БОТАНИКА ТОМ 1 - КЛЕТОЧНАЯ БИОЛОГИЯ. АНАТОМИЯ. МОРФОЛОГИЯ - 2007
3. ТКАНИ ПОБЕГОВЫХ РАСТЕНИЙ
Комплекс сходных клеток обозначают понятием ткань. Сходство прежде всего проявляется во внешнем облике клеток, но также и в их функционировании, поскольку существует общее соответствие структуры и функции. Таким образом, ткани можно охарактеризовать также и по их роли в организме, однако в первую очередь ткани — все-таки морфологические образования. Надклеточные функциональные образования обозначают как органы; часто они построены из нескольких тканей. Изучение тканей — предмет гистологии (от греч. histos — ткань).
Первая грубая морфологическая классификация составляющих ткани клеток основана на их форме. Более или менее изодиаметрические клетки и состоящие из них ткани обозначают как паренхимные, а продольно-вытянутые клетки и волокнистые ткани — как прозенхимные. Свойства паренхимных тканей одинаковы во всех направлениях (изотропия). Для прозенхимных тканей характерно, например, по механической прочности, преобладающее направление, а именно продольное, параллельное их параллельно расположенным клеткам (анизотропия). Наряду с этими двумя основными формами — объемно-многоугольной и нитевидно-волокнистой — имеется еще плоская пластинчатая, которая присуща преимущественно покровным тканям (форма эпидермальных клеток).
Уклоняющиеся по внешнему виду и функции клетки, располагающиеся среди гомогенной ткани, обозначают как идиобласты (см. рис. 3.9).
Чем богаче тканевой состав организма, тем выше достигнутая им степень дифференцировки, т.е. способность разделения функций его клеточным составом. Высота организацииорганизма соответствует числу типов его клеток и тканей. На этом основано крупное разделение царств растений и грибов. Эволюционное развитие царства растений в целом проходило от наиболее простых к все более высоко организованным, а часто и более крупным формам.
Многие водоросли достигли низкой степени дифференциации. В простейшем случае любые клетки их тела могут выполнять все жизненные функции, включая размножение. У более сложно устроенных водорослей и мхов можно уже различить несколько разных тканей. Большего многообразия тканей среди растений достигают побеговые формы1. Поэтому именно на примере последних рассмотрены ткани растений в данной главе (в главе 5 описаны сильно уклоняющиеся формы тканей водорослей и грибов).
1 Побеговую организацию имеют все мхи и многие печеночники, а также все сосудистые растения. Противопоставлять последние мхам — ошибка. — Примеч. ред.
Для высокоорганизованных и эволюционно высокоразвитых исторически наиболее молодых побеговых растений — семенных растений (Spermatophyta) — характерно четкое разделение образовательных (меристем) и постоянных тканей. Функция меристемы (от греч. merizein — делить) состоит в продуцировании клеток тела (от греч. soma — тело). Клетки постоянных тканей, напротив, не способны к делению и специализированы на выполнении определенных функций. Меристематические клетки непрерывно проходят клеточный цикл2 (см. 2.2.3.5; 7,3.2), а клетки постоянных тканей, напротив, в норме задерживаются в фазе G1(Go-фаза). Кроме того, меристематические клетки кончиков побега и корня мелкие, тонкостенные и без центральной вакуоли. Клетки постоянных тканей много крупнее. Часто они даже заметны невооруженным глазом; их объем может более чем в 1 000 раз превышать объем эмбриональных клеток. В них сформирована центральная вакуоль, а стенки значительно толще. В то время как меристематические клетки растут путем увеличения сухого вещества (эмбриональный, или плазматический, рост), рост клетки при переходе в дефинитивное состояние обусловлен увеличением вакуоли (постэмбриональный рост, или растяжение; ср. 7.1.1). Таким образом, эмбриональным клеткам верхушечной меристемы побега и корня (апикальная ме ристема = эвмеристема) и их непосредственным производным в первичной меристеме растяжение еще предстоит, а клетки постоянных тканей, напротив, уже прошли его. Растяжение типично для растительной клетки, а у животных ничего подобного нет. Так как фазы постэмбрионального роста, в общем, сменяются быстро, то растения («растущие») при одинаковом потреблении энергии могут расти быстрее, чем животные.
2 Инициал и меристем обычно делятся через длительные промежутки времени. — Примеч. ред.
При регенерации, а также в ходе нормального развития клетки постоянных тканей могут реэмбрионизироваться и образовать вторичные (производные) меристемы.Реэмбрионизацию раньше обозначали как дедифференцирование. Но это выражение неудачное, потому что и меристематическое состояние может быть результатом дифференцировки — каждая реэмбрионизация четко показывает это.
3.1. Образовательная ткань (меристема)
Оплодотворенная яйцеклетка (зигота) высших растений развивается сначала в зародыш (эмбрио) (рис. 3.1; ср. также 7.4.1 и рис. 7.27). Уже при первом, обычно неравном делении зиготы устанавливается будущая ось полярности: из производных более мелких и оптически более плотных апикальных клеток позже образуется побег, а из более крупных базальных клеток — первичный корень. Но еще раньше базально возникает подвесок (суспензор), через который растущий зародыш связан с материнским растением (от лат. suspendere — вешать) и может потреблять запасы питательной ткани семени, эндосперма.
Как только зародыш станет крупнее, деления клеток сосредоточатся на кончиках (апексах) побега и корня (лат. арех — кончик, верхушка). Таким образом, побеги и корни проявляют верхушечный рост, а клетки, из которых они состоят, есть производные их апикальных меристем. Боковые побеги и корни обладают собственными апикальными меристемами. Непосредственные производные апикальной меристемы часто делятся особенно активно. Они еще сохраняют меристематический характер, но часто по их расположению и внешнему виду можно прогнозировать дальнейшую судьбу их производных. Эту верхушечную образоватетьную ткань обозначают как первичную меристему и различают в ней протодерму, из которой позже образуется покровная ткань (эпидерма), основную меристему как поставщика основной ткани (паренхимы) и прокамбий, который даст проводящие ткани.
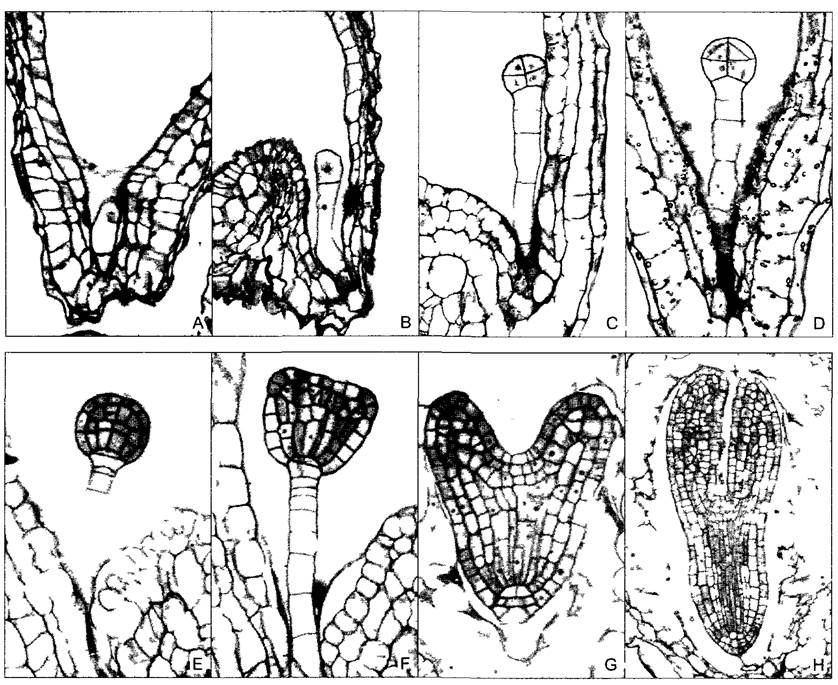
Рис. 3.1. Развитие зародыша Arabidopsis thahana (по U Mayer и G Jurgens)
А—зигота, В—стадия двух клеток после асимметричного деления зиготы, С—«октант», из верхней половины восьмиклеточной шаровидной структуры позже образуются меристема побега и семядоли, из нижней — гипокотиль и основная часть зародышевого корня, D—стадия дерматогена по периферии вычленились предшественники эпидермальных клеток, Е—стадия глобулы, к ряду клеток подвеска (суспензора) примыкает гипофиза, из которой разовьются центральная часть меристемы корня и корневой чехлик, F—стадия треугольника установление билатеральной симметрии, которая еще отчетливее выражена на стадии сердечка (G) и последующей стадии торпеды (Н) вследствие прогрессирующего развития обеих семядолей
По мере удаления от первичной меристемы ее производные превращаются в клетки постоянных тканей. Если в некотором участке, который уже перешел в состояние постоянной ткани, довольно крупные комплексы клеток остаются меристематическими, то их отличают от апикальной меристемы как остаточные меристемы. Особый случаи представляют так называемые интеркалярные меристемы, расположенные в стебле между уже дифференцированными участками и способные вызывать локальный продольный рост много ниже апекса. При наличии мелких групп активно делящихся клеток или одиночных клеток, которые в конце концов полностью переходят в постоянную ткань, говорят о меристемоидах.
Путем делений клеток апикальной и первичной меристем, а также последующих гисто- и морфогенетических процессов образуется первичная структура тела растения. У травянистых одно- или двулетних растений она соответствует и его конечному состоянию. Эти растения отмирают после семеношения, если не размножаются вегетативно ползучими побегами или иными способами. У многолетних древесных растений (кустарники, деревья), напротив, происходит вторичное утолщение, вследствие которого стебли превращаются в массивные одревесневшие стволы, боковые ветви — в одревесневшие сучья, а корни — в толстые образования, также состоящие преимущественно из древесины. На поверхности многолетних стволов, сучьев и корней образуется вторичная кора. Вторичное утолщение, при котором диаметр стебля в итоге может в 10 000 раз превысить первоначальный диаметр, обусловлено деятельностью латеральных меристем (производная меристема, или камбий). Это пластинчатая меристема, параллельная поверхности органа. Такие меристемы расположены не на верхушке побега или корня, как апикальные меристемы, а образуют боковой покров вокруг этих осей (от лат. lateralis — боковой). Есть два вида латеральных меристем: стеблевой, или соответственно корневой, камбий (называемый часто просто «камбий»), который формирует древесину и луб вторично утолщенных стеблей и корней, и пробковый камбий, или феллоген, образующий слои пробковой ткани, которые в свою очередь ведут к возникновению корки.
Все апикальные меристемы и камбии отличаются наличием инициалей — стволовых клеток. Эти клетки делятся неравно (см. рис. 4.43, А; 7.26, А, В): одна дочерняя клетка остается инициальной, в то время как другая превратится в клетку постоянной ткани. Поэтому в меристеме всегда остаются инициальные клетки, и тем самым сохраняется способность к продолжению роста и образованию новых органов. Наличие верхушечных меристем с инициалями характеризует растение как открытую систему, чем оно кардинально отличается от животных.
3.1.1. Апикальная (верхушечная) меристема и первичная меристема
Меристематические клетки побегов и корней изодиаметричные и мелкие (диаметр 10—20 мкм). Их стенки очень нежные и бедны целлюлозой. Все клетки примыкают друг к другу плотно, без межклетников. Клетка заполнена богатой рибосомами цитоплазмой и крупным ядром, расположенным в центре. Отсутствуют большая центральная вакуоль и запасные вещества, а пластиды представлены пропластидами.
У большинства высших растений комплекс апикальной и первичной меристем (точка роста) верхушек побега и корня имеет более или менее конусовидную форму (рис. 3.2; 3.3; 3.5: конус нарастания); однако на концах побегов он также может быть уплощенным или даже вогнутым, как у розеточных растений и в блюдцевидных крупных верхушках многих пальм.
Число клеток увеличивается непосредственно в первичной меристеме, однако деления клеток в ней происходят в течение ограниченного времени. При помощи микрорадиоавтографии удалось показать, что инициальные клетки в апикальной меристеме делятся относительно редко. В корнях кукурузы полный клеточный цикл инициальных клеток (центральные материнские клетки) длится более 7 дней, т. е. почти в 14 раз дольше, чем у их активно делящихся производных. У инициальных клеток, которые вакуолизированы сильнее остальных и обладают более мелкими и плотными ядрами, удлинена Gr фаза. Вследствие этого инициальный комплекс часто характеризуют как покоящийся центр (англ, quiescent centre).
Конусы нарастания побега и корня существенно различны. Конус нарастания побега сразу за апексом (рис. 3.2; 3.3) образует боковые выросты, которые разрастаются в листья или, при определенных условиях, в боковые побеги. При этом листья и боковые побеги берут начало от поверхностных клеточных слоев, сохраняющих меристематический характер, т.е. они экзогенны. Сначала листья растут быстрее, чем конус нарастания, они окутывают и защищают его. Однако в противоположность апексу их рост ограничен.
Рис. 3.2. Апекс побега хвоща (по Е. Strasburger): А—продольный срез; В—вид сверху (180х). Апикальная клетка отделяет косыми стенками р сегменты S, S". Затем сегменты делятся дальше дополнительными стенками m; f, f"—зачатки листьев; g—инициальные клетки боковых почек; I—боковая стенка сегмента
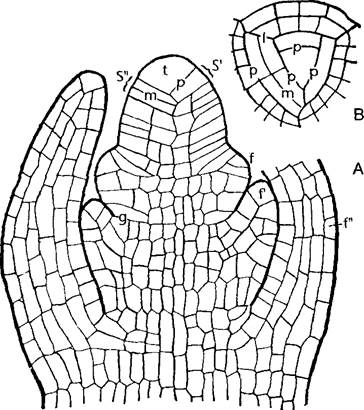
Рис. 3 3. Конус нарастания побега (А В—световая фотофафия Н Faik D—СЭМ микрография W Barthlott)
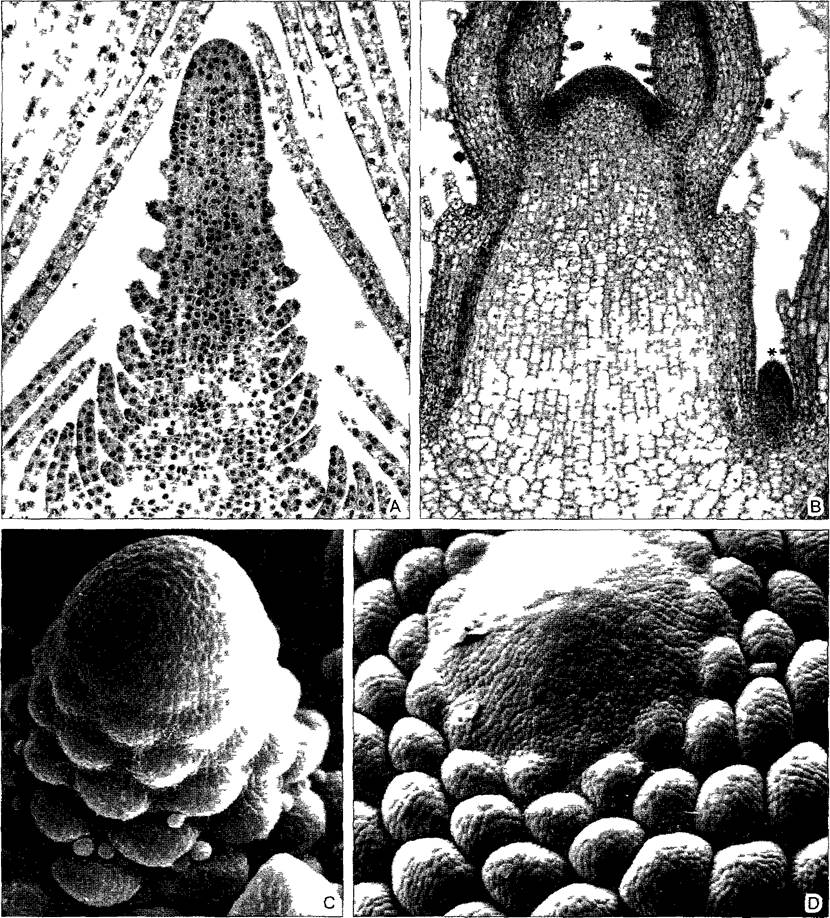
А—высококонусовидныи апекс Elodea densa с 2 слоями туники состоящие только из двух слоев клеток листья превышают апекс побега заметна большая разница между эмбриональными клетка ми в апексе и дифференцированными вакуолизированными клетками листьев (140х) В—апекс побега Coleus меристема терминального (*) и пазушных апексов выделяется оптической плотностью (отсутствие вакуолизации богатая рибосомами плазма крупные ядра) в обоих листьях самого молодого узла уже дифференцирован прокамбии и соответственно проводящая ткань протягивающа яся в ось (85х) С—высококонусовидныи апекс Hippuns vulgaris (мутовчатое листорасположение ср рис 4 12 А)(280х) D—апекс побега Picea abies (очередное листорасположение рис 4 12 D)(100x)
Корни не несут листьев, поэтому конус нарастания корня не защищен молодыми листьями, а укрыт корневым чехликом, образуемым непосредственно апикальной меристемой. Боковые корни возникают не экзо-, а эндогенно и первоначально растут наружу через первичную кору и покровную ткань. Зачаток бокового корня возникает не в апикальной области, а в уже прошедшей дифференцировку зоне. Это предполагает новообразование апикальной меристемы, в то время как меристемы боковых побегов и листовых зачатков образуются непосредственно из апикальной меристемы (фракционирование меристемы).
3.1.1.1. Конус нарастания побега
У многих морских водорослей, мхов и хвощей, а также у множества папоротников в апикальной меристеме одна, причем особенно крупная, инициаль — апикальная клетка. Она имеет форму тетраэдра, у которого выпуклое основание обращено наружу. От трех остальных сторон последовательно отделяются клетки (трехгранная апикальная клетка; см. рис. 3.2; 5.13). Возникающие при этом сегменты делятся далее, сначала очень регулярно. У папоротников листовые зачатки также начинают свое развитие с клиновидных двухгранных апикальных клеток.
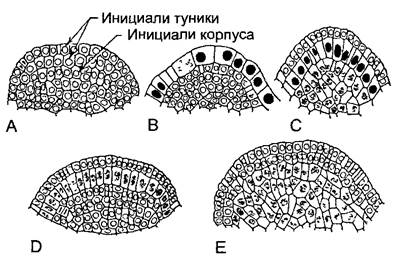
У высших папоротникообразных, особенно плауновых, и многих голосеменных апикальная клетка замещена группой равнозначных инициальных клеток, благодаря чему увеличено число стволовых клеток. В этом инициальном комплексе клетки могут делиться как антиклинально, так и периклинально (перпендикулярно и соответственно параллельно поверхности). У некоторых высокоразвитых голосеменных и всех покрытосеменных инициали расположены послойно. Только клетки самой внутренней группы делятся периклинально и антиклинально, образуя основную массу конуса нарастания — корпус. В слоях инициалей над ним клетки делятся только антиклинально. Эти слои клеток образуют тунику (от лат. hemd — кожа; рис. 3.3, А; 3.4; 3,5). Общее число слоев инициалей соответствует числу слоев туники плюс один.
Понятия туника и корпус чисто описательные, они никак не показывают дальнейшее развитие образующихся из них клеток. Концепция «туника —корпус» сменила более старую концепцию гистогенов, по которой уже в апикальной меристеме должна быть предопределена будущая судьба всех возникших клеток. Но исследования прежде всего мутантов показали, что в меристеме роль выпавших клеток могут принять другие клетки. Апикальная меристема побега (в противоположность четко разграниченной меристеме корня) проявляет себя как изменчивый, корректирующий повреждения структурный комплекс со значительным потенциалом регуляции, без жесткой детерминации последующей судьбы клеток.
Ближе всего по свойствам к гистогену самый наружный слой туники, который обозначают L1. Он превращается позднее в эпидерму и соответственно может быть обозначен как дерматоген. Число слоев туники варьирует (один — у многих голосеменных, однодольных и кактусов, два — у большинства настоящих двудольных, больше двух — например, у сложноцветных). Их число бывает разным у одного и того же вида и часто меняется в течение онтогенеза, например, при переходе к образованию цветков. Если туника многослойна, то каждый ее слой (снаружи внутрь L1, L2...) имеет собственные инициальные клетки. Впервые это удалось показать путем полиплоидизации дурмана колхицином. Полиплоидия может проявиться в любом из двух слоев туники или в ткани корпуса. Величина ядра и клетки скоррелирована со степенью плоидности, поэтому такие периклинальные химеры легко узнаваемы при микроскопировании (рис, 3.4). Таким же методом можно доказать, что каждый слой создают несколько инициальных клеток. Часто полиплоидия ограничена отдельным сектором стебля (секториальные химеры).
Рис. 3.4. Конус нарастания Datura (80х) (по Satina, Blakeslee и Avery).
А—нормальное диплоидное растение (n = 2); В—Е— периклинальные химеры, полученные обработкой колхицином; В—наружный слой туники (протодерма) - 8n; С— второй слой туники - 8n, корпус = 4n; D—второй слой туники = 4n; Е—корпус = 4n
Общая величина апикальной меристемы побега большей частью находится между 50 и 150 мкм. Исключительно большая апикальная меристема с диаметром около миллиметра обнаружена у саговников и в развивающихся корзинках подсолнечника.
В норме апикальная меристема побега (сокращенно SAM, от англ, shoot apical meristem) покрытосеменных растений устроена так, как показано на рис. 3.5. Центральный комплекс инициалей окружен особо активно делящимися клетками первичной меристемы, кольцеобразной фланговой меристемой и расположенной глубже сердцевинной (= медуллярной) меристемой. Самый наружный слой клеток фланговой меристемы функционирует как протодерма (дерматоген). Базипетально фланговая меристема выделяет периферическую меристему первичной коры и полый цилиндр (часто состоящий из обособленных продольных тяжей) из клеток, которые начинают вытягиваться вдоль оси. Этот прокамбий соответствует той части первичной меристемы, которая дольше других типов остаточной меристемы остается меристематической. Позже из него возникает кольцо проводящих пучков стебля и соответственно в него впадают зачатки проводящих пучков молодых листьев, которые впоследствии становятся пучками листовых следов. Медуллярная и коровая меристемы вместе составляют основную меристему.
Рис. 3.5. Конус нарастания побега семенного растения (продольный срез): 1—апикальная группа инициалей со стволовыми клетками (центральные материнские клетки, у голосеменных часто хорошо заметные как «покоящийся центр»). К боковой фланговой меристеме 2 относится поверхностный дерматоген 3; она здесь представлена протодермой, из которой произойдут стеблевая и листовая эпидермы, и субпротодермой, из которой путем антиклинальных делений возникнут зачатки листьев (7′—7"'). В примыкающей снизу гистогенетической зоне можно различить медуллярную меристему 4, тяжи прокамбия 5 и меристему первичной коры 6. Из прокамбиальных тяжей позже возникнут проводящие пучки. Границы между названными меристематическими зонами редко резкие, и исследования по биологии развития показывают, что они (например, при отсутствии делений) часто могут взаимно замещаться
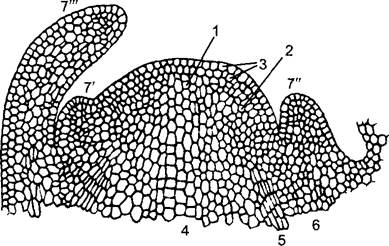
Но прежде чем эта дифференциация тканей станет четкой, на поверхности конуса нарастания уже появляются листовые зачатки (листовые примордии) как боковые выпуклости. Будучи местом множественных антиклинальных митозов, они маркируют упомянутое ранее фракционирование меристемы. Клетки листовых зачатков раньше, чем клетки стебля вступают в фазу растяжения, так что молодые листья перерастают конус нарастания. При этом проявляется градиент возрастающего растяжения клеток (постэмбриональный рост!), который распространяется от конуса нарастания побега базипетально к зонам дифференциации. Наружная сторона почечных чешуй (их «абаксиальная», а позже нижняя сторона) обгоняет в растяжении лежащую ближе к конусу нарастания «адаксиальную» (верхнюю) сторону, так что молодые листья загибаются над конусом, окружают его сводом и таким образом вместе с ним образуют почку1.
1 Так растут не только почечные чешуи, но и все листья. — Примеч. ред.
Вообще, уже в листовом зачатке закладывается различие между верхней и нижней сторонами будущих листьев (их дорсовентральная структура). Это проявляется, например, в асимметричной экспрессии генов. Из адаксиального участка зачатка позже возникают верхняя эпидерма, палисадная паренхима и тяжи ксилемы листа, а из абаксиального — соответственно флоэма, губчатая паренхима и нижняя эпидерма (см. 4.3.1.1).
Образование и расположение листовых зачатков определяются с помощью индол- 3-уксусной кислоты — фитогормона, который в области меристемы транспортируется строго базипетально (см. 7.6.13).
В конусе нарастания, помимо подразделений меристемы, можно различить три следующие одна за другой зоны: зона инициалей (ее длина 10—50 мкм); затем морфогенетическая зона, или зона дифференциацки (20 — 80 мкм), в которой закладываются зачатки листьев и позже устанавливается листорасположение; и,
наконец, гистогенетическая зона, где происходит переход к постоянным клеткам и тканям. Она преимущественно соответствует зоне растяжения стебля.
Апикальную меристему побега можно также формально подразделить по геометрическим особенностям, что часто целесообразно в описательных целях. В соответствии с этим следует отличать объемную меристему (в которой деления клеток идут во всех направлениях) от пластинчатой меристемы (деления только в одной плоскости, клеточные стенки относительно этой плоскости антиклинальны) и тяжевой меристемы (одноразмерная, возникновение из клеточных рядов путем поперечных делений). В апикальной и первичной меристемах корпус соответствует объемной меристеме, туника — пластинчатой меристеме, а прокамбий — тяжевой меристеме.
3.1.1.2. Конус нарастания корня
Конус нарастания корня прикрыт корневым чехликом — калиптрой (в пер. с греч. — покров, чехол). Стенки самых внешних, самых старых клеток чехлика ослизняются благодаря обильному выделению пектина. Таким образом, клетки чехлика недолговечны, они сменяются через несколько дней и восполняются от корневой меристемы. Они претерпевают быструю терминальную дифференциацию, как это часто бывает у растений (например, при вторичном утолщении; см. 4.2.8.2, и в перидерме; см. 3.2.2.2). Корневой чехлик облегчает продвижение в почве нежного конуса нарастания корня, но в первую очередь является органом восприятия силы тяжести (см. 8.3.1.2). В этом важную роль играют заметные амилопласты с крупными крахмальными зернами. Они образуются преимущественно в продольных рядах клеток в центральной области чехлика, которую называют колумелла (лат. — колонка, столбик).
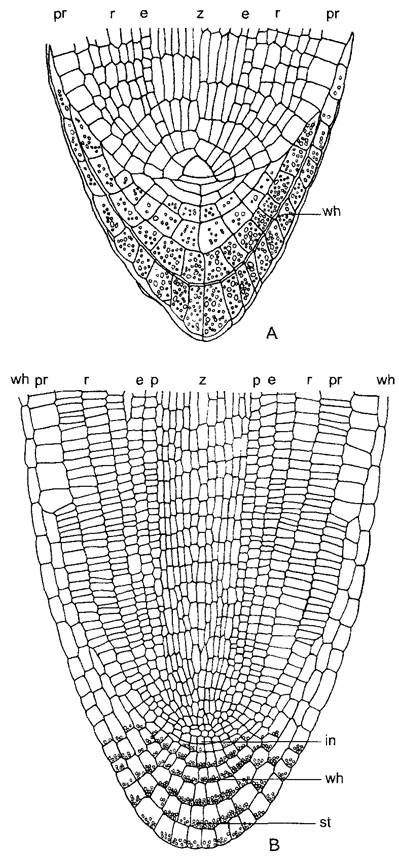
У большинства птеридофитов в центре конуса нарастания корня находится, как и в побеге, тетраэдрическая апикальная клетка (рис. 3.6, А). Будучи четырехгранной, она отделяет по всем четырем плоскостям клетки. Отложенные наружу клетки путем дальнейших делений образуют корневой чехлик. У голосеменных и покрытосеменных, напротив, апикальной клетки нет. На ее месте у голосеменных имеются две группы инициальных клеток. Из них внутренние образуют основную массу корня путем чередующихся антиклинальных и периклинальных делений, в то время как внешние — ткани первичной коры и нечетко отграниченный от нее чехлик. Наконец, у покрытосеменных на вершине апекса корня часто встречается комплексный образовательный центр, состоящий из нескольких независимых групп инициалей, который дает начало разным постоянным тканям (чехлику, эпидерме, первичной коре и центральному цилиндру) (рис. 3.6, В; см. также 4.4.2.1). При этом судьба производных инициалей в корне предопределена намного строже, чем в побеге, так что здесь можно говорить о гистогенах с упорядоченными клеточными линиями. Искусственное исключение определенных инициалей путем локального разрушения клеток с помощью лазерного излучения часто ведет к тому, что соответствующие слои тканей в корне вообще не могут образоваться (см. рис. 7.26).
В отдельных случаях, однако, структура инициальной зоны варьирует. Например, в апексе корня злаков самый внешний слой эвмеристемы (протодерма), который производит покровную ткань корня (ризодерму), вместе с лежащим под ним слоем меристемы, из которого образуется первичная кора, объединены в единую инициальную группу. Снаружи от нее лежит калиптроген — слой меристемы, производящий корневой чехлик. Однако у большинства настоящих двудольных корневой чехлик образуется путем антиклинальных1 делений из той же группы инициалей, которая образует и протодерму (дерматокалиптроген) (рис. 3.6, В). Под ней лежит второй этаж инициальных клеток, которые образуют первичную кору с ее внутренней барьерной тканью — эндодермой. Наконец, третий этаж инициальных клеток, плерома, производит центральный цилиндр с перициклом — частью центрального цилиндра.
1 Это параллельные поверхности периклинальные деления. — Примеч. ред.
Подобным «закрытым» конусам нарастания корня, в которых на всю жизнь сохраняются три четко различимых слоя инициалей как настоящих гистогенов для корневого чехлика, первичной коры и центрального цилиндра (например, Arabidopsis) (см. рис. 7.26), противостоят открытые конуса некоторых покрытосеменных (пример: лук). У них первоначальное разграничение гистогенов скоро исчезает вследствие беспорядочного разрастания комплекса инициалей, так что все постоянные ткани образуются из единой группы меристем этических клеток, в чем проявляется вторичное сходство с голосеменными.
Рис. 3.6. Апекс корня и корневой чехлик (А — по E.Strasburger; В — по L. Кnу): А—продольный срез кончика корня папоротника Pteris crética. Четырехгранная апикальная клетка выделена серым цветом (160х); В — продольный срез кончика корня Brassica napus, растения, относимого к настоящим двудольным. Самый наружный из трех слоев инициалей (дерматокалиптроген, in) производит дерматоген, из которого возникнет ризодерма, и корневой чехлик wh, клетки которого содержат легко различимые крахмальные зерна-статолиты (для восприятия силы тяжести; см. 8.3.1.2). Лежащий над ним второй слой инициалей производит клетки первичной коры с эндодермой. Наконец, третий слой инициалей производит центральный цилиндр с перициклом (50х) (ср. также рис. 7.26); е—эндодерма; in—зона инициалей; р—перицикл; рг— протодерма или соответственно ризодерма; г—первичная кора; st—крахмальные зерна-статолиты; wh—корневой чехлик; z—центральный цилиндр
3.1.2. Латеральные меристемы (камбии)
Инициали камбиев отличаются от соответствующих клеток апикальной мери
стемы более крупными размерами и сильной вакуолизацией. В прозенхимных, так называемых веретеновидных инициалах камбиев побега или корня (см. 4.2.8.2), от которых происходят клетки вторичных проводящих тканей, эта вакуолизация обусловливает особую форму деления клетки: ядро делится в плазматическом тяже, пронизывающем вдоль продольновытянутую вакуоль, фрагмопласт растет в нем центрифугально (см. рис. 2.32). Это занимает необычно много времени, так как веретеновидные инициали могут быть длиной больше миллиметра.
В большинстве случаев инициали камбия происходят не непосредственно от апикальной или первичной меристемы, а возникают путем реэмбрионизации клеток постоянных тканей.
Это свойственно пробковому камбию (см. 3.2,2.2) и большей частью также так называемому межпучковому камбию побегов (см. рис. 4.44).
Структуру и функции камбиев можно понять только на фоне морфологических и анатомических данных, поэтому они будут описаны ниже (см. 4.2.8.2; 4.4.2.3).
Бокс 3.1. Остаточные меристемы и меристемоиды
Позади гистогенетической зоны, т.е. далеко за конусом нарастания, остатки меристемы в форме ограниченных слоев клеток, их групп или тяжей сохраняют способность к делению еще некоторое время. Например, у многих однодольных базальные участки междоузлий долгое время остаются меристематическими как интеркалярные зоны роста. Тяжи камбия в проводящих пучках двудольных позже способствуют вторичному утолщению стеблей (см. 4.2.8.2). Перицикл корня соответствующим образом дает боковые корни (см. 4.4.2.2). У многих однодольных листья долгое время продолжают расти у своего основания, в то время как кончик листа уже полностью дифференцирован. Крайнее проявление такого роста выявлено у голосеменного растения Welwitschia из юго- западной части Африки (см. рис. 11.211, А), у которого два лентовидных листа растут в основании неограниченно, в то время как концевые зоны постоянно укорачиваются, отмирая.
В зонах дифференцировки побегов и листьев часто встречаются мелкие гнезда активно делящихся клеток, не содержащие инициалей. Поэтому все клетки таких меристемоидов1становятся в конце концов постоянными клетками, которые отличаются от остальных клеток ткани визуально и функционально (идиобласты). Из меристемоидов происходят, например, устьичный аппарат и многоклеточные волоски (см. рис. 3.13, 3.14). Листовые зачатки на конусе нарастания также в конечном счете представляют собой меристемоиды2, что выражается в ограниченном росте листьев (выше упомянутая Welwitschiа является исключением).
Меристемоиды часто возникают от единичных клеток, возникших при неравном лечении из одной материнской клетки образуется одна более крупная, сильно вакуолизированная, далее не делящаяся клетка, и одна более мелкая, богатая плазмой клетка, которая путем ограниченно продолжающихся делений образует меристемоид3. С точки эрения биологии развития интересно размещение меристемоидов или возникающих из них образований они составляют правильные узоры (см рис 3 13, бокс 4 1). Это результат плотного расположения полей торможения, которые однажды возникший меристемоид образует вокруг себя и внутри которых возникновение следующих меристемоидов подавлено. На этом основаны, например, закономерности листорасположения (см 4 2 2, 7 4 2)
1 Меристемоиды — не группы («гнезда») клеток, а сами эти клетки — Примеч. ред.
2 Это частное мнение авторов учебника, не разделяемое большинством ботаников — Примеч. ред.
3 Сама мелкая клетка уже является (первичным) меристемоидом — Примеч. ред.