительно низкорослым, травянистым растениям, поскольку существует опасность их надлома (у злаков она исключается благодаря массивным узлам) и невозможно интенсивное ветвление (соломины злаков — кроме соцветий — большей частью неветвящиеся). Кроме того, была бы мала способность гасить напряжения, которая прежде всего важна для деревьев и для которой имеет значение центральная часть оси. Далее было обнаружено, что в периферических областях стволов деревьев ткани пребывают в состоянии напряжения растяжения, которое компенсировано напряжением сжатия в центре ствола. Тем самым меньшая прочность древесины на сжатие уравнивается прочностью ее на растяжение — что лишь в ограниченной мере было бы возможно в полых цилиндрических стволах. У лиан, старым стеблям которых, очевидно, не нужно быть устойчивыми к изгибам, а, наоборот, следует быть гибкими, часть препятствующей изгибанию ткани «возвращается» на периферию стебля в ходе его вторичного утолщения. У корней, которым требуется прочность не на изгиб, а на растяжение, реализована с самого начала конструкция каната1, т.е. все механические элементы сосредоточены в центральном цилиндре, который окружен паренхимой (см. рис. 4.77).
1 Конструкцию каната имеют стебли лиан, в которых отдельные тяжи жестких тканей пронизывают массив мягкой ткани. Располагающиеся в толще грунта корни не испытывают напряжений растяжения. — Примеч. ред.
Рис. 3.21. Целесообразное расположение механических элементов (F, G — пo W Rasdorski, H — L — по T Speck)
A, B—напряжения в балке при ее прогибании растяжение выпуклой стороны, сжатие с вогнутой, «нейтральная линия» n хотя и прогибается, но не изменяет длины Повышенная прочность на изгиб достигается прежде всего укреплением выпуклой и вогнутой наружных сторон, С—Е — схематичные поперечные срезы осевых органов с различно расположенными механическими тканями (черный) при равной их удельной площади (11,1% общей площади поперечного среза) С — центральное расположение, например центральный цилиндр корня, D—срединное расположение, например склеренхимное кольцо с проводящими пучками в стеблях настоящих двудольных, Е—периферическое расположение, например соломина злаков, так как механические ткани примерно в 100 раз тверже, чем паренхимная ткань, то прочность на изгиб при равных затратах материала находится в соотношении 1 2,5 8, F—конструкция индустриальной дымовой трубы, позволяющая экономить материалы, труба укреплена всего 16 стальными тягами, G—для сравнения — поперечный срез стебля пухоноса Trichophorum caespitosum, H, I—черная ольха (Ainus glutinosa), однолетний стебель (Н, диаметром 4 мм) с широкими сердцевиной и корой (заточкована) легко изгибаем, многолетняя ветвь (I, диаметром 37 мм) из-за сильно увеличенной древесинной части существенно прочнее на изгиб, К, L—у лиан (для примера здесь Aristolochia macrophyfla) можно наблюдать противоположную тенденцию развития молодые оси (К, однолетний, самоподдерживающийся «ищущий побег» диаметром 5 мм) прочны на изгиб благодаря периферической колленхиме и расположенному под ней замкнутому кольцу склеренхимы (черное), в то время как более старые оси (L, 14-летняя, диаметром 30 мм) гибки вследствие фрагментированного расположения периферических механических тканей и образования мягкой древесины с широкими лучами и широкопросветными сосудами (ср рис 4 49, С)
Реализованные в эволюции организмов решения проблем предполагают использовать и в технике (бионика). Действительно, на этом пути можно получить важные технические решения. Однако приходится принять во внимание, что биологические конструкции в целом оптимизированы по структуре, а технические, напротив, оптимизированы по материалу.
3.2.4. Проводящие ткани
Чтобы переместить в организме растворенные вещества на клеточные, микроскопические, расстояния достаточно диффузии, вызванной термическим движением частиц в растворе. Однако эффективность диффузии уменьшается пропорционально квадрату расстояния (см. 6.3.1.1 и 6.3.1.2). Уже внутри таких особенно крупных клеток, как корневые волоски, клетки междоузлий харовых (см. рис. 5.9; 11.106, А) и т.п., одной диффузии недостаточно, ее дополняет движение цитоплазмы. Наконец, у еще более крупных многоклеточных организмов, как растений, так и животных, развиваются особые проводящие системы, в которых поддерживаются направленные потоки веществ. В то время как у животных потоки идут в межклеточных пространствах (полости тела, кровеносные сосуды), у высших растений образуются особые клетки, в которых текут жидкости. Эти очень глубоко и терминально дифференцированные (т.е. не способные более к какому-либо дальнейшему развитию) клетки объединены в проводящие пучки. Проводящие пучки в листьях можно видеть невооруженным глазом как «нервы», или «жилки». В корнях проводящие ткани собраны в единый центральный цилиндр.
Во всех проводящих системах существуют две различные ткани. В лубе (флоэме, лептоме) (от греч. рhloios — луб, кора; leptos — тонкий, мягкий) органические соединения транспортируются по живым, хотя и безъядерным, клеткам с тонкими, неодревесневшими оболочками. В древесине (ксилема, гадром) (от греч. хуlon — древесина; hadros — твердый, жесткий) вода с растворенными минеральными питательными веществами от зон всасывания корней перемещается по отмершим, полым трубковидным клеткам или трубкам из многих клеток с крепкими, одревесневшими оболочками в листья, где она выделяется в ходе транспирации или гуттации (транспирационный ток, см. 6.3.5). Как во флоэме, так и в ксилеме клетки прозенхимные и ориентированы в проводящих пучках продольно. При этом продольно связанные ряды клеток составляют проводящие пути.
3.2.4.1. Флоэма
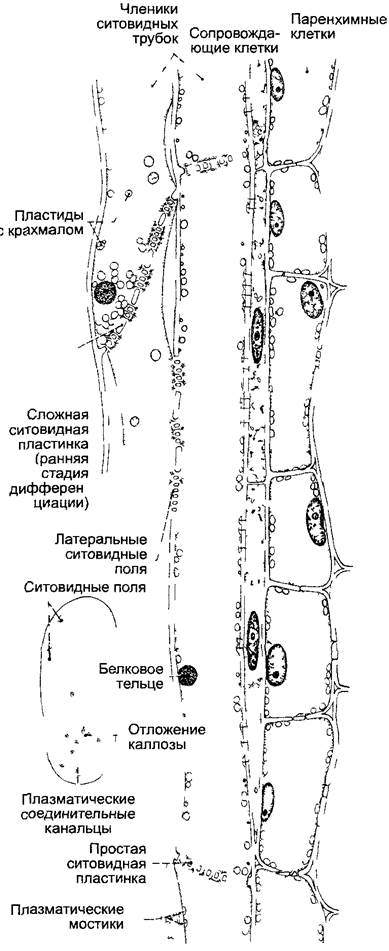
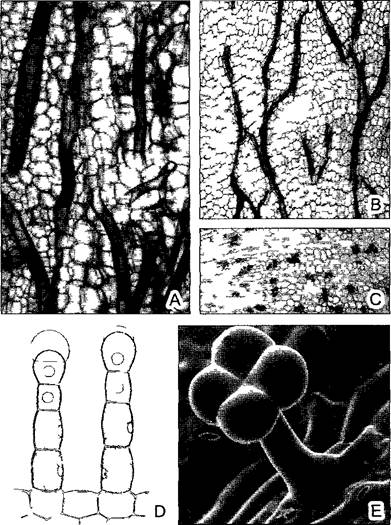
На рис. 3.22 и 3.23 показаны разные формы проводящих элементов флоэмы. Эволюционно исходны и низко эффективны ситовидные клетки. Они узкопросвет- ны и соединяются сильно скошенными конечными стенками1 с соседними ситовидными клетками клеточного ряда. Эти стенки (а также боковые, контактирующие с другими ситовидными клетками) пронизаны увеличенными плазмодесмами, которые здесь обозначают как ситовидные каналы 2. Каналы располагаются группами — ситовидными полями, облик которых послужил основой их названия У многих покрытосеменных такая примитивная проводящая система развилась в систему непрерывных ситовидных трубок из продольно вытянутых клеток большего диаметра с несущими ситовидные поля косыми или поперечными стенками — члеников ситовидных трубок. В наиболее высокоразвитых типах флоэмы, присущей вьющимся и лазающим растениям, поперечные конечные стенки члеников представляют собой простые ситовидные пластинки с особенно крупными ситовидными каналами.
Рис. 3.22. Элементы флоэмы (А—D — по W Zimmermann, Е—по Н Fitting, F—L — по А Resch)
Эволюционно исходны прозенхимные клетки без какой-либо особенной структуры клеточных стенок (например, Rhynia, А) У плауновидных возникли примитивные ситовидные поля (В), в дальнейшем филогенезе образовались ситовидные клетки с ситовидными каналами (например, пасленовые, С) и, наконец, ситовидные пластинки с ситовидными каналами (например, у тыквенных, D) (У всех покрытосеменных растений есть ситовидные трубки с простыми (у тыквенных) или сложными (пасленовые) ситовидными пластинками, а не ситовидные клетки Ситовидные клетки свойственны голосеменным и сосудистым споровым растениям, включая, вероятно, и Rhynia — Примеч. ред. ) Е—Cucurbita реро, поперечный срез ситовидной трубки с ситовидной пластинкой и сопровождающей клеткой g (бООх) (Ситовидная пластинка и сопровождающая клетка располагаются на разных уровнях и не могут попасть на один срез — Примеч. ред. ) F—L — развитие членика ситовидной трубки и сопровождающей клетки у бобов Vicia faba (F—неравное деление клетки, I—L — растворение ядра и тонопласта в членике ситовидной трубки)
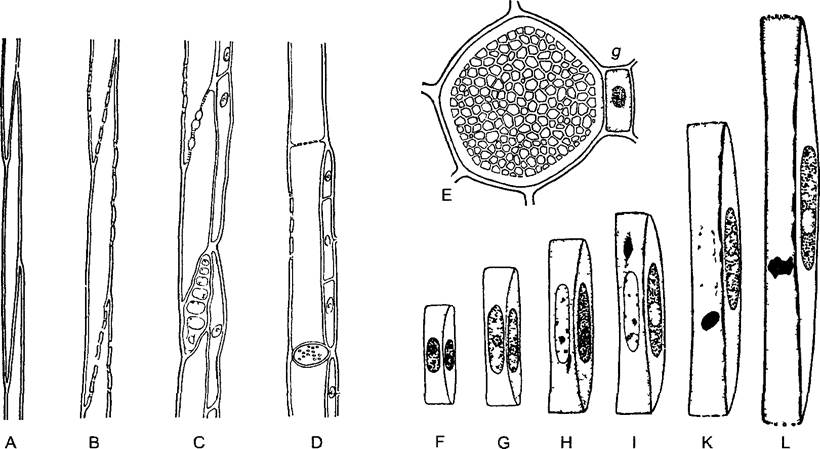
Рис. 3.23. Членик ситовидной трубки с сопровождающими клетками и флоэмной паренхимой у Passiflora coerulea (по R Kolimann)
Слева сложная ситовидная пластинка с пятью ситовидными полями (750х)
1 Ситовидные клетки имеют веретеновидную или уплощенно-веретеновидную форму, и конечных стенок у них нет — Примеч. ред.
2 Ситовидный канал представляет собой не просто крупную плазмодесму, а результат закономерного преобразования исходной плазмодесмы, включающий сложную реорганизацию прилегающей части клеточной стенки — Примеч. ред.
Ситовидные клетки и членики ситовидных трубок имеют живые протопласты с немногими митохондриями и пластидами, содержащими крахмал или белки (см рис 11 167). Клеточное ядро и тонопласт, диктиосомы и рибосомы рано растворяются, цитоплазма и клеточный сок смешиваются (одно из немногих исключений из правила компартментализации, см бокс 2 3). Эндоплазматический ретикулум превращается в так называемый «ситовидный ретикулум», состоящий из разветвленных трубочек и гладких, складчатых цистерн. Характерным компонентом зрелых сито видных элементов являются филаменты или трубочки из Ф-белка (флоэмный белок). Будучи безъядерными, нежными клетками, ситовидные элементы живут недолго, большей частью они деградируют к концу вегетационного периода и у многолетних растений заменяются новыми. У многолетних однодольных, например, у пальм, они могут оставаться живыми годами. Как и плазмодесмы, ситовидные каналы в период покоя закрываются каллозой. В перерезанных или поврежденных ситовидных трубках настоящих двудольных и некоторых однодольных находят ситовидные каналы, закупоренные пробками из Ф-белка или фрагментов топнувших пластид.
У покрытосеменных к каждому членику ситовидной трубки примыкает одна (редко больше) более мелкая, содержащая ядро и много митохондрий сопровождающая клетка(и) (см рис 3 23). Эти напоминающие железистые клетки связаны с члениками ситовидных трубок многочисленными плазмодесмами. Они обеспечивают обмен веществ безъядерных проводящих элементов. Например, Ф-белок (как образующий филаменты ФФ1, 80 — 120 кДа, так и его более мелкий сопровождающий протеин ФФ2) синтезируется в сопровождающих клетках и немедленно передается в членики ситовидных трубок, где образуются сначала высокоупорядоченные белковые тельца, которые позже распадаются на отдельные филаменты. Вторая важная функция сопровождающих клеток — это контролируемая загрузка и разгрузка ситовидных трубок. Это также проявляется в особенных структурных признаках или у них образуется особенно много плазмодесм с соседними клетками (внутриплазматический = симпластический тип, преимущественно у субтропических и тропических растений) или поверхности клеток увеличиваются за счет лабиринта клеточной оболочки (см. рис. 3.27; внеплазматический = апопластический тип, у растений умеренных и холодных зон).
У покрытосеменных комплекс из членика ситовидной трубки и сопровождающих клеток возникает из единой материнской клетки путем ее неравного деления. Ситовидные клетки голосеменных и папоротников не имеют сопровождающих клеток. Однако у этих растений есть богатые белками паренхимные клетки, которые также тесно связаны с ситовидными клетками, как сопровождающие клетки с члениками ситовидных трубок, хотя они в противоположность последним возникают из разных материнских клеток. Эти клетки называют белковыми клетками, или клетками Страсбургера.
3.2.4.2. Ксилема
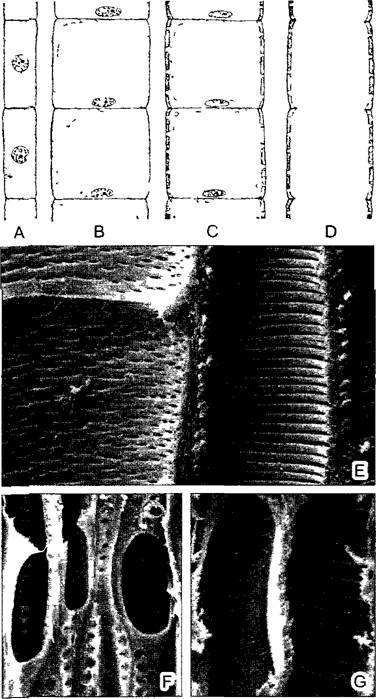
Транспирационный ток (см. 6.3.5) движется по трубковидным клеткам, протопласты которых по достижении функциональной зрелости отмирают и исчезают вследствие саморастворения (автолиза) — выразительный пример запрограммированной смерти клеток (апоптоз) у растений. Остаются только одревесневшие, пронизанные окаймленными порами клеточные оболочки. Есть два типа водопроводящих «трахеальных» элементов: трахеиды и сосуды. Трахеиды — это вытянутые в длину, узкопросветные отдельные клетки с очень косыми конечными стенками, пронизанными множеством пор для сообщения с соседними в продольном направлении трахеидами1 (см. рис. 2.75, С—F). Тяжи трахеид оказывают относительно высокое сопротивление потоку раствора. Существенно меньше оно в широкопросветных, укороченных члениках сосудов, у которых конечные стенки сильно перфорированы или вообще лизированы (рис. 3.24). Более крупный диаметр сосудов (от 60 до более700 мкм) — они чаще всего различимы невооруженным глазом как «поры древесины» — связан с тем, что молодые членики сосудов вследствие полиплоидизации своих ядер (8—16 n) растут в ширину, прежде чем их клеточные оболочки потеряют способность к росту из-за формирования утолщенных вторичных оболочек.
1 Трахеиды имеют веретеновидную форму, и у них нет конечных стенок. — Примеч. ред.
Рис. 3.24. Сосуды (A—D — по Е W Sinnott; Е—СЭМ-микрография W. Barthlott; G—СЭМ- микрография S G.Sombert):
A—D — развитие многочленного сосуда из одного ряда клеток путем их увеличения (полиплоидизация, вакуолизация), образования одревесневших утолщений оболочек, растворения поперечных стенок и отмирания протопластов (150х); Е—по типу утолщений оболочки различают точечные (слева) и спиральные (справа) сосуды, продольный разрез через проводящий пучок тыквы (360х); F—широкопросветные поры между члениками сосудов; пористые сосуды в древесине бузины Sambucus nigra (500х); G—лестничная перфорационная пластинка в точечном сосуде древесины березы (1300х)
Лигнификация оболочек трахеид и члеников сосудов предотвращает коллапс этих трубчатых клеток, в которых при интенсивной транспирации развивается отрицательное давление. Поэтому при перерезании побегов в сосуды засасывается воздух. Так как спиральные сосуды, кроме того, еще и структурно похожи на проводящие воздух трахеи насекомых, для них раньше использовали вводящее в заблуждение название «трахеи» (М. Malpighi, 1628—1694, один из основоположников анатомии растений) (от греч. trachelos — воздушные трубки).
Особенно простые водопроводящие элементы встречаются у лиственных мхов; их стебли содержат центральный тяж из продольно вытянутых клеток без содержимого с утолщенными оболочками (гидроидов). У папоротников и голосеменных преобладают трахеиды, диаметр которых больше, а сопротивление конечных стенок потоку уменьшено из-за их косого расположения и наличия пор. Разделение функций проведения и опоры произошло в эволюции позже. У голосеменных еще преимущественно трахеиды слагают несущий ствол. Сосуды в эволюции возникали многократно и независимо в разных таксонах. Они встречаются уже у некоторых папоротников и голосеменных, но особенно широко представлены у покрытосеменных. У сосудов осталась только функция проведения, а функцию опоры осуществляет собственно механическая ткань из древесинных волокон (волокон либриформа). Конечно, и в древесине покрытосеменных, например, у лиственных деревьев наряду с сосудами еще встречаются трахеиды, а в развитии проводящего пучка в общих чертах повторяется эволюция водопроводящих элементов. Очень совершенные сосуды имеются у лиан. Они очень большого диаметра, и на протяжении до 10 м по длине стебля все поперечные стенки лизированы, в то время как обычно отдельные поперечные стенки остаются на расстоянии от нескольких сантиметров до 1 м, вероятно, для снижения опасности массовой эмболии. Особо высокая проводимость воды проводящими тканями лиан объясняется телеономически тем, что они как лазающие растения не образуют прямостоячий ствол (они нуждаются в растении-опоре, скалах или стенах), но по своим тонким стеблям все-таки должны обеспечить водой крону, объем которой сопоставим с объемом кроны дерева.
3.2.4.3. Проводящий пучок
В корнях, стеблях и листьях проводящие ткани располагаются в виде проводящих пучков (от лат. fasciculi — пучочки). При этом собственно проводящая ткань часто снаружи укреплена тяжами склеренхимных волокон и окружена эндодермой. В свою очередь, проводящие пучки в стеблях и листьях образуют сетчатую структуру, в то время как каждый корень имеет в центральном цилиндре единственный радиальный проводящий пучок, который собственно представляет собой комплекс пучков. По расположению флоэмы и ксилемы можно различить концентрические и коллатеральные пучки (рис. 3.25). Концентрические пучки с внутренней ксилемой присущи папоротникам, а с внешней ксилемой — подземным побегам однодольных. Намного чаще встречаются коллатеральные пучки (хвощи, голо- и покрытосеменные) (рис. 3.26; лат. соllateralis — бок о бок). При этом в осях древесинная часть всегда располагается внутри, в горизонтальных листьях — сверху. Особый тип — биколлатеральный проводящий пучок с двумя тяжами флоэмы; подобные пучки встречаются, например, у пасленовых и тыквенных. Если древесина и луб непосредственно граничат друг с другом, то говорят о закрытом проводящем пучке (рис. 3.26, А). Он состоит целиком из постоянных тканей. Этот тип пучка характерен для однодольных, что имеет важные следствия для роста этих растений (см. бокс 4.3). Напротив, большинство пучков голосеменных и покрытосеменных открытые, т. е. между флоэмой и ксилемой находится слой меристемы — пучковый камбий. На поперечном срезе он заметен благодаря регулярному расположению весьма тонкостенных клеток (рис. 3.26, В, С). Пучковый камбий играет решающую роль при вторичном утолщении стеблей.
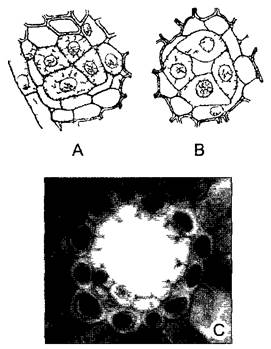
Рис. 3.25. Типы проводящих пучков
Расположение ксилемы (черная), флоэмы (крап) и камбия (белый) на поперечных срезах: А—концентрический проводящий пучок с внутренней ксилемой (амфикрибральный пучок), В—то же с внешней ксилемой (амфивазальный пучок); С—радиальный проводящий пучок с внутренней ксилемой и в данном случае — с четырьмя ксилемными участками (тетрархный пучок) развит в центральном цилиндре корней; на левой половине «закрытый» (однодольные), на правой половине «открытый» (магнолииды и настоящие двудольные); D—F —коллатеральный проводящий пучок: D—закрытый (однодольные); Е—открытый (большинство настоящих двудольных); F—биколлатеральный открытый (например, у тыквы)
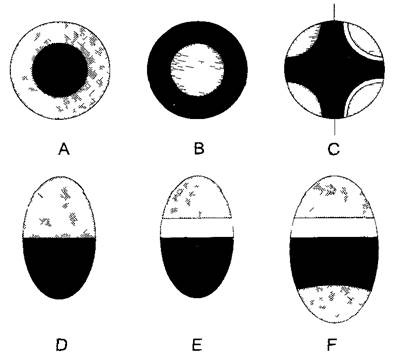
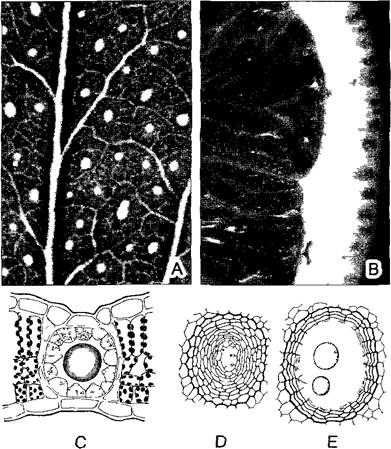
Рис. 3.26. Коллатеральный проводящий пучок (А, В — по D von Denffer, С — по К. Magdefrau).
А—поперечный срез закрытого коллатерального пучка кукурузы Zea mays, в протоксилеме кольчатый сосуд (*), разорванный при растяжении соседней паренхимы (ср рис 4.42); В—поперечный срез открытого коллатерального пучка лютика ползучего Ranunculus repens, С—трехмерное изображение открытого коллатерального пучка (все около 200х)
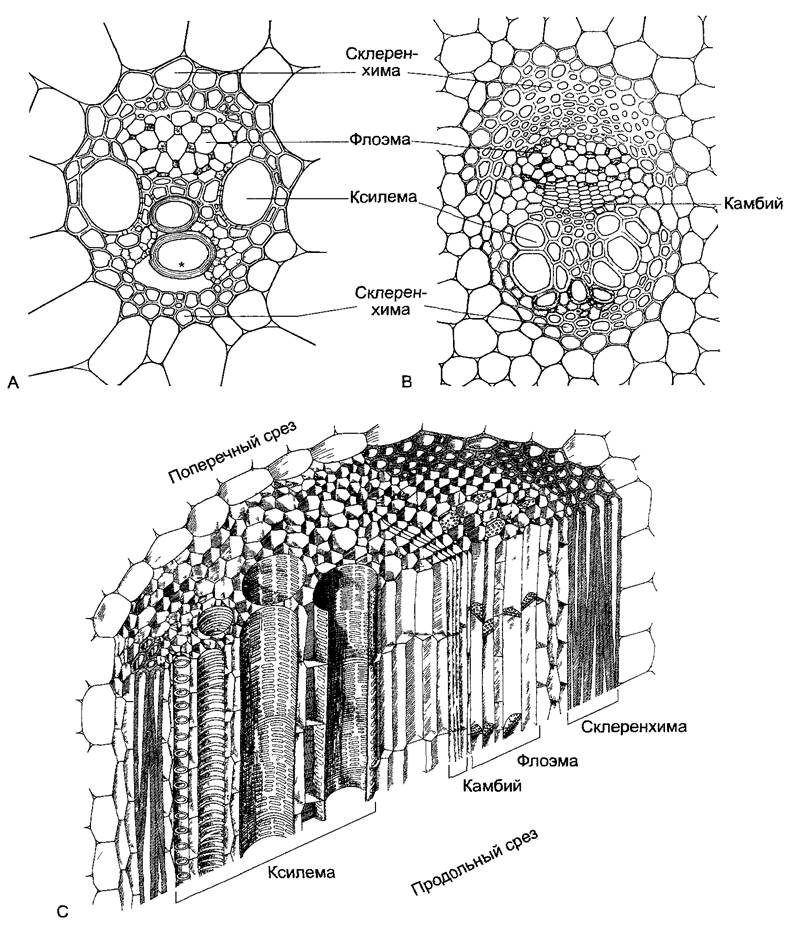
В коллатеральном пучке флоэма в целом развивается снаружи внутрь, ксилема — от внутренней границы пучка наружу, до его середины. Вследствие этого самые старые проводящие элементы ксилемы — сравнительно мало дифференцированные инициали и соответственно возникшая из них протоксилема —- расположены у внутреннего края древесины, а инициали луба и соответственно протофлоэма, — напротив, у внешнего края луба. Образованные позднее, полностью дифференцированные и функционально полноценные проводящие ткани метаксилемы и метафлоэмы расположены в середине пучка или соответственно рядом с пучковым камбием.
3.2.5. Железистые клетки и ткань
Железистые клетки образуют определенные вещества — секреты (лат. secernere — отделять, выбрасывать), которые выделяют наружу. В других случаях в качестве экскретоввыбрасываются шлаки обмена веществ, балластные или вредные вещества (см. 6.18). Вообще, секретами считают выделения, которые необходимы самому организму, в то время как экскреты принесли бы вред, если бы их не удалось удалить. Секреты образуются в цитоплазме железистых клеток, где обильно развиты эндоплазматический ретикулум и/ или аппарат Гольджи. Ядра железистых клеток сравнительно крупные. Напротив, вакуом выражен довольно слабо, если только он не служит местом накопления секрета/экскрета. Продукты железистых клеток часто аккумулируются в неплазматических пространствах, главным образом в вакуолях (внутриклеточная секреция/экскреция: «секреторные клетки», например, млечники и идиобласты с оксалатом; см. ниже). Однако чаще всего секрет/экскрет выделяется в анопласт, чему нередко способствует заметное увеличение поверхности (рис. 3.27). При этом возможно накопление секрета/экскрета внутри растения (во вместилищах секрета или смоляных ходах) или же выделение его в окружающую среду (пахучие вещества, нектар).
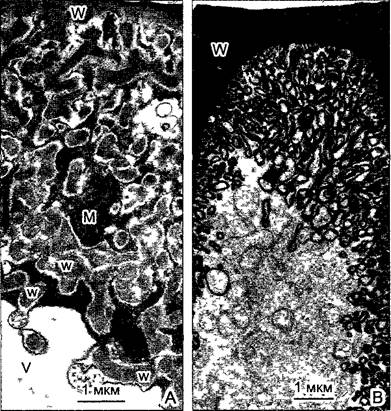
Рис. 3.27. Протуберанцы клеточной оболочки и апикальный лабиринт в клетках нектарников (электронные микрографии Е.Schnepf и Р. Christ).
Для многих железистых клеток типично увеличение поверхности в том месте, где происходит секреция веществ: А—лабиринт оболочки клетки нектарника на околоцветнике Gasteria; отходящие от апикальной клеточной стенки W, где выделяется секрет, многочисленные протуберанцы w почти достигают вакуоли V. Митохондрии М в лабиринте клеточной стенки поставляют энергию для активного транспорта секрета. Передаточные клетки (см. 6.18) сходным образом увеличивают поверхность клеточных мембран при помощи лабиринта клеточных стенок; В—апикальная область нектарника Asclepias curassavica с бесчисленными складками клеточной мембраны, которые выявлены при помощи специального контрастирования углеводов секрета
Железистые клетки у растений обычно встречаются поодиночке, реже несколько или много железистых клеток объединены в локализованную железистую ткань (см. рис. 3.28; 3.30). Крупных желез тела, сравнимых с железами животных, у растений нет. С другой стороны, функциональное многообразие желез растений отражает огромный масштаб вторичного обмена веществ. В конце концов все это — следствия открытой организации тела растения. Многообразие секретов/экскретов соответствует разнообразию функций, в которых они задействованы. Важнейшие примеры:
• Защита растения: многие секреты ядовиты (алкалоиды, стероидные гликозиды), горькие на вкус или действуют как аллергены. Рост грибов подавляют фенольные дубильные вещества и терпеноиды, у животных секреты растений вызывают отвращение или нарушают обмен веществ или развитие (см. 6.16; 9.4.1). С помощью млечного сока, камедей и смол, изливающихся при повреждениях, растения могут дезинфицировать и быстро затягивать раны.
• Привлечение животных: эфирные масла и тому подобные пахучие вещества, часто образуемые в железистой ткани особых осмофоров, играют роль в опылении и распространении семян. Нектарные железки (нектарники) вознаграждают животных, которые необходимы растению. Большей частью нектарники локализованы в цветках, но имеются и внецветковые (экстрафлоральные) нектарники. Их сахаросодержащим секретом питаются, например, насекомые, которые (как муравьи или термиты) являются биологическими врагами насекомых-вредителей. Некоторые насекомоядные растения (см. бокс 4.4, а также 9.1.2) привлекают жертв и ловят их при помощи блестящих выделений клейкой слизи и, наконец, химически переваривают их выделениями пищеварительных железок с последующим усвоением путем осмотического всасывания веществ (см. рис. 3.31).
✵ Специализированные выделительные клетки или выделительная ткань служат для экскреции. Наиболее известный пример — клетки с оксалатами, которые, связывая кальций, выводят его из обмена веществ и накапливают в своих вакуолях в виде кристаллов оксалата кальция1 (см. рис. 2.61). Растения засоленных местообитаний, например, морских побережий, имеют (подобно морским птицам) соляные железки для активного выделения связанных солей наружу.
1 Главная функция этих клеток — выведение из метаболизма ядовитого оксалата, а не Са. — Примеч. ред.
✵ Пограничный случай между железками и нежелезками представляют структуры, осуществляющие интенсивный дальний транспорт собственных веществ. Клетки этого типа часто находятся в пограничной ткани: пропускные клетки эндодермы (см. рис. 3.18, В; 4.78, В), передаточные клетки обкладки пучка, лежащая под гидатодами бедная хлорофиллом «эпитема»; сходную функцию имеют сопровождающие клетки флоэмы покрытосеменных. Эти клетки часто вырабатывают вещества (которые выделяют однонаправленно, т.е. только в одну сторону) не самостоятельно, чем они отличаются от типичных железистых клеток (однако сравните также вышеупомянутые солевые железки). Однако в цитологическом отношении они обладают признаками железистых клеток (крупные ядра, плотная плазма, а также увеличенная посредством выростов оболочки поверхность).
Ряд избранных примеров, приведенных ниже, должен дать представление о разнообразной структуре и функциях железок у растений.
3.2.5.1. Млечники
Некоторые растения при повреждении выделяют млечный сок. Общеизвестные примеры — виды молочая, одуванчик, фикус, чистотел, мак. Млечный сок соответствует клеточному соку или тонкоструйной плазме обильно разветвленной системы трубочек в теле растения. Эта система состоит из типичных выделительных клеток. Их необыкновенные размеры частично обусловлены многоядерностью (полиэнергидностью) гигантских клеток, которые пронизывают паренхиму как
нечленистые млечники (см рис 3 28, А). Подобные плазмодиальные млечники — они могут быть несколько метров длиной и вообще относятся к самым крупным клеткам — встречаются у многих молочаев, олеандра и фикуса (Ficus elástico) Членистые млечники — это, напротив, синцитии, они возникают путем слияния клеток после растворения первоначально существовавших поперечных стенок Млечники этого типа распространены у маковых (чистотел, Chehdonium, с желтым млечным соком, мак снотворный, Papaversomniferum, как источник опиума, содержащий морфин смеси алкалоидов), у сложноцветных с язычковыми цветками (Taraxacum, Scorzonera, рис 3 28, В, С, латук, Lactuca из-за них получил свое название от лат lac — молоко) и многих молочайных (например, гевея, Hevea brasiliensis)
Рис. 3.28. Железистая ткань и железистые волоски (D—по D von Denffer Е—СЭМ-микрография W Barthiott)
А—нечленистые млечники в паренхиме первичной коры молочая (Euphorbia) (50х), В, С—членистые млечники козельца Scorzonera purpurea на продольном и поперечном срезах корня (25х) D—железистые волоски черешка листа первоцвета Primula obconica, секрет скопившийся между клеточными стенками и кутикулой может быть причиной зудящей экземы (80х) Е—железистый волосок Uncanna из семейства Pedaliaceae с четырехклеточной головкой (250х)
3.2.5.2. Смоляные ходы и вместилища выделений
В то время как млечный сок находится в млечниках, в схизогенных межклеточных полостях накапливается вязкая смесь терпеноидов (эфирных масел), называемая смолой, или бальзамом (рис 3.29). Эти смоляные ходы (каналы) выстланы железистым эпителием. Как и млечники, смоляные ходы представляют со бой разветвленную систему сильно вытянутых трубочек, из которой при ранениях вытекает смола/ На воздухе она застывает и дезинфицирует, и закрывает рану.
Рис. 3.29. Смоляные ходы (А, В — по W Н Brown)
А В—схизогенное возникновение смоляного хода с крупноядерным железистым эпителием в древесине сосны (250х) С—смоляной ход в хвоинке сосны, железистый эпителий отграничен от мезофилла обкладкой (все 250х)
Смоляные ходы распространены главным образом у хвойных. Смолу некоторых видов применяют в промышленности (терпентин и соответственно терпентиновое масло, канадский бальзам). Янтарь представляет собой окаменевшую смолу. У покрытосеменных смола встречается редко.
Эфирные масла образуются у большинства высших растений, однако часто их количество так мало или летучие секреты так быстро выделяются наружу, что никаких особых скоплений не образуется Так, многие листочки околоцветников в клетках эпидермы или мезофилла содержат капельки эфирных масел, которые при соответствующей температуре испаряются и улетучиваются наружу (запах цветков роз, фиалок, жасмина) Но у многих видов жидкие эфирные масла накапливаются в схизогенных или лизигенных вместилищах (рис 3.30) Примеры — виды зверобоя (Hypericum) и эвкалипта (Eucalyptus) (схизогенные вместилища), а также лизигенные вместилища в кожуре плодов цитрусов (Citrus).
Рис. 3.30. Вместилище масел (В—по G На- berlandt, D, Е—по A Tschirch)
А, С—схизогенные вместилища масел у Hypericum perforatum, внешний вид (А, 2х) и поперечный срез листа (С, 50х), В, D, Е—вместилища масел в наружном слое кожуры плода апельсина (В, 2х) и лизигенное их образование у Citrus limon (D, Е, 25х)
3.2.5.3. Головчатые волоски и железистые эмергенцы
На концах волосков и эмергенцев часто сидят железистые клетки или их группы (см рис 3 28, D, Е, 3 14, D) Так как железистые клетки (или же железистая ткань) округлые и толще, чем ножка волоска/эмергенца, возникает впечатление головок на тонких шейках, чем и обусловлено их название. У «сидячих» железок ножки нет. Секрет (обычно эфирное масло) скапливается между клеточной стенкой и кутикулой и вследствие своей липофильности может испаряться сквозь отстоящую кутикулу. В других случаях кутикула разрывается и высвобождает гидрофильный секрет (рис 3.31) (содержащая полисахариды ловчая слизь у росянки, Drosera), протеинсодержащие секреты пищеварительных железок у насекомоядных растений).
Рис. 3.31. Скопления ловчей слизи на головках железистых эмергенцев у насекомоядного растения росянки Drosera cuneifolia (см. бокс 4 4)
Билатерально симметричный дорсовентральный лист сверху (А) и сбоку (В) (2,5х)